
Distinct Profiles of Desensitization of µ-Opioid Receptors Caused by Remifentanil or Fentanyl: In Vitro Assay with Cells and Three-Dimensional Structural Analyses
 , , , , ,
, , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Dose-Response Curves of REM, FEN, and D-Ala(2)-N-Me-Phe(4)-Glyol(5)-Enkephalin (DAMGO) with Cells Stably Expressing MOR
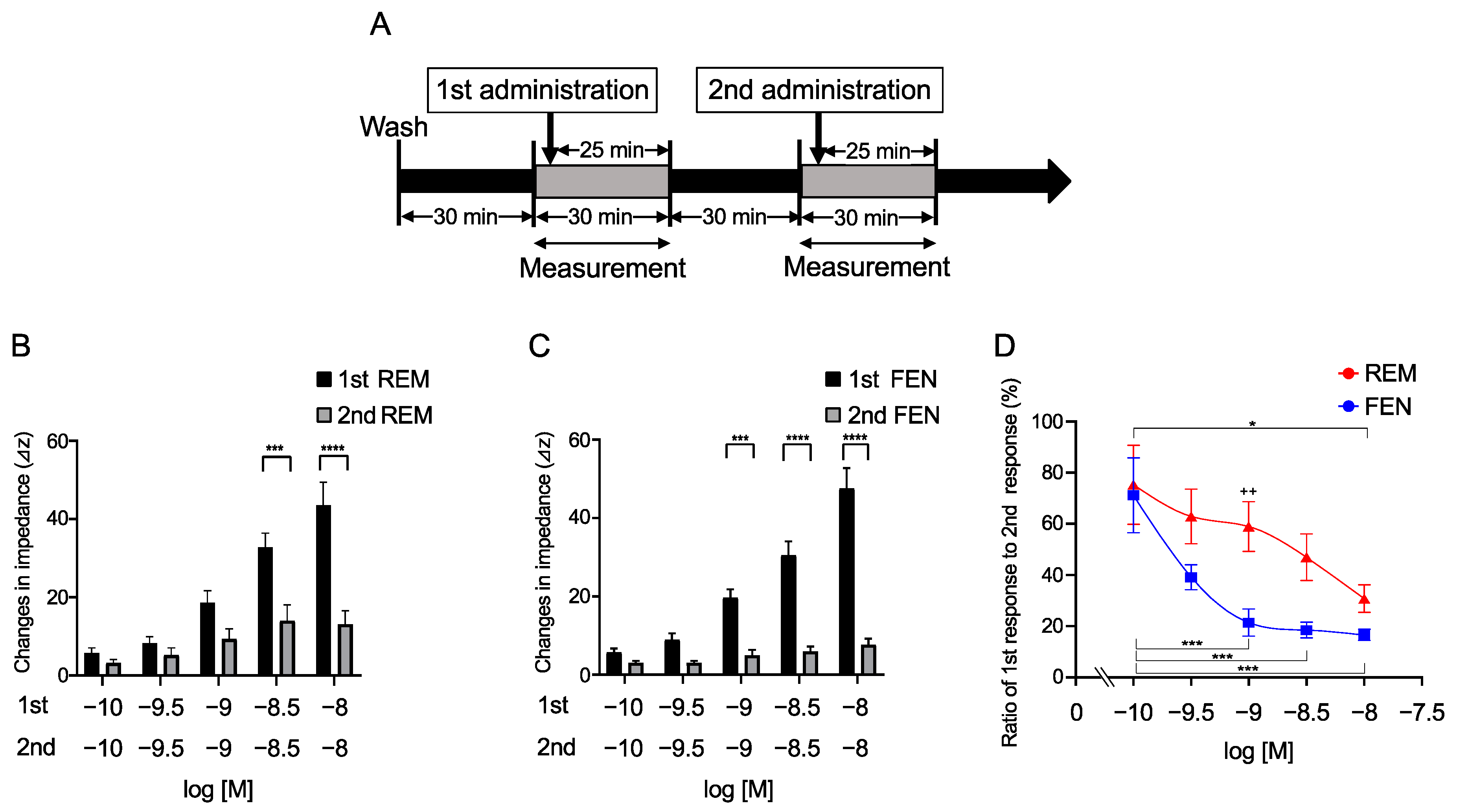
2.2. Repetitive Administration of REM and FEN in Cells for Determination of MOR Desensitization with the CellKeyTM
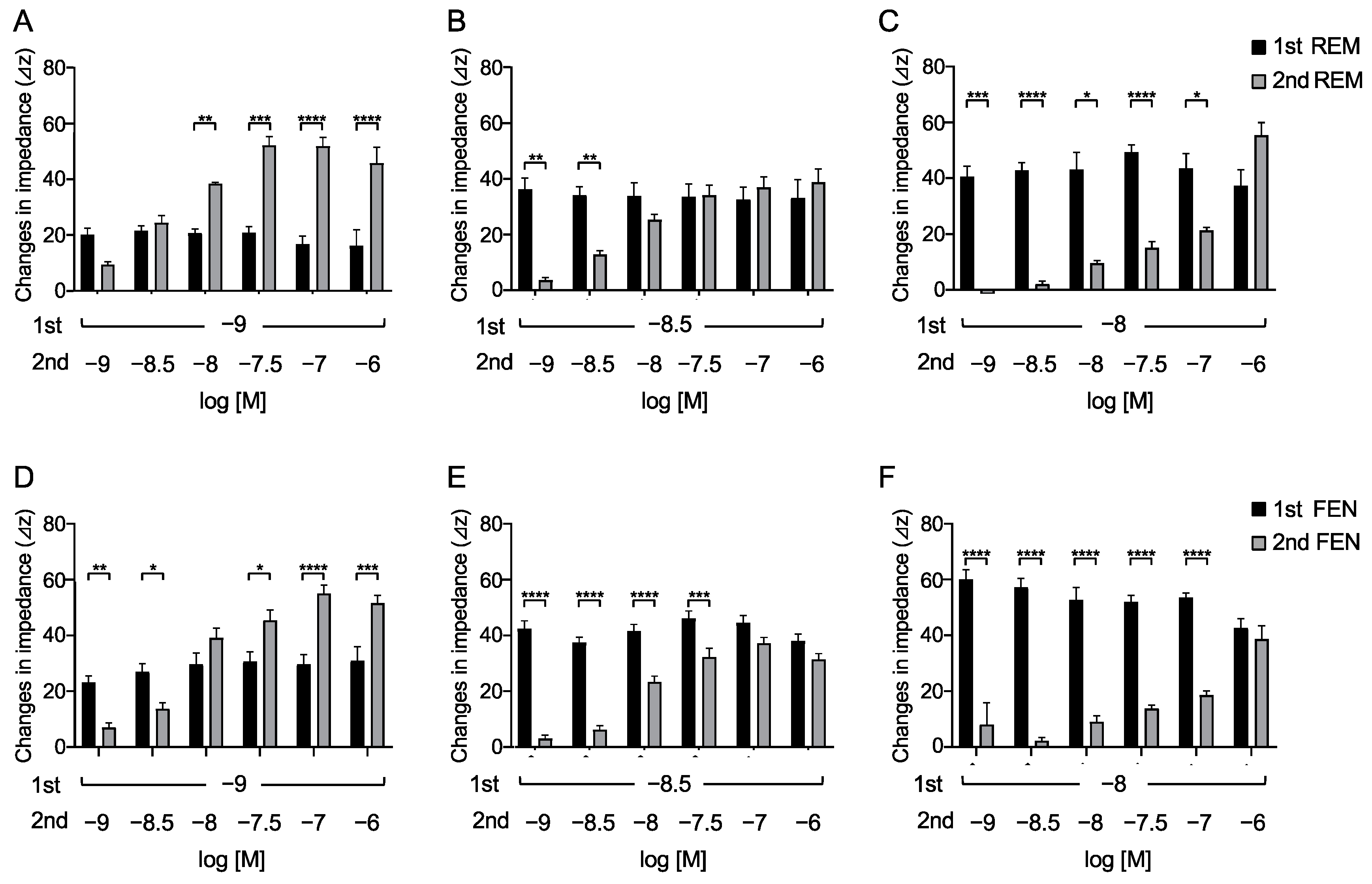
2.3. Requirements of Secondary-Administrated Analgesic Concentrations to Achieve Equivalent Responses Compared with the 1st Response
2.4. Opioid Switching Assay
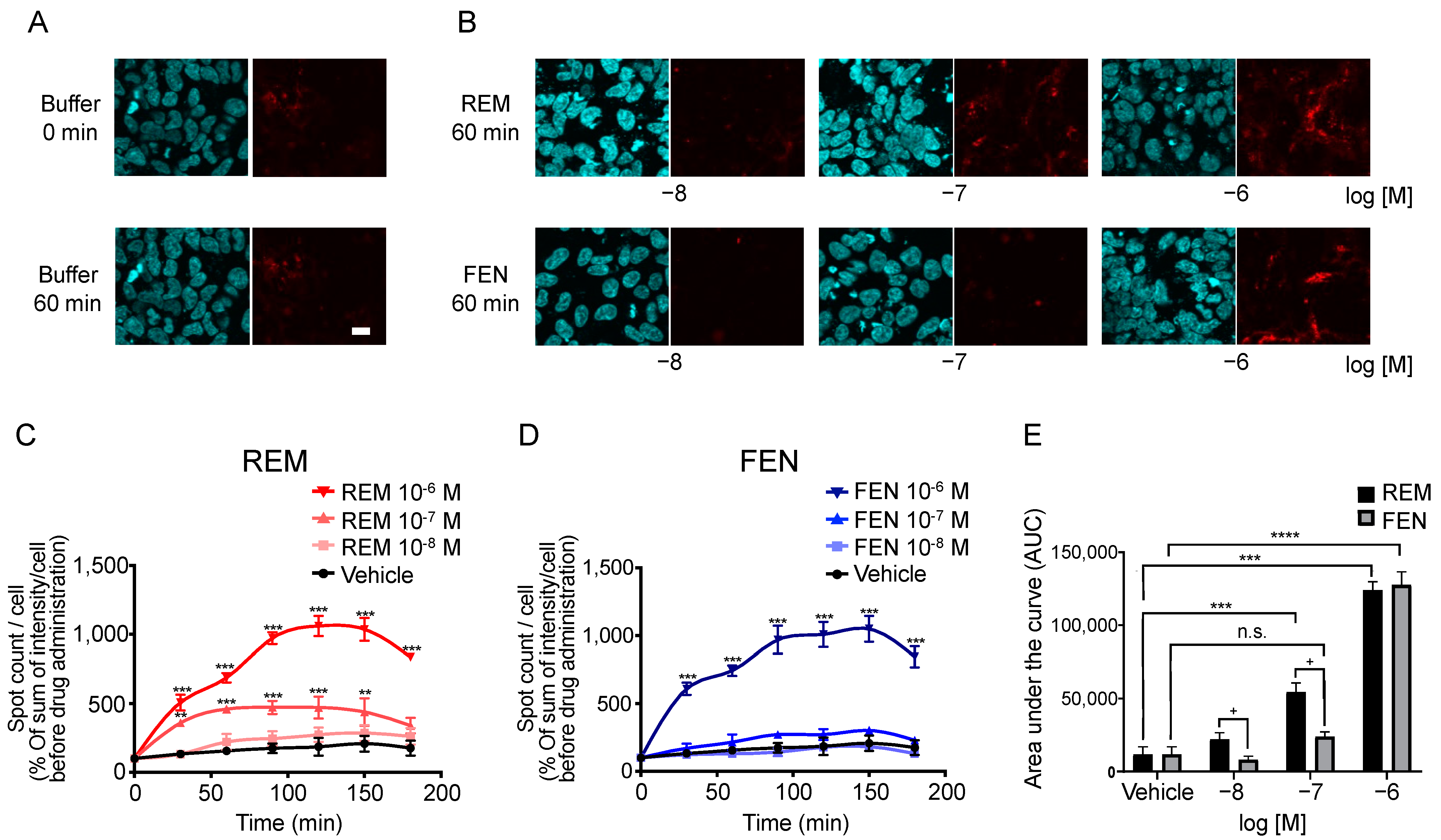
2.5. Visual Internalization Assay of ORs in Cells Stably Expressing Halotag®-Tagged ORs
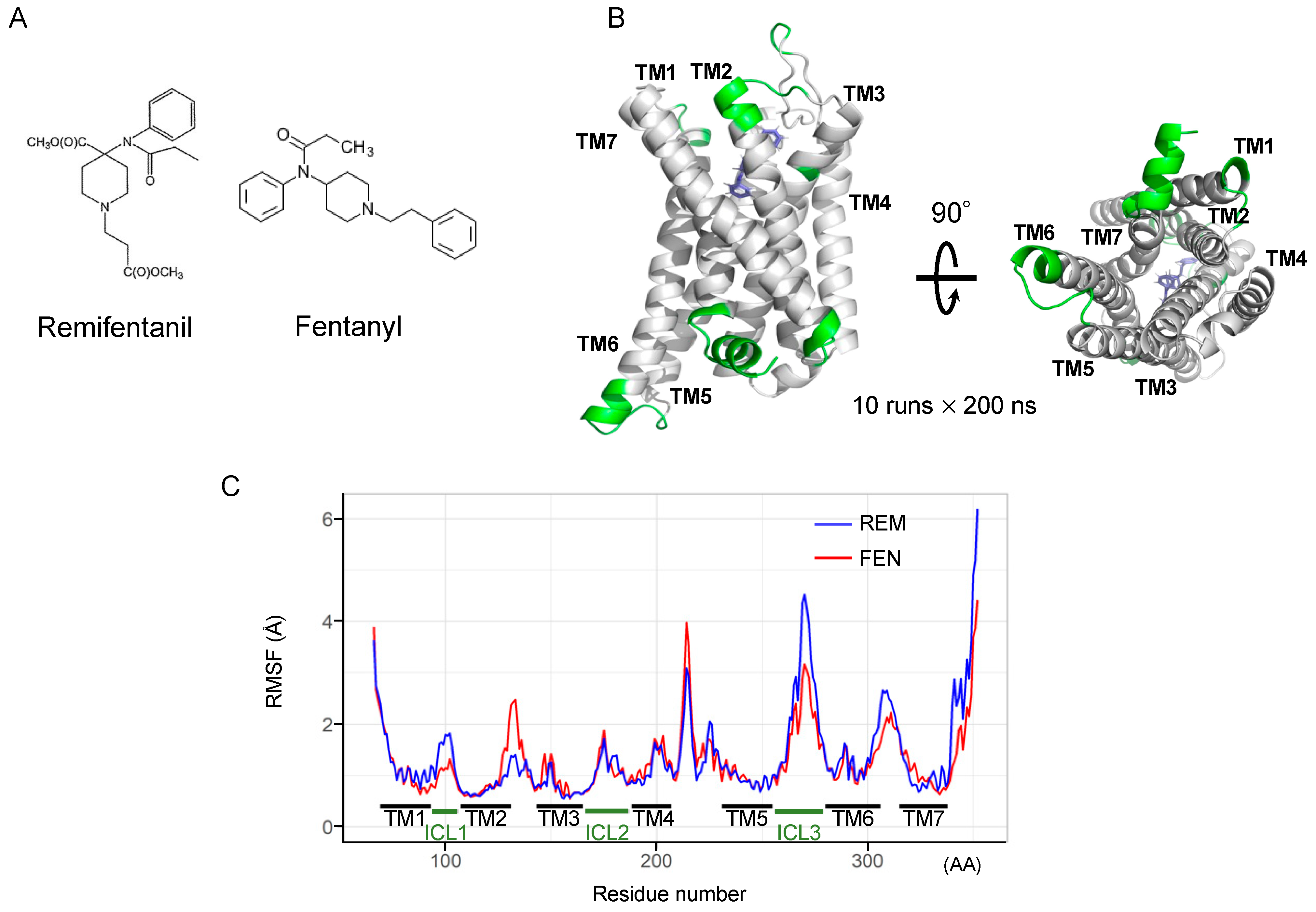
2.6. Binding Mode Estimation by Ligand Docking and Molecular Dynamics (MD) Simulation
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmids and Generation of Stable Cell Line
4.2. Cell Culture
4.3. Chemicals
4.4. Functional Analysis of ORs with the CellKeyTM System
4.5. Functional Analysis of ORs Using Repetitive Administration Protocol with CellKeyTM
4.6. Intracellular cAMP Assay with GloSensor®
4.7. β Arrestin Recruitment Assay with PathHunter®
4.8. Internalization Assay of MOR
4.9. Ligand Docking and Molecular Dynamics Simulation
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, L.J.; Perry, C.M. Remifentanil: A review of its use during the induction and maintenance of general anaesthesia. Drugs 2005, 65, 1793–1823. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.N.; Pavone, K.J.; Naranjo, M. Multimodal General Anesthesia: Theory and practice. Anesth. Analg. 2018, 127, 1246–1258. [Google Scholar] [CrossRef]
- Egan, T.D.; Lemmens, H.J.; Fiset, P.; Hermann, D.J.; Muir, K.T.; Stanski, D.R.; Shafer, S.L. The pharmacokinetics of the new short-acting opioid remifentanil (GI87084B) in healthy adult male volunteers. Anesthesiology 1993, 79, 881–892. [Google Scholar] [CrossRef]
- Kieffer, B.L. Recent advances in molecular recognition and signal transduction of active peptides: Receptors for opioid peptides. Cell. Mol. Neurobiol. 1995, 15, 615–635. [Google Scholar] [CrossRef] [PubMed]
- Law, P.Y.W.; Wong, Y.H.; Loh, H.H. Molecular mechanisms and regulation of opioid receptor signaling. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.T.; Ingram, S.L.; Henderson, G.; Chavkin, C.; von Zastrow, M.; Schulz, S.; Koch, T.; Evans, C.J.; Christie, M.J. Regulation of μ-opioid receptors: Desensitization, phosphorylation, internalization, and tolerance. Pharmacol. Rev. 2013, 65, 223–254. [Google Scholar] [CrossRef]
- Azzam, A.A.H.; McDonald, J.; Lambert, D.G. Hot topics in opioid pharmacology: Mixed and biased opioids. Br. J. Anaesth. 2019, 122, e136–e145. [Google Scholar] [CrossRef]
- Miyano, K.; Sudo, Y.; Yokoyama, A.; Hisaoka-Nakashima, K.; Morioka, N.; Takebayashi, M.; Nakata, Y.; Higami, Y.; Uezono, Y. History of the G protein-coupled receptor (GPCR) assays from traditional to a state-of-the-art biosensor assay. J. Pharmacol. Sci. 2014, 126, 302–309. [Google Scholar] [CrossRef]
- He, L.; Gooding, S.W.; Lewis, E.; Felth, L.C.; Gaur, A.; Whistler, J.L. Pharmacological and genetic manipulations at the µ-opioid receptor reveal arrestin-3 engagement limits analgesic tolerance and does not exacerbate respiratory depression in mice. Neuropsychopharmacology 2021, 46, 2241–2249. [Google Scholar] [CrossRef]
- Zamorano, C.A.; Bruchas, M.R. Visualizing ligand bias at the Mu-opioid receptor. Cell 2022, 185, 4251–4253. [Google Scholar] [CrossRef]
- Allouche, S.; Noble, F.; Marie, N. Opioid receptor desensitization: Mechanisms and its link to tolerance. Front. Pharmacol. 2014, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Shanthanna, H.; Ladha, K.S.; Kehlet, H.; Joshi, G.P. Perioperative opioid administration. Anesthesiology 2021, 134, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Colvin, L.A.; Bull, F.; Hales, T.G. Perioperative opioid analgesia—When is enough too much? A review of opioid-induced tolerance and hyperalgesia. Lancet 2019, 393, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Aurilio, C.; Pace, M.C.; Pota, V.; Sansone, P.; Barbarisi, M.; Grella, E.; Passavanti, M.B. Opioids switching with transdermal systems in chronic cancer pain. J. Exp. Clin. Cancer Res. 2009, 28, 61. [Google Scholar] [CrossRef]
- Fletcher, D.; Martinez, V. Opioid-induced hyperalgesia in patients after surgery: A systematic review and a meta-analysis. Br. J. Anaesth. 2014, 112, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, A.; Miyano, K.; Nakayama, D.; Mizobuchi, Y.; Uezono, E.; Ohshima, K.; Karasawa, Y.; Kuroda, Y.; Nonaka, M.; Yamaguchi, K.; et al. Novel opioid analgesics for the development of transdermal opioid patches that possess morphine-like pharmacological profiles rather than fentanyl: Possible opioid switching alternatives among patch formula. Anesth. Analg. 2022, 134, 1082–1093. [Google Scholar] [CrossRef]
- Kim, S.H.; Stoicea, N.; Soghomonyan, S.; Bergese, S.D. Intraoperative use of remifentanil and opioid induced hyperalgesia/acute opioid tolerance: Systematic review. Front. Pharmacol. 2014, 5, 108. [Google Scholar] [CrossRef]
- Nowoczyn, M.; Marie, N.; Coulbault, L.; Hervault, M.; Davis, A.; Hanouz, J.L.; Allouche, S. Remifentanil produces cross-desensitization and tolerance with morphine on the mu-opioid receptor. Neuropharmacology 2013, 73, 368–379. [Google Scholar] [CrossRef]
- Marie, N.; Aguila, B.; Allouche, S. Tracking the opioid receptors on the way of desensitization. Cell. Signal. 2006, 18, 1815–1833. [Google Scholar] [CrossRef]
- Zhang, J.; Barak, L.S.; Winkler, K.E.; Caron, M.G.; Ferguson, S.S. A central role for beta-arrestins and clathrin-coated vesicle-mediated endocytosis in beta2-adrenergic receptor resensitization. Differential regulation of receptor resensitization in two distinct cell types. J. Biol. Chem. 1997, 272, 27005–27014. [Google Scholar] [CrossRef]
- de Waal, P.W.; Shi, J.; You, E.; Wang, X.; Melcher, K.; Jiang, Y.; Xu, H.E.; Dickson, B.M. Molecular mechanisms of fentanyl mediated beta-arrestin biased signaling. PLoS Comput. Biol. 2020, 16, e1007394. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wang, Y.; He, B.; He, X.; Zhou, X.E.; Guo, S.; Rao, Q.; Yang, J.; Liu, J.; Zhou, Q.; et al. Molecular recognition of morphine and fentanyl by the human μ-opioid receptor. Cell 2022, 185, 4361–4375.e19. [Google Scholar] [CrossRef] [PubMed]
- Mizobuchi, Y.; Miyano, K.; Manabe, S.; Uezono, E.; Komatsu, A.; Kuroda, Y.; Nonaka, M.; Matsuoka, Y.; Sato, T.; Uezono, Y.; et al. Ketamine improves desensitization of µ-opioid receptors induced by repeated treatment with fentanyl but not with morphine. Biomolecules 2022, 12, 426. [Google Scholar] [CrossRef] [PubMed]
- Cahill, T.J., 3rd; Thomsen, A.R.; Tarrasch, J.T.; Plouffe, B.; Nguyen, A.H.; Yang, F.; Huang, L.Y.; Kahsai, A.W.; Bassoni, D.L.; Gavino, B.J.; et al. Distinct conformations of GPCR-beta-arrestin complexes mediate desensitization, signaling, and endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, 2562–2567. [Google Scholar] [CrossRef]
- Manabe, S.; Miyano, K.; Fujii, Y.; Ohshima, K.; Yoshida, Y.; Nonaka, M.; Uzu, M.; Matsuoka, Y.; Sato, T.; Uezono, Y.; et al. Possible biased analgesic of hydromorphone through the G protein-over beta-arrestin-mediated pathway: cAMP, CellKey™, and receptor internalization analyses. J. Pharmacol. Sci. 2019, 140, 171–177. [Google Scholar] [CrossRef]
- Lipiński, P.F.J.; Jarończyk, M.; Dobrowolski, J.C.; Sadlej, J. Molecular dynamics of fentanyl bound to μ-opioid receptor. J. Mol. Model. 2019, 25, 144. [Google Scholar] [CrossRef]
- Latorraca, N.R.; Wang, J.K.; Bauer, B.; Townshend, R.J.L.; Hollingsworth, S.A.; Olivieri, J.E.; Xu, H.E.; Sommer, M.E.; Dror, R.O. Molecular mechanism of GPCR-mediated arrestin activation. Nature 2018, 557, 452–456. [Google Scholar] [CrossRef]
- Ricarte, A.; Dalton, J.A.R.; Giraldo, J. Structural assessment of agonist efficacy in the mu-opioid receptor: Morphine and fentanyl elicit different activation patterns. J. Chem. Inf. Model. 2021, 61, 1251–1274. [Google Scholar] [CrossRef]
- Peters, M.F.; Knappenberger, K.S.; Wilkins, D.; Sygowski, L.A.; Lazor, L.A.; Liu, J.; Scott, C.W. Evaluation of cellular dielectric spectroscopy, a whole-cell, label-free technology for drug discovery on Gi-coupled GPCRs. J. Biomol. Screen. 2007, 12, 312–319. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uezono, E.; Mizobuchi, Y.; Miyano, K.; Ohbuchi, K.; Murata, H.; Komatsu, A.; Manabe, S.; Nonaka, M.; Hirokawa, T.; Yamaguchi, K.; et al. Distinct Profiles of Desensitization of µ-Opioid Receptors Caused by Remifentanil or Fentanyl: In Vitro Assay with Cells and Three-Dimensional Structural Analyses. Int. J. Mol. Sci. 2023, 24, 8369. https://doi.org/10.3390/ijms24098369
Uezono E, Mizobuchi Y, Miyano K, Ohbuchi K, Murata H, Komatsu A, Manabe S, Nonaka M, Hirokawa T, Yamaguchi K, et al. Distinct Profiles of Desensitization of µ-Opioid Receptors Caused by Remifentanil or Fentanyl: In Vitro Assay with Cells and Three-Dimensional Structural Analyses. International Journal of Molecular Sciences. 2023; 24(9):8369. https://doi.org/10.3390/ijms24098369
Chicago/Turabian StyleUezono, Eiko, Yusuke Mizobuchi, Kanako Miyano, Katsuya Ohbuchi, Hiroaki Murata, Akane Komatsu, Sei Manabe, Miki Nonaka, Takatsugu Hirokawa, Keisuke Yamaguchi, and et al. 2023. "Distinct Profiles of Desensitization of µ-Opioid Receptors Caused by Remifentanil or Fentanyl: In Vitro Assay with Cells and Three-Dimensional Structural Analyses" International Journal of Molecular Sciences 24, no. 9: 8369. https://doi.org/10.3390/ijms24098369
APA StyleUezono, E., Mizobuchi, Y., Miyano, K., Ohbuchi, K., Murata, H., Komatsu, A., Manabe, S., Nonaka, M., Hirokawa, T., Yamaguchi, K., Iseki, M., Uezono, Y., Hayashida, M., & Kawagoe, I. (2023). Distinct Profiles of Desensitization of µ-Opioid Receptors Caused by Remifentanil or Fentanyl: In Vitro Assay with Cells and Three-Dimensional Structural Analyses. International Journal of Molecular Sciences, 24(9), 8369. https://doi.org/10.3390/ijms24098369