

Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs


 ,
,  and
and 
Abstract
1. Introduction
2. Results
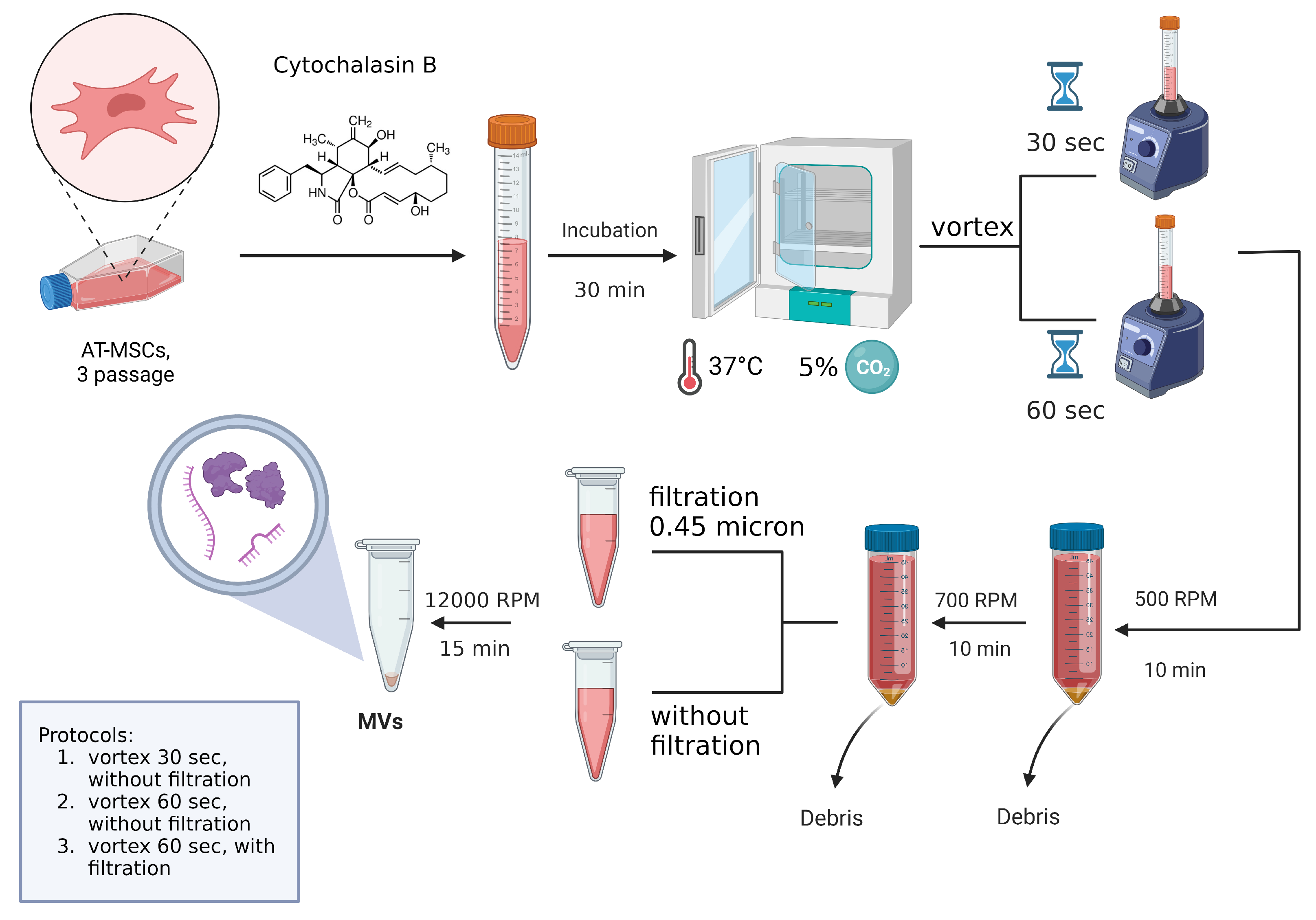
2.1. Analysis of AT-MSC-Derived EVs by Electron Microscopy
2.2. Cytokine Expression Profile of EVs
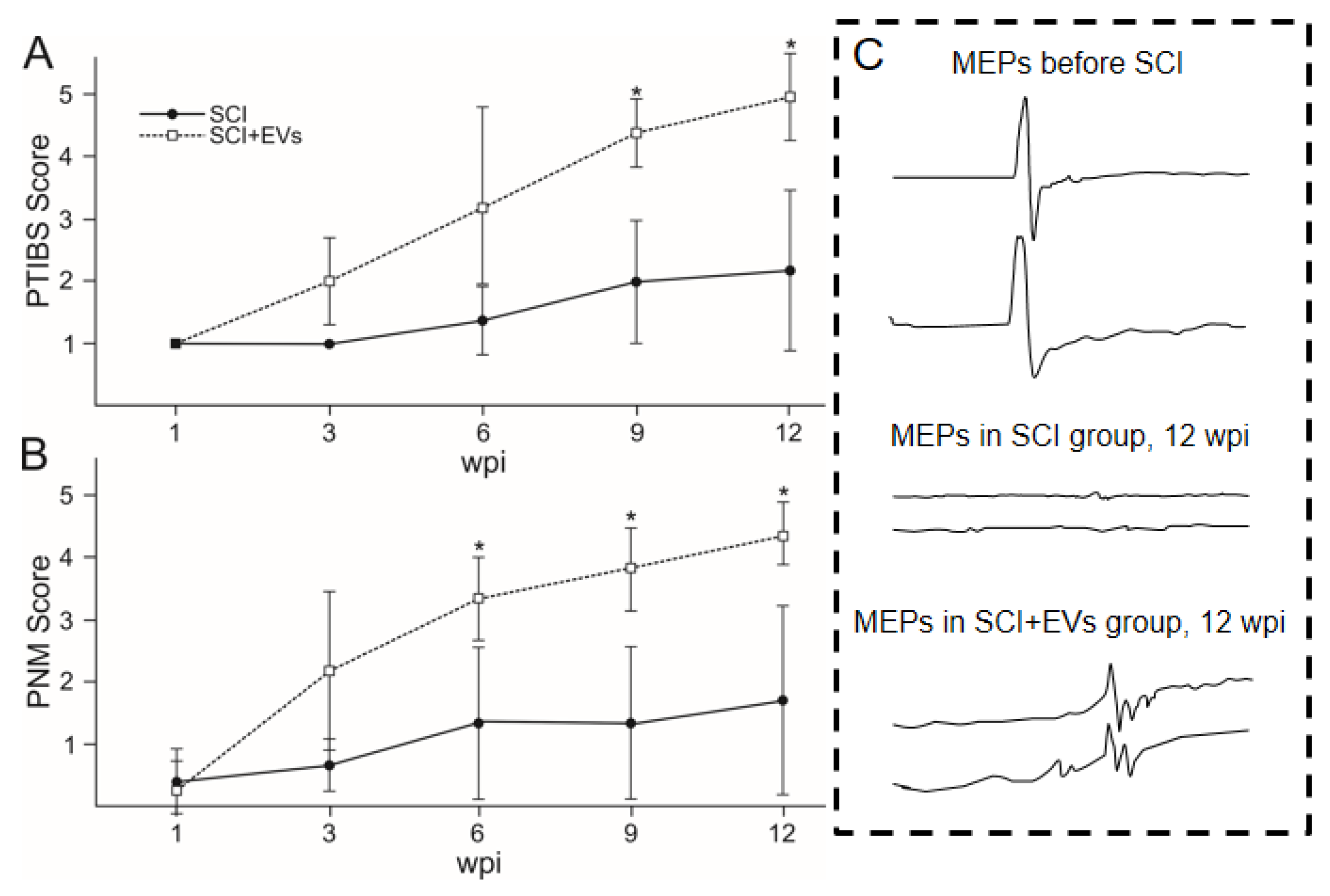
2.3. Behavioural Outcomes
2.4. Electrophysiology
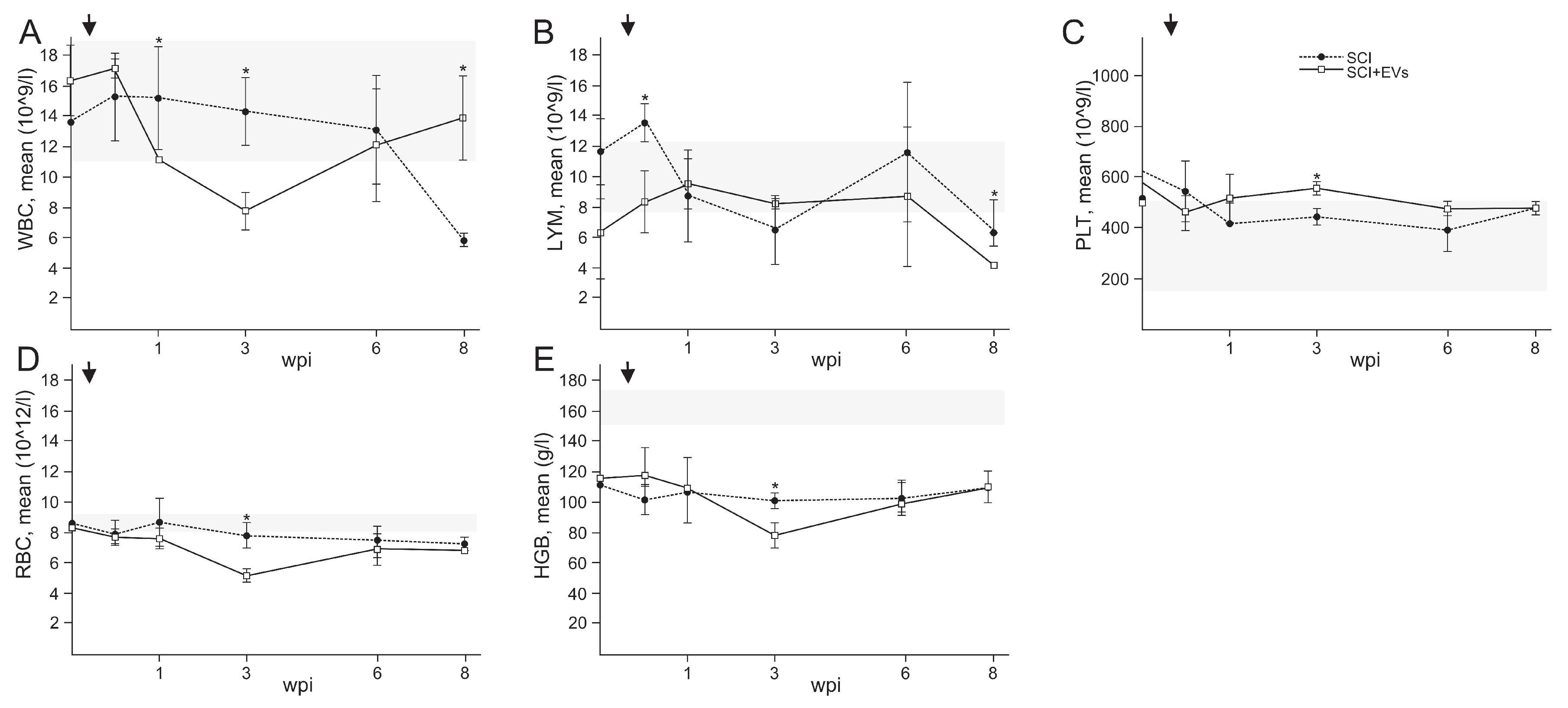
2.5. Routine Blood Examination EVs
2.6. Quantification of the Spared Tissue and Abnormal Cavities
2.7. Histological Evaluation of Myelin-Forming P0-Cells and Vascularization

3. Discussion
4. Materials and Methods
4.1. Cultivation of Mesenchymal Stem Cells and EVs Isolation
4.2. Electron Microscopic Analysis of EVs from AT-MSCs
4.3. Cytokine Assay
4.4. Animals
4.5. Spinal Cord Injury and Intrathecal Injection
4.6. Porcine Thoracic Injury Behavioral Scale
4.7. Porcine Neurological Motor Score
4.8. Electrophysiological Studies
4.9. Complete Blood Count Analysis
4.10. Histological Procedure
4.11. Immunofluorescence Analysis
4.12. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sezer, N.; Akkuş, S.; Uğurlu, F. Chronic Complications of Spinal Cord Injury. World J. Orthop. 2015, 6, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jones, Z.B.; Chen, X. Multiple Organ Dysfunction and Systemic Inflammation After Spinal Cord Injury: A Complex Relationship. J. Neuroinflamm. 2016, 13, 260. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Szabó, T.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, E.; Pap, E.; Kittel, A. Membrane Vesicles, Current State-of-the-art: Emerging Role of Extracellular Vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Di Bella, M. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.; Andreu, Z.; Zavec, A.; Borràs, F.; Buzas, E.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Zeng, Y.; Qiu, Y.; Jiang, W.; Shen, J.; Yao, X.; He, X.; Li, L.; Fu, B.; Liu, X. Biological Features of Extracellular Vesicles and Challenges. Front. Cell Dev. Biol. 2022, 10, 816698. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Galieva, L.; James, V.; Mukhamedshina, Y.; Rizvanov, A. Therapeutic Potential of Extracellular Vesicles for the Treatment of Nerve Disorders. Front. Neurosci. 2019, 13, 163. [Google Scholar] [CrossRef]
- Nieuwland, R.; Siljander, P.R.-M.; Falcón-Pérez, J.M.; Witwer, K.W. Reproducibility of Extracellular Vesicle Research. Eur. J. Cell Biol. 2022, 101, 151226. [Google Scholar] [CrossRef]
- Katsuda, T.; Tsuchiya, R.; Kosaka, N.; Yoshioka, Y.; Takagaki, K.; Oki, K.; Takeshita, F.; Sakai, Y.; Kuroda, M.; Ochiya, T. Human Adipose Tissue-derived Mesenchymal Stem Cells Secrete Functional Neprilysin-bound Exosomes. Sci. Rep. 2013, 3, 1197. [Google Scholar] [CrossRef]
- Han, Y.; Cao, X.; Guo, X.; Chu, T.; Li, H.; Xue, D.; Xin, L.; Yuan, H.; Ke, G.; Li, Z. Mesenchymal Stem Cell-derived Extracellular Vesicles Promote Microglial M2 Polarization After Subarachnoid Hemorrhage in Rats and Involve the Ampk/nf-κb Signaling Pathway. Biomed. Pharmacother. 2021, 133, 111048. [Google Scholar] [CrossRef]
- Yari, H.; Mikhailova, M.V.; Mardasi, M. Emerging Role of Mesenchymal Stromal Cells (mscs)-derived Exosome in Neurodegeneration-associated Conditions: A Groundbreaking Cell-free Approach. Stem Cell Res. Ther. 2022, 13, 423. [Google Scholar] [CrossRef]
- Schepici, G.; Silvestro, S.; Mazzon, E. Regenerative Effects of Exosomes-derived Mscs: An Overview on Spinal Cord Injury Experimental Studies. Biomedicines 2023, 11, 201. [Google Scholar] [CrossRef]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering Exosomes as Refined Biological Nanoplatforms for Drug Delivery. Acta Pharmacol. Sin. 2017, 38, 754–763. [Google Scholar] [CrossRef]
- Gangadaran, P.; Ahn, B. Extracellular Vesicle- and Extracellular Vesicle Mimetics-based Drug Delivery Systems: New Perspectives, Challenges, and Clinical Developments. Pharmaceutics 2020, 12, 442. [Google Scholar] [CrossRef]
- Nair, A.; Bu, J.; Rawding, P.A.; Do, S.C.; Li, H.; Hong, S. Cytochalasin B. Treatment and Osmotic Pressure Enhance the Production of Extracellular Vesicles (EVs) with Improved Drug Loading Capacity. Nanomaterials 2022, 12, 3. [Google Scholar] [CrossRef]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Dang, X.; Zeng, X. Targeted Therapeutic Delivery Using Engineered Exosomes and Its Applications in Cardiovascular Diseases. Gene 2016, 575, 377–384. [Google Scholar] [CrossRef]
- Jang, S.; Kim, O.; Yoon, C.; Choi, D.; Roh, T.; Park, J.; Nilsson, J.; Lötvall, J.; Kim, Y.; Gho, Y. Bioinspired Exosome-mimetic Nanovesicles for Targeted Delivery of Chemotherapeutics to Malignant Tumors. ACS Nano 2013, 7, 7698–7710. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.; Jeong, D.; Kim, J.; Cho, S.; Jang, S.; Han, C.; Kang, J.; Gho, Y.; Park, J. Microfluidic Fabrication of Cell-derived Nanovesicles as Endogenous RNA Carriers. Lab Chip 2014, 14, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.; Zou, S.; Ong, W.; Torta, F.; Alexandra, A.; Schiffelers, R.; Storm, G.; Wang, J.; Czarny, B.; Pastorin, G. Bioinspired Cell-derived Nanovesicles Versus Exosomes as Drug Delivery Systems: A Cost-effective Alternative. Sci. Rep. 2017, 7, 14322. [Google Scholar] [CrossRef] [PubMed]
- Toledano Furman, N.E.; Lupu-Haber, Y.; Bronshtein, T.; Kaneti, L.; Letko, N.; Weinstein, E.; Baruch, L.; Machluf, M. Reconstructed Stem Cell Nanoghosts: A Natural Tumor Targeting Platform. Nano Lett. 2013, 13, 3248–3255. [Google Scholar] [CrossRef] [PubMed]
- Gomzikova, M.; Kletukhina, S.; Kurbangaleeva, S.; Rizvanov, A. Evaluation of Cytochalasin B-induced Membrane Vesicles Fusion Specificity with Target Cells. BioMed Res. Int. 2018, 2018, 7053623. [Google Scholar] [CrossRef]
- Lee, J.H.T.; Jones, C.F.; Okon, E.B.; Anderson, L.; Tigchelaar, S.; Kooner, P.; Godbey, T.; Chua, B.; Gray, G.; Hildrandt, R.; et al. A novel porcine model of traumatic thoracic spinal cord injury. J. Neurotrauma 2013, 30, 142–159. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, S.; Ding, Y.; Li, S.; Yang, G.; Zhang, J.; Li, Y.; Cheng, J.; Lu, Y. Reference values of biochemical and hematological parameters for Guizhou minipigs. Exp. Biol. Med. 2011, 236, 477–482. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, C.; Wang, X.; Chen, H.; Wang, C.; Cao, P.; Gao, R.; Ren, X.; Luo, Z.; Wang, B. Timing of Surgical Decompression for Traumatic Cervical Spinal Cord Injury. Int. Orthop. 2015, 39, 2457–2463. [Google Scholar] [CrossRef]
- Yakushenko, E.V.; Lopatnikova, Y.A.; Sennikov, S.V. Interleukin-18 and its role in the immune response. Med. Immunol. 2005, 7, 355–364. [Google Scholar] [CrossRef]
- Li, X.; Chen, S.; Mao, L.; Li, D.; Xu, C.; Tian, H.; Mei, X. Zinc Improves Functional Recovery by Regulating the Secretion of Granulocyte Colony Stimulating Factor from Microglia/macrophages After Spinal Cord Injury. Front. Mol. Neurosci. 2019, 12, 18. [Google Scholar] [CrossRef]
- Bakhtiary, A.H.; Fatemi, M.J.; Emami Razavi, S.H.; Hassannejad, Z. Effects of Macrophage Colony-stimulating Factor on Functional Recovery of Locomotor Activity After Spinal Cord Injury in Rats. J. Neurosurg. Spine 2018, 41, 309–317. [Google Scholar]
- Schmitz, T.; Chew, L. Cytokines and Myelination in the Central Nervous System. Sci. World J. 2008, 8, 1119–1147. [Google Scholar] [CrossRef]
- Alves, S.; Churlaud, G.; Audrain, M.; Michaelsen-Preusse, K.; Fol, R.; Souchet, B.; Braudeau, J.; Korte, M.; Klatzmann, D.; Cartier, N. Interleukin-2 Improves Amyloid Pathology, Synaptic Failure and Memory in Alzheimer’s Disease Mice. Brain 2017, 140, 826–842. [Google Scholar] [CrossRef]
- Dansokho, C.; Aucouturier, P.; Dorothée, G. Beneficial Effect of Interleukin-2-based Immunomodulation in Alzheimer-like Pathology. Brain 2017, 140, e39. [Google Scholar] [CrossRef]
- Spittau, B. Interleukin 4-induced Neuroprotection and Regulation of Microglia Activation as a Therapeutic Approach in the MPTP Model of Parkinson’s Disease. Neural Regen. Res. 2017, 12, 1433–1434. [Google Scholar] [CrossRef]
- Hühner, L.; Rilka, J.; Gilsbach, R.; Zhou, X.; Machado, V.; Spittau, B. Interleukin-4 Protects Dopaminergic Neurons in Vitro but Is Dispensable for Mptp-induced Neurodegeneration in Vivo. Front. Mol. Neurosci. 2017, 10, 62. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, W.; Xu, F.; Dai, X.; Shi, L.; Cai, W. The Interleukin-4/ppary Signaling Axis Promotes Oligodendrocyte Differentiation and Remyelination After Brain Injury. PLoS Biol. 2019, 17, e3000330. [Google Scholar] [CrossRef]
- Nakayama, M.; Takahashi, K.; Kitamura, T. Induction of Differentiation and Apoptosis by Ligands of Peroxisome Proliferator-activated Receptor Gamma in Non-small Cell Lung Cancer. Cancer Res. 2000, 60, 7102–7109. [Google Scholar]
- Li, L.; Zhang, Y.; Mu, J.; Chen, J.; Zhang, C.; Cao, H.; Gao, J. Transplantation of Human Mesenchymal Stem-cell-derived Exosomes Immobilized in an Adhesive Hydrogel for Effective Treatment of Spinal Cord Injury. Nano Lett. 2020, 20, 4298–4305. [Google Scholar] [CrossRef]
- Romanelli, P.; Bieler, L.; Heimel, P.; Škokić, S.; Jakubecova, D.; Kreutzer, C.; Zaunmair, P.; Smolčić, T.; Benedetti, B.; Rohde, E. Enhancing Functional Recovery Through Intralesional Application of Extracellular Vesicles in a Rat Model of Traumatic Spinal Cord Injury. Nano Lett. 2022, 15, 795008. [Google Scholar] [CrossRef]
- Drommelschmidt, K.; Serdar, M.; Bendix, I.; Herz, J.; Bertling, F.; Prager, S. Mesenchymal Stem Cell-derived Extracellular Vesicles Ameliorate Inflammation-induced Preterm Brain Injury. Brain Behav. Immun. 2017, 60, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Thomi, G.; Joerger-Messerli, M.; Haesler, V.; Muri, L.; Surbek, D.; Schoeberlein, A. Intranasally Administered Exosomes from Umbilical Cord Stem Cells Have Preventive Neuroprotective Effects and Contribute to Functional Recovery After Perinatal Brain Injury. Cells 2019, 8, 855. [Google Scholar] [CrossRef] [PubMed]
- Figley, S.; Khosravi, R.; Legasto, J.; Tseng, Y.; Fehlings, M. Characterization of Vascular Disruption and Blood-spinal Cord Barrier Permeability Following Traumatic Spinal Cord Injury. J. Neurotrauma 2014, 31, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, C.; Yang, D.; Liao, Q.; Luo, H.; Wang, X.; Zhou, F.; Yang, X.; Yang, J.; Zeng, C. Mesenchymal Stem Cells-derived Extracellular Vesicles, via Mir-210, Improve Infarcted Cardiac Function by Promotion of Angiogenesis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2085–2092. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, M.; Gong, X.; Hu, J.; Chen, G.; Ning, X.; Huang, S. Human Umbilical Cord Mesenchymal Stem Cell Exosomes Enhance Angiogenesis Through the Wnt4/β-catenin Pathway. Stem Cells Transl. Med. 2020, 11, 61. [Google Scholar] [CrossRef]
- Wang, X.; Thomsen, P. Mesenchymal Stem Cell-derived Small Extracellular Vesicles and Bone Regeneration. Basic Clin. Pharmacol. Toxicol. 2021, 128, 18–36. [Google Scholar] [CrossRef]
- Kugler, E.; Greenwood, J.; MacDonald, R. The “neuro-glial-vascular” Unit: The Role of Glia in Neurovascular Unit Formation and Dysfunction. Front. Cell Dev. Biol. 2021, 9, 732820. [Google Scholar] [CrossRef]
- Oudega, M. Molecular and Cellular Mechanisms Underlying the Role of Blood Vessels in Spinal Cord Injury and Repair. Cell Tissue Res. 2012, 349, 269–288. [Google Scholar] [CrossRef]
- Kim, K.; Streijger, F.; Manouchehri, N.; So, K.; Shortt, K.; Okon, E.; Tigchelaar, S.; Cripton, P.; Kwon, B. Review of the UBC Porcine Model of Traumatic Spinal Cord Injury. J. Korean Neurosurg. Soc. 2018, 61, 539–547. [Google Scholar] [CrossRef]
- Shulman, I.; Ogurcov, S.; Kostennikov, A.; Rogozin, A.; Garanina, E.; Masgutova, G.; Sergeev, M.; Rizvanov, A.; Mukhamedshina, Y. Application of Autologous Peripheral Blood Mononuclear Cells into the Area of Spinal Cord Injury in a Subacute Period: A Feasibility Study in Pigs. Biology 2021, 10, 87. [Google Scholar] [CrossRef]
- Navarro, R.; Juhas, S.; Keshavarzi, S.; Juhasova, J.; Motlik, J.; Johe, K.; Marsala, S.; Scadeng, M.; Lazar, P.; Tomori, Z. Chronic Spinal Compression Model in Minipigs: A Systematic Behavioral, Qualitative, and Quantitative Neuropathological Study. J. Neurotrauma 2012, 29, 499–513. [Google Scholar] [CrossRef]
- Kostennikov, A.; Kabdesh, I.; Sabirov, D.; Timofeeva, A.; Rogozhin, A.; Shulman, I.; Rizvanov, A.; Mukhamedshina, Y. A Comparative Study of Mesenchymal Stem Cell-derived Extracellular Vesicles’ Local and Systemic Dose-dependent Administration in Rat Spinal Cord Injury. Biology 2022, 11, 1853. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants of Protocol | Nuclei of MSCs | Disorganized Non-Membrane-Bound Inclusion | Mitochondria | Multilamellar Bodies in the Cytoplasm | Shape | Size, μm 1 |
|---|---|---|---|---|---|---|
| 1st protocol (active vortexing of cells for 30 s) | Yes | Yes | Yes | multiple | N/A | N/A |
| 2nd protocol (active vortexing of cells for 60 s without post-filtering) | Yes | Yes | Yes | multiple | varies | 0.06–0.6 |
| 3rd protocol active vortexing of cells for 60 s with post-filtering) | No | No | Yes | individual | mostly round | 0.02–0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shulman, I.; Ageeva, T.; Kostennikov, A.; Ogurcov, S.; Tazetdinova, L.; Kabdesh, I.; Rogozhin, A.; Ganiev, I.; Rizvanov, A.; Mukhamedshina, Y. Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs. Int. J. Mol. Sci. 2023, 24, 8240. https://doi.org/10.3390/ijms24098240
Shulman I, Ageeva T, Kostennikov A, Ogurcov S, Tazetdinova L, Kabdesh I, Rogozhin A, Ganiev I, Rizvanov A, Mukhamedshina Y. Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs. International Journal of Molecular Sciences. 2023; 24(9):8240. https://doi.org/10.3390/ijms24098240
Chicago/Turabian StyleShulman, Ilya, Tatyana Ageeva, Alexander Kostennikov, Sergei Ogurcov, Leysan Tazetdinova, Ilyas Kabdesh, Alexander Rogozhin, Ilnur Ganiev, Albert Rizvanov, and Yana Mukhamedshina. 2023. "Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs" International Journal of Molecular Sciences 24, no. 9: 8240. https://doi.org/10.3390/ijms24098240
APA StyleShulman, I., Ageeva, T., Kostennikov, A., Ogurcov, S., Tazetdinova, L., Kabdesh, I., Rogozhin, A., Ganiev, I., Rizvanov, A., & Mukhamedshina, Y. (2023). Intrathecal Injection of Autologous Mesenchymal Stem-Cell-Derived Extracellular Vesicles in Spinal Cord Injury: A Feasibility Study in Pigs. International Journal of Molecular Sciences, 24(9), 8240. https://doi.org/10.3390/ijms24098240