
Unraveling the Role of Antimicrobial Peptides in Insects

Abstract
1. Introduction
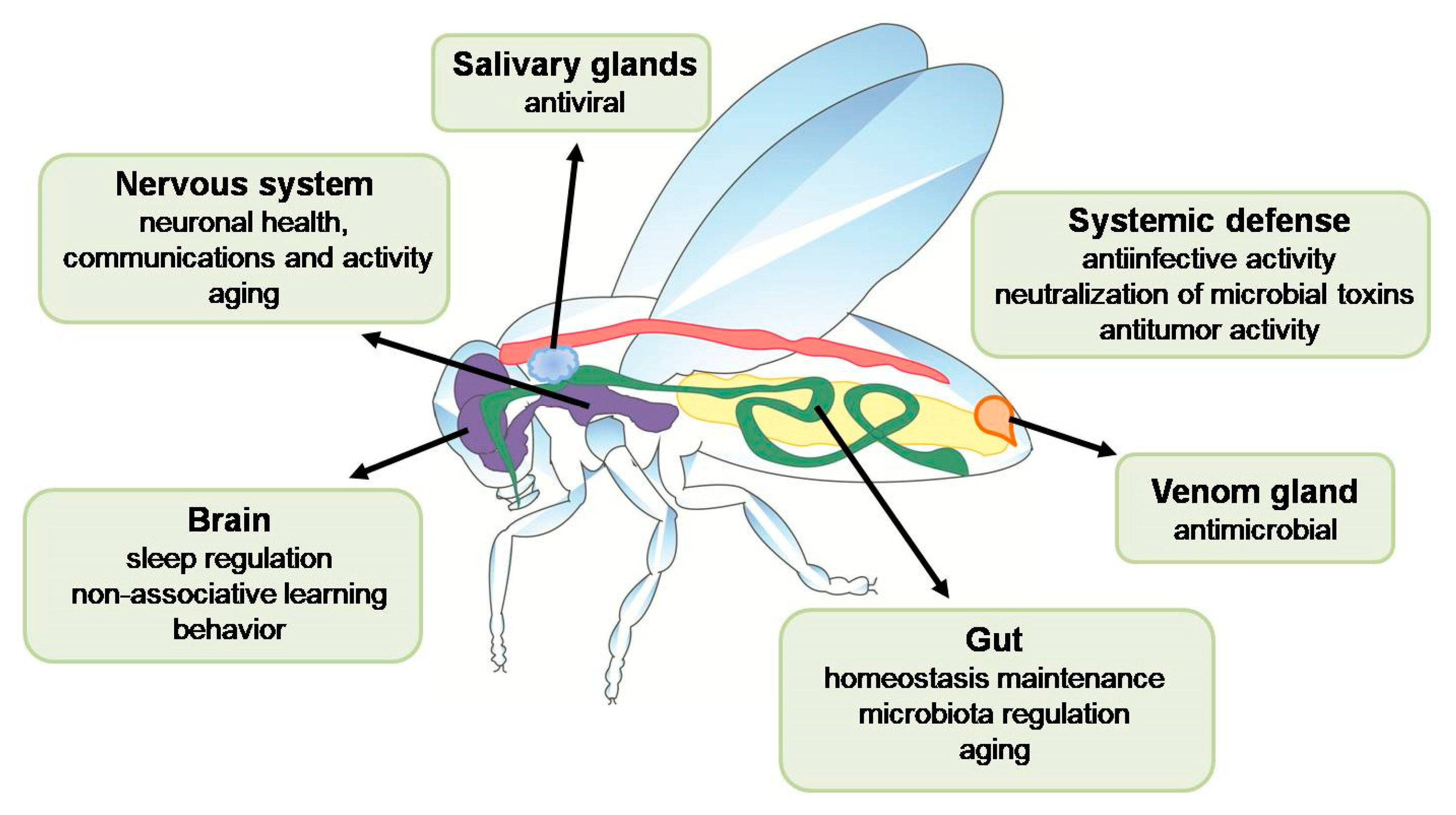
2. AMPs in the Nervous System
3. AMPs in Aging
4. AMPs as Antitumor Agents
5. AMPs and Symbiotic Microbiota

{kind=link}
{kind=link}
| Order | Host Insect | Symbionts | Reference |
|---|---|---|---|
| Lepidoptera | Bombyx mori | Proteus vulgaris, Erwinia sp. Klebsiella pneumoniae Citobacter freundii Pseudomonas fluorescens | [125] |
| Galleria mellonella | Streptococcus faecalis (Enterecoccus mundii) | [99,126] | |
| Plutella xylostella | Enterococcus sp. Enterobacter sp., Serratia sp. | [127] | |
| Spodoptera litura | Serratia sp. | [128] | |
| Diptera | Aedes aegypti | Wolbachia | [129] |
| Anopheles gambiae | Enterobacter asburiae, Serratia sp. Microbacterium sp. Sphingomonas sp. Chryseobacterium meningosepticum | [130] | |
| Drosophila melanogaster | Spiroplasma poulsonii Acetobacter thailandicus Lactobacillus plantarum | [96,131,132,133] | |
| Drosophila nebulosa | Spiroplasma poulsonii | [124,134] | |
| Glossinia spp. | Sodalis glossimidia Wigglesworthia sp. Wigglesworthia glossinia | [79,135,136,137,138] | |
| Melophagus ovinus | Arsenophonus melophagi Sodalis melophagi | [139,140] | |
| Hymenoptera | Apis mellifera | Gilliamella apicola, Snodgrassiella sp. Frischella perrara, Snodgrassella alvi Bartonella apis | [102,103,141,142,143,144,145,146] |
| Bombus spp. | Giliamella bombicola Snodgrassiella sp. | [146,147] | |
| Camponotus floridanus | Blochmannia floridanus | [112] | |
| Coleoptera | Acyrthosiphon pisum | Buchnera aphidicola | [148,149,150] |
| Cyrtotrachelus buqueti | Lactococcus sp., Serratia sp. Dysgonomonas sp., Enterrococus sp. | [151] | |
| Holotrichia parallela | Pseudomonas sp., Ochrobacterium sp. Cellulosimicrobrium sp. | [152] | |
| Hylobius abietis | Erwinia sp., Rabnella sp., Serratia sp. | [153,154] | |
| Nicrophorus vespilloides | Providencia sp., Morganella sp. Vagococcus sp., Proteus sp. Koukoulia sp., Serratia sp. | [155,156,157] | |
| Pachyrhynchus infernalis | Nardonella sp. | [123] | |
| Paradieuches dissimilis | Caballeronia, Symbiopectobacterium Wolbachia, Rickettsiella | [158] | |
| Riptortus pedestris | Burkholderia | [159,160] | |
| Sirex noctilio | Streptomyces sp. | [161] | |
| Sitophilus oryze | Sodalis pierantonius | [162] | |
| Sitophilus weevils | Sodalis pierantonius | [163] | |
| Blattodea | Blattella germanica | Blattabacterium cuenoti | [164] |
6. Non-Classical AMPs and Infection
7. AMPs in Antiviral Response
8. AMPs in Insect Venoms
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Steiner, H.; Hultmark, D.; Engström, A.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef]
- Cytryńska, M.; Mak, P.; Zdybicka-Barabas, A.; Suder, P.; Jakubowicz, T. Purification and characterization of eight peptides from Galleria mellonella immune hemolymph. Peptides 2007, 28, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Nesa, J.; Sadat, A.; Buccini, D.F.; Kati, A.; Mandal, A.K.; Franco, O.L. Antimicrobial peptides from Bombyx mori: A splendid immune defense response in silkworms. RSC Adv. 2020, 10, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I.; Cytryńska, M.; Zdybicka-Barabas, A.; Kordaczuk, J. Insect defense proteins and peptides. Subcell. Biochem. 2020, 94, 81–121. [Google Scholar]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Cytryńska, M.; Zdybicka-Barabas, A. Defense peptides: Recent developments. Biomol. Concepts 2015, 6, 237–251. [Google Scholar] [CrossRef]
- Malanovic, N.; Marx, L.; Blondelle, S.E.; Pabst, G.; Semeraro, E.F. Experimental concepts for linking the biological activities of antimicrobial peptides to their molecular modes of action. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183275. [Google Scholar] [CrossRef]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A review on antimicrobial peptides databases and the computational tools. Database 2022, 2022, baac011. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Zhang, W.; Heryanto, C.; Mohamed, A.; Contreras, G.; Tettamanti, G.; Wink, M.; Bassal, T. Diversity of insect antimicrobial peptides and proteins—A functional perspective: A review. Int. J. Biol. Macromol. 2021, 191, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Song, Y. Mechanism of antimicrobial peptides: Antimicrobial, anti-inflammatory and antibiofilm activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef]
- Hafeez, A.B.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial peptides: An update on classifications and databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Yount, N.Y.; Weaver, D.C.; Lee, E.Y.; Lee, M.W.; Wang, H.; Chan, L.C.; Wong, G.C.L.; Yeaman, M.R. Unifying structural signature of eukaryotic α-helical host defense peptides. Proc. Natl. Acad. Sci. USA 2019, 116, 6944–6953. [Google Scholar] [CrossRef]
- Yount, N.Y.; Yeaman, M.R. Multidimensional signatures in antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 7363–7368. [Google Scholar] [CrossRef] [PubMed]
- Kordaczuk, J.; Sułek, M.; Wojda, I. General overview on the role of Peptidoglycan Recognition Proteins in insect immunity. Acta Biochim. Pol. 2020, 67, 319–326. [Google Scholar] [CrossRef]
- Charroux, B.; Royet, J. Drosophila immune response: From systemic antimicrobial peptide production in fat body cells to local defense in the intestinal tract. Fly 2010, 4, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tettamanti, G.; Bassal, T.; Heryanto, C.; Eleftherianos, I.; Mohamed, A. Regulators and signalling in insect antimicrobial innate immunity: Functional molecules and cellular pathways. Cell. Signal. 2021, 83, 110003. [Google Scholar] [CrossRef]
- Bai, S.; Yao, Z.; Raza, M.F.; Cai, Z.; Zhang, H. Regulatory mechanisms of microbial homeostasis in insect gut. Insect Sci. 2021, 28, 286–301. [Google Scholar] [CrossRef]
- Zhai, Z.; Huang, X.; Yin, Y. Beyond immunity: The Imd pathway as a coordinator of host defense, organismal physiology and behavior. Dev. Comp. Immunol. 2018, 83, 51–59. [Google Scholar] [CrossRef]
- Bland, M.L. Regulating metabolism to shape immune function: Lessons from Drosophila. Semin. Cell Dev. Biol. 2023, 138, 128–141. [Google Scholar] [CrossRef]
- Manniello, M.D.; Moretta, A.; Salvia, R.; Scieuzo, C.; Lucchetti, D.; Vogel, H.; Sgambato, A.; Falabella, P. Insect antimicrobial peptides: Potential weapons to counteract the antibiotic resistance. Cell Mol. Life Sci. 2021, 78, 4259–4282. [Google Scholar] [CrossRef]
- Mangold, C.A.; Hughes, D.P. Insect behavioral change and the potential contributions of neuroinflammation—A call for future research. Genes 2021, 12, 465. [Google Scholar] [CrossRef]
- Stuart, B.A.R.; Franitza, A.L.; Lezi, E. Regulatory roles of antimicrobial peptides in the nervous system: Implications for neuronal aging. Front. Cell Neurosci. 2022, 16, 843790. [Google Scholar] [CrossRef]
- Montanari, M.; Royet, J. Impact of microorganisms and parasites on neuronally controlled Drosophila behaviours. Cells 2021, 10, 2350. [Google Scholar] [CrossRef] [PubMed]
- van Alphen, B.; Stewart, S.; Iwanaszko, M.; Xu, F.; Li, K.; Rozenfeld, S.; Ramakrishnan, A.; Itoh, T.Q.; Sisobhan, S.; Qin, Z.; et al. Glial immune-related pathways mediate effects of closed head traumatic brain injury on behavior and lethality in Drosophila. PLoS Biol. 2022, 20, e3001456. [Google Scholar] [CrossRef] [PubMed]
- Dissel, S.; Seugnet, L.; Thimgan, M.S.; Silverman, N.; Angadi, V.; Thacher, P.V.; Burnham, M.M.; Shaw, P.J. Differential activation of immune factors in neurons and glia contribute to individual differences in resilience/vulnerability to sleep disruption. Brain Behav. Immun. 2015, 47, 75–85. [Google Scholar] [CrossRef]
- Toda, H.; Williams, J.A.; Gulledge, M.; Sehgal, A. A sleep-inducing gene, nemuri, links sleep and immune function in Drosophila. Science 2019, 363, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Bozler, J.; Kacsoh, B.Z.; Chen, H.; Theurkauf, W.E.; Weng, Z.; Bosco, G. A systems level approach to temporal expression dynamics in Drosophila reveals clusters of long term memory genes. PLoS Genet. 2017, 13, e1007054. [Google Scholar] [CrossRef] [PubMed]
- Barajas-Azpeleta, R.; Wu, J.; Gill, J.; Welte, R.; Seidel, C.; McKinney, S.; Dissel, S.; Si, K. Antimicrobial peptides modulate long-term memory. PLoS Genet. 2018, 14, e1007440. [Google Scholar] [CrossRef]
- Shaik, H.A.; Mishra, A.; Sehadová, H.; Kodrík, D. Responses of sericotropin to toxic and pathogenic challenges: Possible role in defense of the wax moth Galleria mellonella. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 227, 108633. [Google Scholar] [CrossRef]
- Pizzorno, M.C.; Field, K.; Kobokovich, A.L.; Martin, P.L.; Gupta, R.A.; Mammone, R.; Rovnyak, D.; Capaldi, E.A. Transcriptomic responses of the honey bee brain to infection with deformed wing virus. Viruses 2021, 13, 287. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; Keyhani, N.O.; Jin, K.; Wei, Q.; Xia, Y. Central nervous system responses of the oriental migratory, Locusta migratoria manilensis, to fungal infection. Sci. Rep. 2017, 7, 10340. [Google Scholar] [CrossRef]
- Katzenberger, R.J.; Loewen, C.A.; Wassarman, D.R.; Petersen, A.J.; Ganetzky, B.; Wassarman, D.A. A Drosophila model of closed head traumatic brain injury. Proc. Natl. Acad. Sci. USA 2013, 110, E4152–E4159. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.J.; Katzenberger, R.J.; Wassarman, D.A. The innate immune response transcription factor relish is necessary for neurodegeneration in a Drosophila model of ataxia-telangiectasia. Genetics 2013, 194, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Bolus, H.; Crocker, K.; Boekhoff-Falk, G.; Chtarbanova, S. Modeling neurodegenerative disorders in Drosophila melanogaster. Int. J. Mol. Sci. 2020, 21, 3055. [Google Scholar] [CrossRef]
- Nayak, N.; Mishra, M. Drosophila melanogaster as a model to understand the mechanisms of infection mediated neuroinflammation in neurodegenerative diseases. J. Integr. Neurosci. 2022, 21, 66. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Luo, F.; Xu, Y.; Zhang, Y.; Jin, L.H. Drosophila innate immunity involves multiple signaling pathways and coordinated communication between different tissues. Front. Immunol. 2022, 13, 905370. [Google Scholar] [CrossRef]
- Cao, Y.; Chtarbanova, S.; Petersen, A.J.; Ganetzky, B. Dnr1 mutations cause neurodegeneration in Drosophila by activating the innate immune response in the brain. Proc. Natl. Acad. Sci. USA 2013, 110, E1752–E1760. [Google Scholar] [CrossRef] [PubMed]
- Myllymäki, H.; Rämet, M. Transcription factor zfh1 downregulates Drosophila Imd pathway. Dev. Comp. Immunol. 2013, 39, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Kounatidis, I.; Chtarbanova, S.; Cao, Y.; Hayne, M.; Jayanth, D.; Ganetzky, B.; Ligoxygakis, P. NF-κB immunity in the brain determines fly lifespan in healthy aging and age-related neurodegeneration. Cell Rep. 2017, 19, 836–848. [Google Scholar] [CrossRef]
- Barati, A.; Masoudi, R.; Yousefi, R.; Mosefi, M.; Mirshafiey, A. Tau and amyloid beta differentially affect the innate immune genes expression in Drosophila models of Alzheimer’s disease and b-D monnuroic acid (M2000) modulates the dysregulation. Gene 2022, 808, 145972–145984. [Google Scholar] [CrossRef]
- Dubey, S.K.; Tapadia, M.G. Yorkie regulates neurodegeneration through canonical pathway and innate immune response. Mol. Neurobiol. 2018, 55, 1193–1207. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.C.; Rimkus, S.A.; Ganetzky, B.; Wassarman, D.A. Loss of the antimicrobial peptide metchnikowin protects against traumatic brain injury outcomes in Drosophila melanogaster. G3 Genes Genomes Genet. 2020, 10, 3109–3119. [Google Scholar] [CrossRef]
- Wang, M.; Peng, I.F.; Li, S.; Hu, X. Dysregulation of antimicrobial peptide expression distinguishes Alzheimer’s disease from normal aging. Aging 2020, 12, 690–706. [Google Scholar] [CrossRef] [PubMed]
- Sandin, L.; Bergkvist, L.; Nath, S.; Kielkopf, C.; Janefjord, C.; Helmfors, L.; Zetterberg, H.; Blennow, K.; Li, H.; Nilsberth, C.; et al. Beneficial effects of increased lysozyme levels in Alzheimer’s disease modelled in Drosophila melanogaster. FEBS J. 2016, 283, 3508–3522. [Google Scholar] [CrossRef] [PubMed]
- Vojtechova, I.; Machacek, T.; Kristofikova, Z.; Stuchlik, A.; Petrasek, T. Infectious origin of Alzheimer’s disease: Amyloid beta as a component of brain antimicrobial immunity. PLoS Pathog. 2022, 18, e1010929. [Google Scholar] [CrossRef]
- Sudmeier, L.J.; Samudrala, S.S.; Howard, S.P.; Ganetzky, B. Persistent activation of the innate immune response in adult Drosophila following radiation exposure during larval development. G3 Genes Genomes Genet. 2015, 5, 2299–2306. [Google Scholar]
- Schluesener, H.J.; Su, Y.; Ebrahimi, A.; Pouladsaz, D. Antimicrobial peptides in the brain: Neuropeptides and amyloid. Front. Biosci. 2012, 4, 1375–1380. [Google Scholar] [CrossRef]
- Augustyniak, D.; Kramarska, E.; Mackiewicz, P.; Orczyk-Pawiłowicz, M.; Lundy, F.T. Mammalian Neuropeptides as Modulators of Microbial Infections: Their dual role in defense versus virulence and pathogenesis. Int. J. Mol. Sci. 2021, 22, 3658. [Google Scholar] [CrossRef]
- Lee, E.Y.; Chan, L.C.; Wang, H.; Lieng, J.; Hung, M.; Srinivasan, Y.; Wang, J.; Waschek, J.A.; Ferguson, A.L.; Lee, K.F.; et al. PACAP is a pathogen-inducible resident antimicrobial neuropeptide affording rapid and contextual molecular host defense of the brain. Proc. Natl. Acad. Sci. USA 2021, 118, e1917623117. [Google Scholar] [CrossRef]
- Piper, M.D.W.; Partridge, L. Drosophila as a model for ageing. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2707–2717. [Google Scholar] [CrossRef]
- Garschall, K.; Flatt, T. The interplay between immunity and aging in Drosophila. F1000Res 2018, 7, 160. [Google Scholar] [CrossRef]
- Maruzs, T.; Simon-Vecsei, Z.; Kiss, V.; Csizmadia, T.; Juhász, G. On the fly: Recent progress on autophagy and aging in Drosophila. Front. Cell Dev. Biol. 2019, 7, 140. [Google Scholar] [CrossRef]
- Lai, C.-Q.; Parnell, L.D.; Lyman, R.F.; Ordovas, J.M.; Mackay, T.F.C. Candidate genes affecting Drosophila life span identified by integrating microarray gene expression analysis and QTL mapping. Mech. Ageing Dev. 2007, 28, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Aggarwal, K.; Paquette, N.; Silverman, N. NF-κB/Rel proteins and the humoral immune responses of Drosophila melanogaster. NF-Kb Health Dis. 2011, 349, 25–60. [Google Scholar]
- Badinloo, M.; Nguyen, E.; Suh, W.; Alzahrani, F.; Castellanos, J.; Klichko, V.I.; Orr, W.C.; Radyuk, S.N. Overexpression of antimicrobial peptides contributes to aging through cytotoxic effects in Drosophila tissues. Arch. Insect Biochem. Physiol. 2018, 98, e21464. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.I.; Honda, D.; Okumura, M.; Miura, M.; Chihara, T. Systemic innate immune response induces death of olfactory receptor neurons in Drosophila. Genes Cells 2022, 27, 113–123. [Google Scholar] [CrossRef]
- Li, J.; Terry, E.E.; Fejer, E.; Gamba, D.; Hartmann, N.; Logsdon, J.; Michalski, D.; Rois, L.E.; Scuderi, M.J.; Kunst, M.; et al. Achilles is a circadian clock-controlled gene that regulates immune function in Drosophila. Brain Behav. Immun. 2017, 61, 127–136. [Google Scholar] [CrossRef]
- Loch, G.; Zinke, I.; Mori, T.; Carrera, P.; Schroer, J.; Takeyama, H.; Hoch, M. Antimicrobial peptides extend lifespan in Drosophila. PLoS ONE 2017, 12, e0176689. [Google Scholar] [CrossRef]
- Zhao, H.W.; Zhou, D.; Nizet, V.; Haddad, G.G. Experimental selection for Drosophila survival in extremely high O2 environments. PLoS ONE 2010, 5, e11701. [Google Scholar] [CrossRef]
- Zhao, H.W.; Zhou, D.; Haddad, G.G. Antimicrobial peptides increase tolerance to oxidant stress in Drosophila melanogaster. J. Biol. Chem. 2011, 286, 6211–6218. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Lemaitre, B.; Unckless, R.L. Dynamic evolution of antimicrobial peptides underscores trade-offs between immunity and ecological fitness. Front. Immunol. 2019, 10, 2620. [Google Scholar] [CrossRef]
- Carboni, A.L.; Hanson, M.A.; Lindsay, S.A.; Wasserman, S.A.; Lemaitre, B. Cecropins contribute to Drosophila host defense against a subset of fungal and Gram-negative bacterial infection. Genetics 2022, 220, iyab188. [Google Scholar] [CrossRef]
- Belmonte, R.L.; Corbally, M.K.; Duneau, D.F.; Regan, J.C. Sexual dimorphisms in innate immunity and responses to infection in Drosophila melanogaster. Front. Immunol. 2020, 10, 3075. [Google Scholar] [CrossRef]
- Shit, B.; Prakash, A.; Sarkar, S.; Vale, P.F.; Khan, I. Ageing leads to reduced specificity of antimicrobial peptide responses in Drosophila melanogaster. Proc. Biol. Sci. 2022, 289, 20221642. [Google Scholar]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A.C.; Carrera, P.; Inhester, T.; Schultze, J.L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 436, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Fink, C.; Hoffmann, J.; Knop, M.; Li, Y.; Isermann, K.; Roeder, T. Intestinal FoxO signaling is required to survive oral infection in Drosophila. Mucosal Immunol. 2016, 9, 927–936. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, W.; Xu, J.; Yang, W.; Li, Q.; Zhong, Y.; Cao, Y.; Yu, X.Q.; Deng, X. Regulation of antimicrobial peptide genes via insulin-like signaling pathway in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2018, 103, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Varma, D.; Bülow, M.H.; Pesch, Y.Y.; Loch, G.; Hoch, M. Forkhead, a new cross regulator of metabolism and innate immunity downstream of TOR in Drosophila. J. Insect Physiol. 2014, 69, 80–88. [Google Scholar] [CrossRef]
- Suttmann, H.; Retz, M.; Paulsen, F.; Harder, J.; Zwergel, U.; Kamradt, J.; Wullich, B.; Unteregger, G.; Stöckle, M.; Lehmann, J. Antimicrobial peptides of the Cecropin-family show potent antitumor activity against bladder cancer cells. BMC Urol. 2008, 8, 5. [Google Scholar] [CrossRef]
- Kao, F.S.; Pan, Y.R.; Hsu, R.Q.; Chen, H.M. Efficacy verification and microscopic observations of an anticancer peptide, CB1a, on single lung cancer cell. Biochim. Biophys. Acta 2012, 1818, 2927–2935. [Google Scholar] [CrossRef]
- Tonk, M.; Vilcinskas, A.; Rahnamaeian, M. Insect antimicrobial peptides: Potential tools for the prevention of skin cancer. Appl. Microbiol. Biotechnol. 2016, 100, 7397–7405. [Google Scholar] [CrossRef]
- Ramos-Martín, F.; Herrera-León, C.; D’Amelio, N. Bombyx mori Cecropin D could trigger cancer cell apoptosis by interacting with mitochondrial cardiolipin. Biochim. Biophys. Acta. Biomembr. 2022, 1864, 184003. [Google Scholar] [CrossRef] [PubMed]
- Parvy, J.P.; Yu, Y.; Dostalova, A.; Kondo, S.; Kurjan, A.; Bulet, P.; Lemaitre, B.; Vidal, M.; Cordero, J.B. The antimicrobial peptide defensin cooperates with tumour necrosis factor to drive tumour cell death in Drosophila. eLife 2019, 8, e45061. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Kurihara, M.; Kinoshita, S.; Awane, R.; Sato, T.; Ohkawa, Y.; Inoue, Y.H. Anti-tumour effects of antimicrobial peptides, components of the innate immune system, against haematopoietic tumours in Drosophila mxc mutants. Dis. Model. Mech. 2019, 12, dmm037721. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, S.; Takarada, K.; Kinoshita, Y.; Inoue, Y.H. Drosophila hemocytes recognize lymph gland tumors of mxc mutants and activate the innate immune pathway in a reactive oxygen species-dependent manner. Biol. Open. 2022, 11, bio059523. [Google Scholar] [CrossRef]
- Jacqueline, C.; Parvy, J.P.; Rollin, M.L.; Faugère, D.; Renaud, F.; Missé, D.; Thomas, F.; Roche, B. The role of innate immunity in the protection conferred by a bacterial infection against cancer: Study of an invertebrate model. Sci. Rep. 2020, 10, 10106. [Google Scholar] [CrossRef]
- Zaidman-Rémy, A.; Vigneron, A.; Weiss, B.L.; Heddi, A. What can a weevil teach a fly, and reciprocally? Interaction of host immune systems with endosymbionts in Glossina and Sitophilus. BMC Microbiol. 2018, 18, 279–292. [Google Scholar] [CrossRef]
- Mateos, M.; Castrezana, S.J.; Nankivell, B.J.; Estes, A.M.; Markow, T.A.; Moran, N.A. Heritable endosymbionts of Drosophila. Genetics 2006, 174, 363–376. [Google Scholar] [CrossRef]
- Gross, R.; Vavre, F.; Heddi, A.; Hurst, G.D.; Zchori-Fein, E.; Bourtzis, K. Immunity and symbiosis. Mol. Microbiol. 2009, 73, 751–759. [Google Scholar] [CrossRef]
- Douglas, A.E. Lessons from studying insect symbiosis. Cell Host Microbe 2011, 10, 359–367. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. An ancient but promiscuous host-symbiont association between Burkholderia gut symbionts and their heteropteran hosts. ISME J. 2011, 5, 446–460. [Google Scholar] [CrossRef]
- Weiss, B.; Aksoy, S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011, 27, 514–522. [Google Scholar] [CrossRef]
- Kuraishi, T.; Hori, A.; Kurata, S. Host-microbe interactions in the gut of Drosophila melanogaster. Front. Physiol. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-A.; Lee, W.-J. Drosophila as a model for intestinal dysbiosis and chronic inflammatory diseases. Dev. Comp. Immunol. 2014, 42, 102–110. [Google Scholar] [CrossRef]
- Masson, F.; Zaidman-Rémy, A.; Heddi, A. Antimicrobial peptides and cell processes tracking endosymbiont dynamics. Philos. Trans. R. Soc. B 2016, 371, 20150298. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Engel, P. Mechanisms underlying gut microbiota-host interactions in insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial word: Insights from Drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef]
- Margaert, P. Role of antimicrobial peptides in controlling symbiotic bacterial populations. Nat. Prod. Rep. 2018, 35, 336–356. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffman, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Bosco-Drayon, V.; Poidevin, M.; Boneca, I.G.; Narbonne-Reveau, K.; Royet, J.; Charroux, B. Peptidoglican sensing by the receptor PGRP-LE in the Drosophila melanogaster gut induces immune response to infectious bacteria and tolerance to microbiota. Cell. Host Microbe 2012, 12, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Broderic, N.A.; Buchon, N.; Lemaitre, B. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. mBio 2014, 5, e01117-14. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Hanson, M.A.; Kondo, S.; Erkosar, B.; Lemaitre, B. Drosophila antimicrobial peptides and lysozyme regulate gut microbiota composition and abudance. mBio 2021, 12, e00824-21. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Jaffar, S.; Xu, Y.; Qi, Y. The intestinal immune defense system in insects. Int. J. Mol. Sci. 2022, 23, 15132. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Kim, S.H.; Lee, H.Y.; Bai, J.Y.; Nam, Y.D.; Bae, J.W.; Lee, D.G.; Shin, S.C.; Ha, E.M.; Lee, W.J. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef]
- Clayton, A.M.; Cirimotich, C.M.; Dong, Y.; Dimopoulos, G. Caudal is a negative regulator of the Anopheles IMD pathway that controls resistance to P. falciparum infection. Dev. Comp. Immunol. 2013, 39, 323–332. [Google Scholar] [CrossRef]
- Sarvari, M.; Mikani, A.; Mehrabadi, M. The innate immune gene Relish and Caudal jointly contribute to the gut immune homeostasis by regulating antimicrobial peptides in Galleria mellonella. Dev. Comp. Immunol. 2020, 110, 103732. [Google Scholar] [CrossRef]
- Jarosz, J. Gut flora of Galleria mellonella suppressing ingested bacteria. J. Invertebr. Pathol. 1979, 34, 192–198. [Google Scholar] [CrossRef]
- Gohl, P.; LeMoine, C.M.; Cassone, B.J. Diet and ontogeny drastically alter the larval microbiome of the invertebrate model Galleria mellonella. Can. J. Microbiol. 2022, 68, 594–604. [Google Scholar] [CrossRef]
- Krams, I.A.; Kecko, S.; Jõers, P.; Trakimas, G.; Elferts, D.; Krams, R.; Luoto, S.; Rantala, M.J.; Inashkina, I.; Gudrā, D.; et al. Microbiome symbionts and diet diversity incur costs on the immune system of insect larvae. J. Exp. Biol. 2017, 220, 4204–4212. [Google Scholar] [CrossRef]
- Emery, O.; Schmit, K.; Engel, P. Immune system stimulation by the gut symbiont Frischella perrara in the honey bee (Apis mellifera). Mol. Ecol. 2017, 26, 2576–2590. [Google Scholar] [CrossRef]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, A.N.K.G.; Nair, A.J.; Sugunan, V.S. A review on royal jelly proteins and peptides. J. Funct. Foods 2018, 44, 255–264. [Google Scholar] [CrossRef]
- Collazo, N.; Carpena, M.; Nuñez-estevez, B.; Otero, P.; Simal-Gandara, J.; Prieto, M.A. Health promoting properties of bee royal jelly: Food of the queens. Nutrients 2021, 13, 543. [Google Scholar] [CrossRef]
- Bílikova, K.; Huang, S.-C.; Linb, I.-P.; Simuth, J.; Peng, C.-C. Peptides structure and antimicrobial activity relationship of royalisin, an antimicrobial peptide from royal jelly of Apis mellifera. Peptides 2015, 68, 190–196. [Google Scholar] [CrossRef]
- Jia, F.; Wang, J.; Peng, J.; Zhao, P.; Kong, Z.; Wang, K.; Yan, W.; Wang, R. The in vitro, in vivo antifungal activity and the action mode of Jelleine-I against Candida species. Amino Acids 2018, 50, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Li, M.; Guo, H.; Peng, W.; Xue, X.; Hu, Y.; Liu, Y.; Zhao, Y.; Fang, X.; Wang, K.; et al. Architecture of the native major royal jelly protein 1 oligomer. Nat. Commun. 2018, 9, 3373. [Google Scholar] [CrossRef]
- Jia, F.; Wang, J.; Zhang, L.; Zhou, J.; He, Y.; Lu, Y.; Liu, K.; Yan, W.; Wang, K. Multiple action mechanism and in vivo antimicrobial efficacy of antimicrobial peptide Jelleine-I. J. Pept. Sci. 2021, 27, e3294. [Google Scholar] [CrossRef]
- de Souza, D.J.; Bézier, A.; Depoix, D.; Drezen, J.M.; Lenoir, A. Blochmannia endosymbionts improve colony growth and immune defence in the ant Camponotus fellah. BMC Microbiol. 2009, 9, 29. [Google Scholar] [CrossRef]
- Sinotte, V.M.; Freedman, S.N.; Ugelvig, L.V.; Seid, M.A. Camponotus floridanus ants incur a trade-off between phenotypic development and pathogen susceptibility from their mutualistic endosymbiont Blochmannia. Insects 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Ratzka, C.; Gross, R.; Feldhaar, H. Gene expression analysis of the endosymbiont-bearing midgut tissue during ontogeny of the carpenter ant Camponotus floridanus. J. Insect Physiol. 2013, 59, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Cai, Z.; Ma, Q.; Bai, S.; Wang, Y.; Zhang, P.; Guo, Q.; Gu, J.; Lemaitre, B.; Zhang, H. Compartmentalized PGRP expression along the dipteran Bactrocera dorsalis gut forms a zone of protection for symbiotic bacteria. Cell Rep. 2022, 41, 111523. [Google Scholar] [CrossRef]
- Login, F.H.; Balmand, S.; Vallier, A.; Vincent-Monégat, C.; Vigneron, A.; Weiss-Gayet, M.; Rochat, D.; Heddi, A. Antimicrobial peptides keep insect endosymbionts under control. Science 2011, 334, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Maire, J.; Vincent-Monégat, C.; Masson, F.; Zaidman-Rémy, A.; Heddi, A. An IMD-like pathway mediates both endosymbiont control and host immunity in the cereal weevil Sitophilus spp. Microbiome 2018, 6, 6. [Google Scholar] [CrossRef]
- Masson, F.; Rommelaere, S.; Marra, A.; Schüpfer, F.; Lemaitre, B. Dual proteomics of Drosophila melanogaster hemolymph infected with the heritable endosymbiont Spiroplasma poulsonii. PLoS ONE 2021, 16, e0250524. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Cha, W.H.; Lee, D.-W. Multiple precursor proteins of thanatin isoforms, an antimicrobial peptide associated with the gut symbiont of Riptorius pedestris. Front. Microbiol. 2022, 12, 796548. [Google Scholar] [CrossRef]
- Park, K.E.; Jang, S.H.; Lee, J.S.; Lee, A.; Kikuchi, Y.; Seo, Y.S.; Lee, B.L. The roles of antimicrobial peptide, rip-thanatin, in the midgut of Riptortus pedestris. Dev. Comp. Immunol. 2018, 78, 83–90. [Google Scholar] [CrossRef]
- Dash, R.; Bhattacharjya, S. Thanatin: An emerging host defense antimicrobial peptide with multiple modes of action. Int. J. Mol. Sci. 2021, 22, 1522. [Google Scholar] [CrossRef]
- Bourtiz, K.; Nirgianaki, A.; Onyango, P.; Savakis, C. A prokaryotic dnaA sequence in Drosophila melanogaster Wolbachia infection and cytoplasmatic incompatibility among laboratory strains. Insect Mol. Biol. 1994, 3, 131–142. [Google Scholar]
- Hamilton, P.T.; Perlman, S.J. Host defense via symbiosis in Drosophila. PLoS Pathog. 2013, 9, e1003808. [Google Scholar] [CrossRef]
- Hurst, G.D.D.; Anbutsu, H.; Kutsukake, M.; Fukatsu, T. Hidden from the host: Spiroplasma bacteria infecting Drosophila do not cause an immune response, but are suppressed by ectopic immune activation. Insect Mol. Biol. 2003, 12, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Anbutsu, H.; Moriyama, M.; Nikoh, N.; Hosokawa, T.; Futahashi, R.; Tanahashi, M.; Meng, X.Y.; Kuriwada, T.; Mori, N.; Oshima, K.; et al. Small genome symbiont underlies cuticule hardness in beetles. Proc. Natl. Acad. Sci. USA 2017, 114, E8382–E8391. [Google Scholar] [CrossRef] [PubMed]
- Herren, J.K.; Lemaitre, B. Spiroplasma and host immunity: Activation of humoral immune responses increases endosymbiont load and susceptibility to certain Gram-negative bacterial pathogens in Drosophila melanogaster. Cell. Microbiol. 2011, 13, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.A.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.; Vasan, P.T.; Raghurman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar] [CrossRef]
- Johnston, P.R.; Rolff, J. Host and symbiont jointly control gut microbiota during complete metamorphosis. PloS Pathog. 2015, 11, e1005246. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut microbiota mediate insecticide resistance in the diamondback moth, Plutella xylostella. Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef]
- Subhashini, D.V. Role of gut bacteria associated with the chlorpyrifos resistant tobacco leaf eating caterpillar Spodoptera litura on the efficacy of entomopathogenic fungi Beauvaria bassiana and Poecilomyces spp. Biol. Control. 2015, 29, 98–112. [Google Scholar] [CrossRef]
- Caragata, E.P.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Competition for amino acids between Wolbachia and the mosquito host, Aedes aegypti. Microb. Ecol. 2014, 67, 205–218. [Google Scholar] [CrossRef]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef]
- Paredes, J.C.; Herren, J.K.; Schüpfer, F.; Lamaitre, B. The role of lipid competition for endosymbiont-mediated protection against parasitoid wasp in Drosophila. mBio 2016, 7, e01006-16. [Google Scholar] [CrossRef] [PubMed]
- Pais, I.S.; Valente, R.S.; Sporniak, M.; Teixeira, L. Drosophila melanogaster establishes a species-specific mutualistic interaction with stable gut-colonizing bacteria. PLoS Biol. 2018, 16, e2005710. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Strigini, M.; Grenier, T.; Bozonnet, L.; Schwarzer, M.; Daniel, C.; Matos, R.; Leulier, F. Drosophila perpetuates nutritional mutualism by promoting the fitness of its intestinal symbiont Lactabacillus plantarum. Cell. Metab. 2018, 27, 362–377.e8. [Google Scholar] [CrossRef]
- Counce, S.J.; Poulson, D.F. The developmental effects of hereditary infections in Drosophila. Am. Zool. 1961, 1, 443. [Google Scholar]
- Weiss, B.L.; Maltz, M.; Aksoy, S. Obligate symbionts activate immune system development in the tsetse fly. J. Immunol. 2012, 188, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Weiss, B.L.; Aksoy, S. Tsetse fly microbiome: Form and function. Front. Cell. Infect. Microbiol. 2013, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, F.; Telleria, E.L.; Echodu, R.; Wu, Y.; Okedi, L.M.; Weiss, B.L.; Aksoy, S.; Caccone, A. Analysis of multiple tsetse fly populations in Uganda reveals limited diversity and species-specific gut microbiota. Appl. Environ. Microbiol. 2014, 10, 20140407. [Google Scholar] [CrossRef]
- Griffith, B.C.; Weiss, B.L.; Aksoy, E.; Mireji, P.O.; Auma, J.E.; Wamwiri, F.N.; Echodu, R.; Murilla, G.; Aksoy, S. Analysis of the gut-specific microbiome from field-captured tsetse flies, and its potential relevance to host trypanosome vector competence. BMC Microbiol. 2018, 18, 146. [Google Scholar] [CrossRef]
- Nováková, E.; Husnik, F.; Šochová, E.; Hypša, V. Arsenophonus and Sodalis symbionts in louse flies: An analogy to the Wigglesworthia and Sodalis system in tsetse flies. Appl. Environ. Microbiol. 2015, 81, 6189–6199. [Google Scholar] [CrossRef]
- Husnik, F.; Hypsa, V.; Darby, A. Insect-symbiont gene expression in the midgut bacteriocytes of blood-sucking parasite. Genome Biol. Evol. 2020, 12, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef]
- Martinson, V.G.; Moy, J.; Moran, A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef]
- Kwong, W.K.; Moran, N.A. Cultivation and characterization of the gut symbionts of honey bees and bumble bees: Description of Snodgrassella alvi gen. nov., sp. nov., a member of the family Neisseriaceae of the Betaprobacteria, and Gilliamella apicola gen. nov., sp. nov., a member of Orbaceae fam. nov., Orbales ord. nov., a sister taxon to the order ‘Enterbacteriales’ of the Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 2008–2018. [Google Scholar] [PubMed]
- Kešnerová, L.; Mortiz, R.; Engel, P. Bartonella apis sp. nov., a honey bee gut symbiont of the class Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2016, 66, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Nishida, A.; Kwong, W.K.; Koch, H.; Engel, P.; Steele, M.I.; Moran, N.A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. mBio 2016, 7, e01326-16. [Google Scholar] [CrossRef]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef]
- Skaljac, M. Bacterial symbionts of aphids (Hemiptera: Aphididae). In Biology and Ecology of Aphids; CRC Press: Boca Raton, FL, USA, 2016; pp. 100–125. [Google Scholar]
- Luna-Ramirez, K.; Skaljac, M.; Grotmann, J.; Kirfel, P.; Vilcinskas, A. Orally delivered scorpion antimicrobial peptides exhibit activity against pea aphid (Acyrthosiphon pisum) and its bacterial symbionts. Toxins 2017, 9, 261. [Google Scholar] [CrossRef]
- Luo, C.; Li, Y.; Chen, Y.; Fu, C.; Long, W.; Xiao, X.; Liao, H.; Yang, Y. Bamboo lignocellulose degradation by gut symbiotic microbiota of the bamboo snout beetle Cyrtotrachelus buqueti. Biotechnol. Biofuels 2019, 12, 70. [Google Scholar] [CrossRef]
- Huang, S.; Sheng, P.; Zhang, H. Isolation and identification of cellulolytic bacteria from the gut of Holotrichia parallela larvae (Coleoptera: Scarabaeidae). Int. J. Mol. Sci. 2012, 13, 2563–2577. [Google Scholar] [CrossRef]
- Berasategui, A.; Axelsson, K.; Nordlander, G.; Schmidt, A.; Borg-Karlson, A.K.; Gershenzon, J.; Terenius, O.; Kaltenpoth, M. The gut microbiota of the pine weevil is similar across Europe and resembles that of other conifer-feeding beetles. Mol. Ecol. 2016, 25, 4014–4031. [Google Scholar] [CrossRef] [PubMed]
- Berasategui, A.; Salem, H.; Paetz, C.; Santoro, M.; Geshenzon, J.; Kaltenpoth, M.; Schmidt, A. Gut microbiota of the pine weevil degrades conifer diterpenes and increases insect fitness. Mol. Ecol. 2017, 26, 4099–4110. [Google Scholar] [CrossRef]
- Wang, Y.; Rosen, D. Gut microbiota colonization and transmission in the burying beetle Nicrophorus vespilloides throughout development. Appl. Environ. Microbiol. 2017, 83, e03250-16. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.P.; Plata, C.; Reichelt, M.; Steiger, S.; Heckel, D.G.; Kaltenpoth, M.; Vilcinskas, A.; Vogel, H. Mirobiome-assisted carrion preservation aids larval development in a burying beetle. Proc. Natl. Sci. USA 2018, 115, 11274–11279. [Google Scholar] [CrossRef]
- Heise, P.; Liu, Y.; Degenkolb, T.; Vogel, H.; Schäberle, T.F.; Vilcinskas, A. Antibiotic-producing beneficial bacteria in gut of the burying bettle Nicrophorus vespilloides. Front. Microbiol. 2019, 10, 1178. [Google Scholar] [CrossRef]
- Ishigami, K.; Jang, S.; Itoh, H.; Kikuchi, Y. Obligate gut symbiotic association with Caballeronia in the mulberry seed bug Paradieuches dissimilis (Lygaeoidea: Rhyparochromidae). Microb. Ecol. 2022, 1–12. [Google Scholar] [CrossRef]
- Kim, J.K.; Han, S.H.; Kim, C.-H.; Jo, Y.H.; Funtahashi, R.; Kikuchi, Y.; Fukatsu, T.; Lee, B.L. Molting-associated suppression of symbiont population and up-regulation of the Riptortus-Burkholderia symbionts. Dev. Comp. Immunol. 2014, 43, 10–14. [Google Scholar] [CrossRef]
- Kim, J.K.; Lee, J.B.; Jang, H.A.; Han, Y.S.; Fukatsu, T. Understanding regulation of the host-mediated gut symbiont population and the symbiont-mediated host immunity in the Riptortus-Burkholderia symbiosis system. Dev. Comp. Immunol. 2016, 64, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Jordan, M.S.; Adams, S.M.; Suen, G.; Goodwin, L.A.; Davenport, K.W.; Currie, C.R.; Raffa, K.F. Cellulose-degrading bacteria associated with the invasive woodwasp Sirex noctilio. ISME J. 2011, 5, 1323–1331. [Google Scholar] [CrossRef]
- Heddi, A.; Charles, H.; Khatchadourin, C.; Bonnot, G.; Nardon, P. Molecular characterization of the principal symbiotic bacteria of the weevil Sitophilus oryze: A peculiar G + C content of an endocytobiotic DNA. J. Mol. Evol. 1998, 47, 52–61. [Google Scholar] [CrossRef]
- Oakeson, K.F.; Gil, R.; Clayton, A.L.; Dunn, D.M.; von Niederhausern, A.C.; Hamil, C.; Aoyagi, A.; Duval, B.; Baca, A.; Silva, F.J.; et al. Genome degeneration and adaptation in a nascent stage of symbiosis. Genome Biol. Evol. 2014, 6, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Zuber, L.; Dominguez-Santos, R.; Garcia-Ferris, C.; Silva, F.J. Identification of the gene repetitorie of the IMD pathway and expression of antimicrobial peptides genes in several tissues and hemolymph of the cocroach Blattella germanica. Int. J. Mol. Sci. 2022, 23, 8444. [Google Scholar] [CrossRef]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. Biol. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Simhadri, R.K.; Fast, F.M.; Guo, R.; Schultz, M.J.; Vaisman, N.; Ortiz, I.; Bybee, J.; Slatko, B.E.; Frydman, H.M. The gut commensal microbiome of Drosophila melanogaster is modified by the endosymbiont Wolbachia. mSphere 2017, 2, e00287-17. [Google Scholar] [CrossRef]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, 2753–2763. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Vasanthakrishnan, R.B.; Siva-Jothy, J.; Monteih, K.M.; Brown, S.P.; Vale, P.F. The route of infection determines Wolbachia antibacterial protection in Drosophila. Proc. Biol. Sci. 2017, 284, 20170809. [Google Scholar] [CrossRef]
- Wong, Z.S.; Hedges, L.M.; Brownlie, J.C.; Johnson, K.N. Wolbachia-mediated antibacterial protection and immune gene regulation in Drosophila. PLoS ONE 2011, 6, e25430. [Google Scholar] [CrossRef] [PubMed]
- Rottschaefer, S.M.; Lazzaro, B.P. No effect of Walbacia on resistance to intracellular infection by pathogenic bacteria in Drosophila melanogaster. PLoS ONE 2012, 7, e40500. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.H.; Cohen, L.B.; Wasserman, S.A. Effector specificity and function in Drosophila innate immunity: Getting AMPed and dropping Boms. PLoS Pathog. 2020, 16, e1008480. [Google Scholar] [CrossRef]
- Clemmons, A.W.; Lindsay, S.A.; Wasserman, S.A. An effector Peptide family required for Drosophila toll-mediated immunity. PLoS Pathog. 2015, 11, e1004876. [Google Scholar] [CrossRef]
- Lindsay, S.A.; Lin, S.J.H.; Wasserman, S.A. Short-form bomanins mediate humoral immunity in Drosophila. J. Innate Immun. 2018, 10, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Lou, Y.; Tidu, A.; Bulet, P.; Heinekamp, T.; Martin, F.; Brakhage, A.; Li, Z.; Liégeois, S.; Ferrandon, D. The Toll pathway mediates Drosophila resilience to Aspergillus mycotoxins through specific bomanins. EMBO Rep. 2022, 24, e56036. [Google Scholar] [CrossRef]
- Hanson, M.A.; Cohen, L.B.; Marra, A.; Iatsenko, I.; Wasserman, S.A.; Lemaitre, B. The Drosophila baramicin polypeptide gene protects against fungal infection. PLoS Pathog. 2021, 17, e1009846. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lou, Y.; Liu, J.; Bulet, P.; Jiao, R.; Hoffmann, J.A.; Liégeois, S.; Li, Z.; Ferrandon, D. A Toll pathway effector protects Drosophila specifically from distinct toxins secreted by a fungus or a bacterium. bioRxiv 2022. [Google Scholar] [CrossRef]
- Cohen, L.B.; Lindsay, S.A.; Xu, Y.; Lin, S.J.H.; Wasserman, S.A. The daisho peptides mediate Drosophila defense against a subset of filamentous fungi. Front. Immunol. 2020, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- West, C.; Silverman, N. p38b and JAK-STAT signaling protect against Invertebrate iridescent virus 6 infection in Drosophila. PLoS Pathog. 2018, 14, e1007020. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Imler, J.L. Sensing and signalling viral infection in Drosophila. Dev. Comp. Immunol. 2021, 117, 103985. [Google Scholar] [CrossRef]
- Guo, Z.; Li, Y.; Ding, S.W. Small RNA-based antimicrobial immunity. Nat. Rev. Immunol. 2019, 19, 31–44. [Google Scholar] [CrossRef]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasites Vectors 2019, 12, 165. [Google Scholar] [CrossRef]
- McMenamin, A.J.; Daughenbaugh, K.F.; Parekh, F.; Pizzorno, M.C.; Flenniken, M.L. Honey bee and bumble bee antiviral defense. Viruses 2018, 10, 395. [Google Scholar] [CrossRef]
- Mondotte, J.A.; Saleh, M.C. Antiviral immune response and the route of infection in Drosophila melanogaster. Adv. Virus Res. 2018, 100, 247–278. [Google Scholar] [PubMed]
- Huang, Z.; Kingsolver, M.B.; Avadhanula, V.; Hardy, R.W. An antiviral role for antimicrobial peptides during the arthropod response to alphavirus replication. J. Virol. 2013, 87, 4272–4280. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.R.; van der Most, T.; Miesen, P. Genetic determinants of antiviral immunity in dipteran insects—Compiling the experimental evidence. Dev. Comp. Immunol. 2021, 119, 104010. [Google Scholar] [CrossRef]
- Luplertlop, N.; Surasombatpattana, P.; Patramool, S.; Dumas, E.; Wasinpiyamongkol, L.; Saune, L.; Hamel, R.; Bernard, E.; Sereno, D.; Thomas, F. Induction of a peptide with activity against a broad spectrum of pathogens in the Aedes aegypti salivary gland, following infection with dengue virus. PLoS Pathog. 2011, 7, e1001252. [Google Scholar] [CrossRef]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Dimopoulos, G. The Toll immune signaling pathway control conserved anti-dengue defenses across diverse Ae. aegypti strains and against multiple dengue virus serotypes. Dev. Comp. Immunol. 2010, 34, 625–629. [Google Scholar] [CrossRef]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef]
- Xiao, X.; Liu, Y.; Zhang, X.; Wang, J.; Li, Z.; Pang, X.; Wang, P.; Cheng, G. Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides. PLoS Pathog. 2014, 10, e1004027. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-dorsal pathway is required for resistance to viral oral infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [PubMed]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, L.; Jouanguy, E.; Dostert, C.; Zachary, D.; Dimarcq, J.L.; Bulet, P.; Imler, J.L. Pherokine-2 and-3: Two Drosophila molecules related to pheromone/odor-binding proteins induced by viral and bacterial infections. Eur. J. Biochem. 2003, 270, 3398–3407. [Google Scholar] [CrossRef]
- Avadhanula, V.; Weasner, B.P.; Hardy, G.G.; Kumar, J.P.; Hardy, R.W. A novel system for the launch of alphavirus RNA synthesis reveals a role for the Imd pathway in arthropod antiviral response. PLoS Pathog. 2009, 5, e1000582. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Brown, S.P.; Annoscia, D.; Del Piccolo, F.; Di Prisco, G.; Varricchio, P.; Vedova, G.D.; Cattonaro, F.; Caprio, E.; Pennacchio, F. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 2012, 8, e1002735. [Google Scholar] [CrossRef]
- Palmer, W.H.; Joosten, J.; Overheul, G.J.; Jansen, P.W.; Vermeulen, M.; Obbard, D.J.; Van Rij, R.P. Induction and suppression of NF-κB signalling by a DNA virus of Drosophila. J. Virol. 2019, 93, e01443-18. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, W.; Gao, X.; Li, W.; Qi, S.; Guo, D.; Ajayi, O.E.; Ding, S.W.; Wu, Q. IncRNA sensing of a viral suppressor of RNAi activates non-canonical innate immune signaling in Drosophila. Cell Host Microbe 2020, 27, 115–128. [Google Scholar] [CrossRef]
- Lü, P.; Pan, Y.; Yang, Y.; Zhu, F.; Li, C.; Guo, Z.; Yao, Q.; Chen, K. Discovery of anti-viral molecules and their vital functions in Bombyx mori. J. Invertebr. Pathol. 2018, 154, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.Y.; Tang, X.D.; Lv, Z.Y.; Wang, X.Y.; Tian, C.H.; Xu, Y.P.; Zhang, C.X. Gene expression profiling of resistant and susceptible Bombyx mori strains reveals nucleopolyhedrovirus-associated variations in host gene transcript levels. Genomics 2009, 94, 138–145. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; He, H.J.; Zhao, X.F.; Wang, J.X. Immune responses of Helicoverpa armigera to different kinds of pathogens. BMC Immunol. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Moreno-Habel, D.A.; Biglang-awa, I.M.; Dulce, A.; Luu, D.D.; Garcia, P.; Weers, P.M.; Haas-Stapleton, E.J. Inactivation of the budded virus of Autographa californica M nucleopolyhedrovirus by gloverin. J. Invertebr. Pathol. 2012, 110, 92–101. [Google Scholar] [CrossRef]
- Traiyasut, P.; Mookhploy, W.; Kimura, K.; Yoshiyama, M.; Khongphinitbunjong, K.; Chantawannakul, P.; Chukeatirote, E. First detection of honey bee viruses in wax moth. Chiang Mai J. Sci. 2016, 43, 695–698. [Google Scholar]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial peptides as potential antiviral factors in insect antiviral immune response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Wang, H.; Smagghe, G.; Meeus, I. The Single von Willebrand factor C-domain protein (SVC) coding gene is not involved in the hymenoptaecin upregulation after Israeli acute paralysis virus (IAPV) injection in the bumblebee Bombus terrestris. Dev. Comp. Immunol. 2018, 81, 152–155. [Google Scholar] [CrossRef]
- Quintana, S.; Brasesco, C.; Negri, P.; Marin, M.; Pagnuco, I.; Szawarski, N.; Reynaldi, F.; Larsen, A.; Eguaras, M.; Maggi, M. Up-regulated pathways in response to deformed wing virus infection in Apis mellifera (Hymenoptera: Apidae). Rev. De La Soc. Entomológica Argent. 2019, 78, 1–11. [Google Scholar] [CrossRef]
- Phokasem, P.; Mookhploy, W.; Krongdang, S.; Sinpoo, C.; Chantawannakul, P. Interaction between thiamethoxam and deformed wing virus type A on wing characteristics and expression of immune and apoptosis genes in Apis mellifera. Insects 2022, 13, 515. [Google Scholar] [CrossRef] [PubMed]
- Mookhploy, W.; Krongdang, S.; Chantawannakul, P. Effects of deformed wing virus infection on expressions of immune-and apoptosis-related genes in western honeybees (Apis mellifera). Insects 2021, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- Moran, D.; Dutta, U.; Kunnumakkara, A.B.; Daimari, E.; Deka, B. Insect venoms and their bioactive components: A novel therapeutic approach in chronic diseases and cancer. J. Cancer Sci. Clin. Ther. 2022, 6, 360–382. [Google Scholar] [CrossRef]
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef]
- dos Santos-Pinto, J.R.A.; Perez-Riverol, A.; Lasa, A.M.; Palma, M.S. Diversity of peptidic and proteinaceous toxins from social Hymenoptera venoms. Toxicon 2018, 148, 172–196. [Google Scholar] [CrossRef]
- Orivel, J.; Redeker, V.; Le Caer, J.P.; Krier, F.; Revol-Junelles, A.M.; Longeon, A.; Chaffotte, A.; Dejean, A.; Rossier, J. Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii. J. Biol. Chem. 2001, 276, 17823–17829. [Google Scholar] [CrossRef]
- He, S.; Stone, T.A.; Deber, C.M. Uncoupling amphipathicity and hydrophobicity: Role of charge clustering in membrane interactions of cationic antimicrobial peptides. Biochemistry 2021, 60, 2586–2592. [Google Scholar] [CrossRef]
- Schifano, N.P.; Caputo, G.A. Investigation of the role of hydrophobic amino acids on the structure-activity relationship in the antimicrobial venom peptide ponericin L1. J. Membr. Biol. 2022, 255, 537–551. [Google Scholar] [CrossRef]
- Wanandy, T.; Gueven, N.; Davies, N.W.; Brown, S.G.; Wiese, M.D. Pilosulins: A review of the structure and mode of action of venom peptides from an Australian ant Myrmecia pilosula. Toxicon 2015, 98, 54–61. [Google Scholar] [CrossRef]
- Yacoub, T.; Rima, M.; Karam, M.; Sabatier, J.M.; Fajloun, Z. Antimicrobials from venomous animals: An overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- Rádis-Baptista, G.; Dodou, H.V.; Prieto-da-Silva, Á.R.B.; Zaharenko, A.J.; Kazuma, K.; Nihei, K.I.; Inagaki, H.; Mori-Yasumoto, K.; Konno, K. Comprehensive analysis of peptides and low molecular weight components of the giant ant Dinoponera quadriceps venom. Biol. Chem. 2020, 401, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.F.; Huang, C.; Chong, C.M.; Leung, S.W.; Prieto-da-Silva, A.R.; Havt, A.; Quinet, Y.P.; Martins, A.M.; Lee, S.M.; Radis-Baptista, G. Transcriptome analysis in venom gland of the predatory giant ant Dinoponera quadriceps: Insights into the polypeptide toxin arsenal of hymenopterans. PLoS ONE 2014, 9, e87556. [Google Scholar] [CrossRef] [PubMed]
- Dodou Lima, H.V.; de Paula Cavalcante, C.S.; Rádis-Baptista, G. Antifungal in vitro activity of pilosulin- and ponericin-like peptides from the giant ant Dinoponera quadriceps and synergistic effects with antimycotic drugs. Antibiotics 2020, 9, 354. [Google Scholar] [CrossRef]
- Huang, S.; Jianhua, W.A.N.G.; Xiaozhong, W.A.N.G.; Chenghong, L.I. Melittin: A key composition of honey bee venom with diverse pharmaceutical function. In Proceedings of the International Conference on Biological Engineering and Pharmacy 2016 (BEP 2016), Shanghai, China, 9–11 December 2016; Atlantis Press: Dordrecht, The Netherlands, 2016; pp. 193–197. [Google Scholar]
- Wang, A.; Zheng, Y.; Zhu, W.; Yang, L.; Yang, Y.; Peng, J. Melittin-based nano-delivery systems for cancer therapy. Biomolecules 2022, 12, 118. [Google Scholar] [CrossRef]
- Wehbe, R.; Frangieh, J.; Rima, M.; El Obeid, D.; Sabatier, J.M.; Fajloun, Z. Bee Venom: Overview of main compounds and bioactivities for therapeutic interests. Molecules 2019, 24, 2997. [Google Scholar] [CrossRef] [PubMed]
- Carpena, M.; Nuñez-Estevez, B.; Soria-Lopez, A.; Simal-Gandara, J. Bee Venom: An updating review of its bioactive molecules and its health applications. Nutrients 2020, 12, 3360. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Han, S.M.; Park, K.-K. Therapeutic effects of apamin as a bee venom component for non-neoplastic disease. Toxins 2020, 12, 195. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Jin, B.R. Secapin, a bee venom peptide, exhibits anti-fibrinolytic, anti-elastolytic, and anti-microbial activities. Dev. Comp. Immunol. 2016, 63, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Doublet, V.; Poeschl, Y.; Gogol-Döring, A.; Alaux, C.; Annoscia, D.; Aurori, C.; Barribeau, S.M.; Bedoya-Reina, O.C.; Brown, M.J.; Bull, J.C.; et al. Unity in defence: Honeybee workers exhibit conserved molecular responses to diverse pathogens. BMC Genom. 2017, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.J.; Park, E.; Asandei, A.; Choi, J.Y.; Lee, S.C.; Seo, C.H.; Luchian, T.; Park, Y. Bee venom-derived antimicrobial peptide melectin has broad-spectrum potency, cell selectivity, and salt-resistant properties. Sci. Rep. 2020, 10, 10145. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.; Kaminska, D.; Matuszewska, E.; Hołderna-Kedzia, E.; Rogacki, J.; Matysiak, J. Promising antimicrobial properties of bioactive compounds from different honeybee products. Molecules 2021, 26, 4007. [Google Scholar] [CrossRef]
- Shi, N.; Szanto, T.G.; He, J.; Schroeder, C.I.; Walker, A.A.; Deuis, J.R.; Vetter, I.; Panyi, G.; King, G.F.; Robinson, S.D. Venom composition and pain-causing toxins of the Australian great carpenter bee Xylocopa aruana. Sci. Rep. 2022, 12, 22168. [Google Scholar] [CrossRef]
- Roberson, M.G.; Smith, D.K.; White, S.M.; Wallace, I.S.; Tucker, M.J. Interspecies bombolitins exhibit structural diversity upon membrane binding, leading to cell specificity. Biophys. J. 2019, 116, 1064–1074. [Google Scholar] [CrossRef]
- Rajesh, R.P.; Arun, R.; Selvam, M.M.; Alphonse, C.R.W.; Rajasekar, M.; Franklin, J.B. Identification and characterisation of novel wasp mastoparans and chemotactic peptides from the venom of social wasp Polistes stigma (Hymenoptera: Vespidae: Polistinae). J. Venom Res. 2021, 11, 16–20. [Google Scholar]
- Aschi, M.; Perini, N.; Bouchemal, N.; Luzi, C.; Savarin, P.; Migliore, L.; Bozzi, A.; Sette, M. Structural characterization and biological activity of Crabrolin peptide isoforms with different positive charge. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183055. [Google Scholar] [CrossRef]
- Cantini, F.; Luzi, C.; Bouchemal, N.; Savarin, P.; Bozzi, A.; Sette, M. Effect of positive charges in the structural interaction of crabrolin isoforms with lipopolysaccharide. J. Pept. Sci. 2020, 26, e3271. [Google Scholar] [CrossRef]
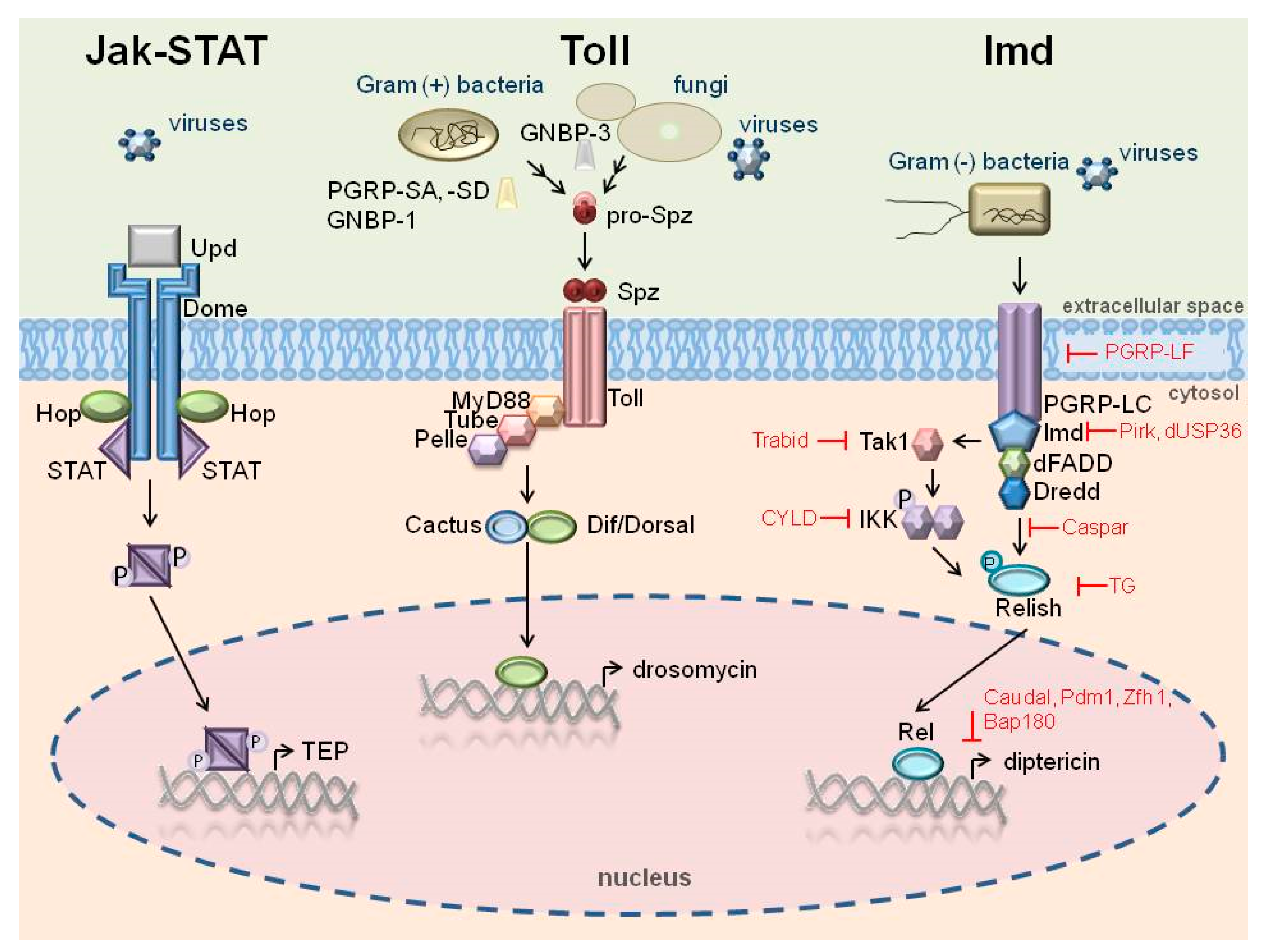
 . The Imd pathway can also be activated by viruses.
. The Imd pathway can also be activated by viruses.
. The Imd pathway can also be activated by viruses.
. The Imd pathway can also be activated by viruses.

| G. mellonella AMP | Accession Number/ Peptide ID | Predicted Antiviral Activity | AVP Value | Non-AVP Value |
|---|---|---|---|---|
| gallerimycin | XP_026765221.1 | + | 1 | 0 |
| lebocin-like anionic peptide 1 | P85211.1 | + | 0.98 | 0.02 |
| cecropin D-like peptide | P85210.1 | + | 0.93 | 0.07 |
| G. mellonella cecropin | XP_026754247.1 | + | 0.746 | 0.254 |
| apolipophoricin | P80703 | + | 0.698 | 0.302 |
| proline rich peptide 2 | P85212.1 | − | 0.324 | 0.676 |
| defensin-like peptide | P85215 | − | 0.272 | 0.728 |
| defensin | P85213 | − | 0.268 | 0.732 |
| proline rich peptide 1 | P85214.1 | − | 0.004 | 0.996 |
| anionic peptide 2 | P85016 | − | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the Role of Antimicrobial Peptides in Insects. Int. J. Mol. Sci. 2023, 24, 5753. https://doi.org/10.3390/ijms24065753
Stączek S, Cytryńska M, Zdybicka-Barabas A. Unraveling the Role of Antimicrobial Peptides in Insects. International Journal of Molecular Sciences. 2023; 24(6):5753. https://doi.org/10.3390/ijms24065753
Chicago/Turabian StyleStączek, Sylwia, Małgorzata Cytryńska, and Agnieszka Zdybicka-Barabas. 2023. "Unraveling the Role of Antimicrobial Peptides in Insects" International Journal of Molecular Sciences 24, no. 6: 5753. https://doi.org/10.3390/ijms24065753
APA StyleStączek, S., Cytryńska, M., & Zdybicka-Barabas, A. (2023). Unraveling the Role of Antimicrobial Peptides in Insects. International Journal of Molecular Sciences, 24(6), 5753. https://doi.org/10.3390/ijms24065753