
Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats
 , , , and
, , , and
Abstract

1. Introduction
2. Results
2.1. Kisspeptin-10 Suppresses Placental Activation of the Inflammasome-NLRP3 Pathway Caused by Maternal Hypothyroidism
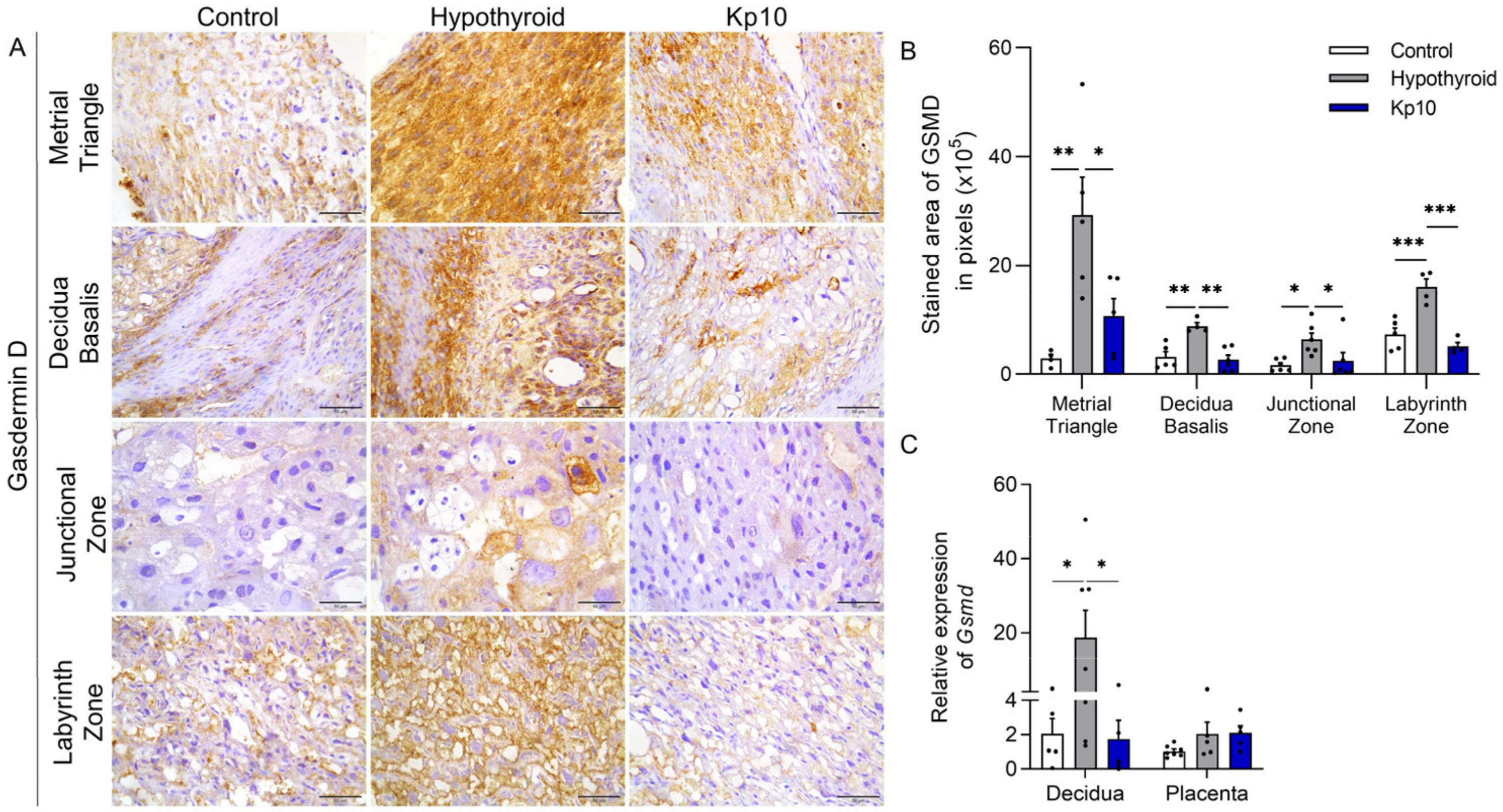
2.2. Kisspeptin-10 Blocks Decidual and Placental Pyroptosis Activation Caused by Maternal Hypothyroidism
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Necropsy and Collection of Material
4.3. Immunohistochemistry
4.4. qRT-PCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sullivan, S.A. Hypothyroidism in Pregnancy. Clin. Obstet. Gynecol. 2019, 62, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Biondi, B.; Kahaly, G.J.; Robertson, R.P. Thyroid Dysfunction and Diabetes Mellitus: Two Closely Associated Disorders. Endocr. Rev. 2019, 40, 789–824. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.Y.C.-P.P.; Chen, C.-Y.Y.C.-P.P.; Lin, K.-H.H. Biological Functions of Thyroid Hormone in Placenta. Int. J. Mol. Sci. 2015, 16, 4161–4179. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Xu, W.; Zhang, J. Association between Thyroid Dysfunction and Perinatal Outcomes in Women with Gestational Hypertension: A Retrospective Study. BMC Pregnancy Childbirth 2020, 20, 119. [Google Scholar] [CrossRef]
- Kent, N.L.; Atluri, S.C.; Cuffe, J.S.M. Maternal Hypothyroidism in Rats Reduces Placental Lactogen, Lowers Insulin Levels, and Causes Glucose Intolerance. Endocrinology 2022, 163, bqab231. [Google Scholar] [CrossRef]
- Shrestha, A.; Tripathi, P.; Dongol, A. Pregnancy Outcomes in Patients with Hypothyroidism. Kathmandu Univ. Med. J. (KUMJ) 2019, 17, 57–60. [Google Scholar]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Thyroid Hormones and Female Reproduction. Biol. Reprod. 2018, 99, 907–921. [Google Scholar] [CrossRef]
- Wang, J.; Gong, X.-H.; Peng, T.; Wu, J.-N. Association of Thyroid Function during Pregnancy with the Risk of Preeclampsia and Gestational Diabetes Mellitus. Endocr. Pract. 2021, 27, 819–825. [Google Scholar] [CrossRef]
- Silva, J.F.; Vidigal, P.N.; Galvão, D.D.; Boeloni, J.N.; Nunes, P.P.; Ocarino, N.M.; Nascimento, E.F.; Serakides, R. Fetal Growth Restriction in Hypothyroidism Is Associated with Changes in Proliferative Activity, Apoptosis and Vascularisation of the Placenta. Reprod. Fertil. Dev. 2012, 24, 923. [Google Scholar] [CrossRef]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Spatiotemporal Expression Profile of Proteases and Immunological, Angiogenic, Hormonal and Apoptotic Mediators in Rat Placenta before and during Intrauterine Trophoblast Migration. Reprod. Fertil. Dev. 2017, 29, 1774–1786. [Google Scholar] [CrossRef]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Placental Angiogenic and Hormonal Factors Are Affected by Thyroid Hormones in Rats. Pathol. Res. Pract. 2015, 211, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Maternal Thyroid Dysfunction Affects Placental Profile of Inflammatory Mediators and the Intrauterine Trophoblast Migration Kinetics. Reproduction 2014, 147, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.A.; Silva, J.F.; Ocarino, N.M.; Silva, C.L.R.; Gomes, L.A.; Assunção, G.S.M.; Silva, C.M.O.; Oliveira, K.P.; Serakides, R. Efeito Do Hipotireoidismo Materno Na Expressão Espaço-Temporal de Mediadores Imunológicos e População de Células Natural Killers Na Decídua e Na Glândula Metrial de Ratas. Arq. Bras. Med. Veterinária E Zootec. 2020, 72, 177–190. [Google Scholar] [CrossRef]
- Souza, C.A.; Silva, J.F.; Silva, C.L.R.; Ocarino, N.M.; Serakides, R. Thyroid Hormones Affect Decidualization and Angiogenesis in the Decidua and Metrial Gland of Rats. Pesqui. Vet. Bras. 2017, 37, 1002–1014. [Google Scholar] [CrossRef]
- Dos Anjos Cordeiro, J.M.; Santos, L.C.; de Oliveira, L.S.; Santos, B.R.; Santos, E.O.; Barbosa, E.M.; de Macêdo, I.O.; de Freitas, G.J.C.; de Assis Santos, D.; de Lavor, M.S.L.; et al. Maternal Hypothyroidism Causes Oxidative Stress and Endoplasmic Reticulum Stress in the Maternal-Fetal Interface of Rats. Free Radic. Biol. Med. 2022, 191, 24–39. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Spasic-Boskovic, O.; Jauniaux, E.; Charnock-Jones, D.S.; Burton, G.J. Nuclear Factor-ΚB, P38, and Stress-Activated Protein Kinase Mitogen-Activated Protein Kinase Signaling Pathways Regulate Proinflammatory Cytokines and Apoptosis in Human Placental Explants in Response to Oxidative Stress: Effects of Antioxidant Vitamins. Am. J. Pathol. 2007, 170, 1511–1520. [Google Scholar] [CrossRef]
- Zhang, K. Integration of ER Stress, Oxidative Stress and the Inflammatory Response in Health and Disease. Int. J. Clin. Exp. Med. 2010, 3, 33–40. [Google Scholar]
- Blevins, H.M.; Xu, Y.; Biby, S.; Zhang, S. The NLRP3 Inflammasome Pathway: A Review of Mechanisms and Inhibitors for the Treatment of Inflammatory Diseases. Front. Aging Neurosci. 2022, 14, 879021. [Google Scholar] [CrossRef]
- Jo, E.K.; Kim, J.K.; Shin, D.M.; Sasakawa, C. Molecular Mechanisms Regulating NLRP3 Inflammasome Activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef]
- Li, W.; Cao, T.; Luo, C.; Cai, J.; Zhou, X.; Xiao, X.; Liu, S. Crosstalk between ER Stress, NLRP3 Inflammasome, and Inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 6129–6140. [Google Scholar] [CrossRef]
- Mangan, M.S.J.J.; Olhava, E.J.; Roush, W.R.; Seidel, H.M.; Glick, G.D.; Latz, E. Targeting the NLRP3 Inflammasome in Inflammatory Diseases. Nat. Rev. Drug Discov. 2018, 17, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 Inflammasome: Molecular Activation and Regulation to Therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Han, T.-L.; Zhou, X.; Qi, H.; Baker, P.N.; Zhou, W.; Zhang, H. Endoplasmic Reticulum Stress May Activate NLRP3 Inflammasomes via TXNIP in Preeclampsia. Cell Tissue Res. 2020, 379, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Peng, Q. NLRP3 Inhibitor Tranilast Attenuates Gestational Diabetes Mellitus in a Genetic Mouse Model. Drugs R D 2022, 22, 105–112. [Google Scholar] [CrossRef]
- Corrêa-Silva, S.; Alencar, A.P.; Moreli, J.B.; Borbely, A.U.; Lima, L.D.S.; Scavone, C.; Damasceno, D.C.; Rudge, M.V.C.C.; Bevilacqua, E.; Calderon, I.M.P.P.; et al. Hyperglycemia Induces Inflammatory Mediators in the Human Chorionic Villous. Cytokine 2018, 111, 41–48. [Google Scholar] [CrossRef]
- Gao, P.; Zha, Y.; Gong, X.; Qiao, F.; Liu, H. The Role of Maternal–Foetal Interface Inflammation Mediated by NLRP3 Inflammasome in the Pathogenesis of Recurrent Spontaneous Abortion. Placenta 2020, 101, 221–229. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Motomura, K.; Miller, D.; Garcia-Flores, V.; Galaz, J.; Romero, R. Inflammasomes: Their Role in Normal and Complicated Pregnancies. J. Immunol. 2019, 203, 2757–2769. [Google Scholar] [CrossRef]
- Shirasuna, K.; Karasawa, T.; Takahashi, M. Role of the NLRP3 Inflammasome in Preeclampsia. Front. Endocrinol. (Lausanne) 2020, 11, 80. [Google Scholar] [CrossRef]
- Zhou, F.; Li, C.; Zhang, S.Y. NLRP3 Inflammasome: A New Therapeutic Target for High-Risk Reproductive Disorders? Chin. Med. J. 2020, 134, 20–27. [Google Scholar] [CrossRef]
- Bin Cheng, S.; Nakashima, A.; Huber, W.J.; Davis, S.; Banerjee, S.; Huang, Z.; Saito, S.; Sadovsky, Y.; Sharma, S. Pyroptosis Is a Critical Inflammatory Pathway in the Placenta from Early Onset Preeclampsia and in Human Trophoblasts Exposed to Hypoxia and Endoplasmic Reticulum Stressors. Cell Death Dis. 2019, 10, 927. [Google Scholar] [CrossRef]
- Yamada, S.M.; Pontillo, A. The Genetics behind Inflammasome Regulation. Mol. Immunol. 2022, 145, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.R.; dos Anjos Cordeiro, J.M.; Santos, L.C.; de Oliveira, L.S.; Mendonça, L.D.; Santos, E.O.; de Macedo, I.O.; Szawka, R.E.; Serakides, R.; Silva, J.F. Maternal Hypothyroidism Reduces the Expression of the Kisspeptin/Kiss1r System in the Maternal-Fetal Interface of Rats. Reprod. Biol. 2022, 22, 100615. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.R.; dos Anjos Cordeiro, J.M.; Santos, L.C.; Barbosa, E.M.; Mendonça, L.D.; Santos, E.O.; de Macedo, I.O.; de Lavor, M.S.L.; Szawka, R.E.; Serakides, R.; et al. Kisspeptin Treatment Improves Fetal-Placental Development and Blocks Placental Oxidative Damage Caused by Maternal Hypothyroidism in an Experimental Rat Model. Front. Endocrinol. 2022, 13, 908240. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, O.; Shirshev, S. The Effect of Kisspeptin on the Functional Activity of Peripheral Blood Monocytes and Neutrophils in the Context of Physiological Pregnancy. J. Reprod. Immunol. 2022, 151, 103621. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, O.L.; Shirshev, S.V. The Role of Kisspeptin in Immune Tolerance Formation during Pregnancy. Dokl. Biol. Sci. 2014, 457, 258–260. [Google Scholar] [CrossRef]
- Xing, R.; Liu, F.; Yang, Y.; Cui, X.; Wang, T.; Xie, L.; Zhao, Y.; Fang, L.; Yi, T.; Zheng, B.; et al. GPR54 Deficiency Reduces the Treg Population and Aggravates Experimental Autoimmune Encephalomyelitis in Mice. Sci. China Life Sci. 2018, 61, 675–687. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in Cell Death, Inflammation, and Disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Li, Z.; Ji, S.; Jiang, M.-L.; Xu, Y.; Zhang, C.-J. The Regulation and Modification of GSDMD Signaling in Diseases. Front. Immunol. 2022, 13, 893912. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, X.; Shan, H.Y.; Gao, H.; Wang, P. MicroRNA-520c-3p Suppresses NLRP3 Inflammasome Activation and Inflammatory Cascade in Preeclampsia by Downregulating NLRP3. Inflamm. Res. 2019, 68, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Xia, L.Z.; Liang, L.; Chen, Y.; Wei, D.; Meng, J.; Wu, S.Y.; Wang, Z. MiR-124–3p Promotes Trophoblast Cell HTR-8/SVneo Pyroptosis by Targeting Placental Growth Factor. Placenta 2020, 101, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zou, H.; Liu, J.; Wang, J.; Ma, C.; Yin, J.; Peng, X.; Li, D.; Yang, Y.; Ren, Y.; et al. Inhibition of HMGB1 Ameliorates the Maternal-Fetal Interface Destruction in Unexplained Recurrent Spontaneous Abortion by Suppressing Pyroptosis Activation. Front. Immunol. 2021, 12, 782792. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Bektur, E.; Baycu, C.; Burukoglu Dönmez, D.; Kaygısız, B. Hypothyroidism increases expression of sterile inflammation proteins in rat heart tissue. Acta Endocrinol. (Bucharest Rom. 2005) 2019, 5, 39–45. [Google Scholar] [CrossRef] [PubMed]
- De Vasconcelos, N.M.; Lamkanfi, M. Recent Insights on Inflammasomes, Gasdermin Pores, and Pyroptosis. Cold Spring Harb. Perspect. Biol. 2020, 12, a036392. [Google Scholar] [CrossRef]
- Fang, Y.; Tian, S.; Pan, Y.; Li, W.; Wang, Q.; Tang, Y.; Yu, T.; Wu, X.; Shi, Y.; Ma, P.; et al. Pyroptosis: A New Frontier in Cancer. Biomed. Pharmacother. 2020, 121, 109595. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and Functions of Inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef]
- Sullivan-Pyke, C.; Haisenleder, D.J.; Senapati, S.; Nicolais, O.; Eisenberg, E.; Sammel, M.D.; Barnhart, K.T. Kisspeptin as a New Serum Biomarker to Discriminate Miscarriage from Viable Intrauterine Pregnancy. Fertil. Steril. 2018, 109, 137–141.e2. [Google Scholar] [CrossRef]
- Pérez-López, F.R.; López-Baena, M.T.; Varikasuvu, S.R.; Ruiz-Román, R.; Fuentes-Carrasco, M.; Savirón-Cornudella, R. Preeclampsia and Gestational Hypertension Are Associated to Low Maternal Circulating Kisspeptin Levels: A Systematic Review and Meta-Analysis. Gynecol. Endocrinol. 2021, 37, 1055–1062. [Google Scholar] [CrossRef]
- Cartwright, J.E.; Williams, P.J. Altered Placental Expression of Kisspeptin and Its Receptor in Pre-Eclampsia. J. Endocrinol. 2012, 214, 79–85. [Google Scholar] [CrossRef]
- Ćetković, A.; Miljic, D.; Ljubić, A.; Patterson, M.; Ghatei, M.; Stamenkoví, J.; Nikolic-Djurovic, M.; Pekic, S.; Doknic, M.; Glišić, A.; et al. Plasma Kisspeptin Levels in Pregnancies with Diabetes and Hypertensive Disease as a Potential Marker of Placental Dysfunction and Adverse Perinatal Outcome. Endocr. Res. 2012, 37, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, R.V.; Drobintseva, A.O.; Alekseenkova, E.N.; Onopriychuk, A.R.; Arzhanova, O.N.; Polyakova, V.O.; Kvetnoy, I.M. Placental Protein Expression of Kisspeptin-1 (KISS1) and the Kisspeptin-1 Receptor (KISS1R) in Pregnancy Complicated by Diabetes Mellitus or Preeclampsia. Arch. Gynecol. Obstet. 2020, 301, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Tsoutsouki, J.; Patel, B.; Comninos, A.N.; Dhillo, W.S. Kisspeptin in the Prediction of Pregnancy Complications. Front. Endocrinol. 2022, 13, 942664. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, H.; Eyuboglu, S.; Erkanlı Senturk, G.; Yilmaz, B. Investigation of the Effects of Kisspeptin-10 in Methionine-Induced Lipid Peroxidation in Testicle Tissue of Young Rats. J. Biochem. Mol. Toxicol. 2017, 31, e21881. [Google Scholar] [CrossRef]
- Aslan, M.; Erkanli Senturk, G.; Akkaya, H.; Sahin, S.; Yılmaz, B. The Effect of Oxytocin and Kisspeptin-10 in Ovary and Uterus of Ischemia-Reperfusion Injured Rats. Taiwan. J. Obstet. Gynecol. 2017, 56, 456–462. [Google Scholar] [CrossRef]
- Aydin, M.; Oktar, S.; Yonden, Z.; Ozturk, O.H.; Yilmaz, B. Direct and Indirect Effects of Kisspeptin on Liver Oxidant and Antioxidant Systems in Young Male Rats. Cell Biochem. Funct. 2010, 28, 293–299. [Google Scholar] [CrossRef]
- Gorbunova, O.L.; Shirshev, S.V. Role of Kisspeptin in Regulation of Reproductive and Immune Reactions. Biochem. 2020, 85, 839–853. [Google Scholar] [CrossRef]
- Zahid, A.; Li, B.; Kombe, A.J.K.; Jin, T.; Tao, J. Pharmacological Inhibitors of the Nlrp3 Inflammasome. Front. Immunol. 2019, 10, 2538. [Google Scholar] [CrossRef]
- Dixon, L.J.; Flask, C.A.; Papouchado, B.G.; Feldstein, A.E.; Nagy, L.E. Caspase-1 as a Central Regulator of High Fat Diet-Induced Non-Alcoholic Steatohepatitis. PLoS ONE 2013, 8, e56100. [Google Scholar] [CrossRef]
- Kotas, M.E.; Jurczak, M.J.; Annicelli, C.; Gillum, M.P.; Cline, G.W.; Shulman, G.I.; Medzhitov, R. Role of Caspase-1 in Regulation of Triglyceride Metabolism. Proc. Natl. Acad. Sci. USA 2013, 110, 4810–4815. [Google Scholar] [CrossRef]
- Bowe, J.E.; King, A.J.; Kinsey-Jones, J.S.; Foot, V.L.; Li, X.F.; O’Byrne, K.T.; Persaud, S.J.; Jones, P.M. Kisspeptin Stimulation of Insulin Secretion: Mechanisms of Action in Mouse Islets and Rats. Diabetologia 2009, 52, 855–862. [Google Scholar] [CrossRef]
- Bowe, J.E.; Foot, V.L.; Amiel, S.A.; Huang, G.C.; Lamb, M.; Lakey, J.; Jones, P.M.; Persaud, S.J. GPR54 Peptide Agonists Stimulate Insulin Secretion from Murine, Porcine and Human Islets. Islets 2012, 4, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Bowe, J.E.; Hill, T.G.; Hunt, K.F.; Smith, L.I.F.; Simpson, S.J.S.; Amiel, S.A.; Jones, P.M. A Role for Placental Kisspeptin in β Cell Adaptation to Pregnancy. JCI Insight 2019, 4, e124540. [Google Scholar] [CrossRef] [PubMed]
- Izzi-Engbeaya, C.; Comninos, A.N.; Clarke, S.A.; Jomard, A.; Yang, L.; Jones, S.; Abbara, A.; Narayanaswamy, S.; Eng, P.C.; Papadopoulou, D.; et al. The Effects of Kisspeptin on β-Cell Function, Serum Metabolites and Appetite in Humans. Diabetes Obes. Metab. 2018, 20, 2800–2810. [Google Scholar] [CrossRef] [PubMed]
- Schwetz, T.A.; Reissaus, C.A.; Piston, D.W. Differential Stimulation of Insulin Secretion by Glp-1 and Kisspeptin-10. PLoS ONE 2014, 9, e113020. [Google Scholar] [CrossRef] [PubMed]
- Wahab, F.; Riaz, T.; Shahab, M. Study on the Effect of Peripheral Kisspeptin Administration on Basal and Glucose-Induced Insulin Secretion under Fed and Fasting Conditions in the Adult Male Rhesus Monkey (Macaca Mulatta). Horm. Metab. Res. 2011, 43, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Molla, M.D.; Ayelign, B.; Dessie, G.; Geto, Z.; Admasu, T.D. Caspase-1 as a Regulatory Molecule of Lipid Metabolism. Lipids Health Dis. 2020, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Ilie, M.; Khambata-Ford, S.; Copie-Bergman, C.; Huang, L.; Juco, J.; Hofman, V.; Hofman, P. Use of the 22C3 Anti–PD-L1 Antibody to Determine PD-L1 Expression in Multiple Automated Immunohistochemistry Platforms. PLoS ONE 2017, 12, e0183023. [Google Scholar] [CrossRef]
- Solano, M.E.; Thiele, K.; Kowal, M.K.; Arck, P.C. Identification of Suitable Reference Genes in the Mouse Placenta. Placenta 2016, 39, 7–15. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′->3′) | Accession Number |
|---|---|---|
| Il18 | F: ACCACTTTGGCAGACTTCACT | NM_019165.1 |
| R: ACACAGGCGGGTTTCTTTTG | ||
| Il1β | F: GCACAGTTCCCCAACTGGTA | NM_031512.2 |
| R: TGTCCCGACCATTGCTGTTT | ||
| Nlrp3 | F: CTCTGCATGCCGTATCTGGT | NM_001191642.1 |
| R: GTCCTGAGCCATGGAAGCAA | ||
| Casp1 | F: ACAAAGAAGGTGGCGCATTT | NM_012762.2 |
| R: GTGCTGCAGATAATGAGGGC | ||
| Gsmd | F: AAGATCGTGGATCATGCCGT | NM_001130553.1 |
| F: AAGATCGTGGATCATGCCGT | ||
| Polr2a | F: GCTGGACCTACTGGCATGTT | XM_001079162.5 |
| R: ACCATAGGCTGGAGTTGCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, B.R.; Cordeiro, J.M.d.A.; Santos, L.C.; Santana, L.d.S.; Nascimento, A.E.d.J.; Silva, J.F. Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats. Int. J. Mol. Sci. 2023, 24, 6820. https://doi.org/10.3390/ijms24076820
Santos BR, Cordeiro JMdA, Santos LC, Santana LdS, Nascimento AEdJ, Silva JF. Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats. International Journal of Molecular Sciences. 2023; 24(7):6820. https://doi.org/10.3390/ijms24076820
Chicago/Turabian StyleSantos, Bianca Reis, Jeane Martinha dos Anjos Cordeiro, Luciano Cardoso Santos, Larissa da Silva Santana, Acácia Eduarda de Jesus Nascimento, and Juneo Freitas Silva. 2023. "Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats" International Journal of Molecular Sciences 24, no. 7: 6820. https://doi.org/10.3390/ijms24076820
APA StyleSantos, B. R., Cordeiro, J. M. d. A., Santos, L. C., Santana, L. d. S., Nascimento, A. E. d. J., & Silva, J. F. (2023). Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats. International Journal of Molecular Sciences, 24(7), 6820. https://doi.org/10.3390/ijms24076820