
Semi-Targeted Profiling of the Lipidome Changes Induced by Erysiphe Necator in Disease-Resistant and Vitis vinifera L. Varieties
 , , , , , and
, , , , , and
Abstract
1. Introduction
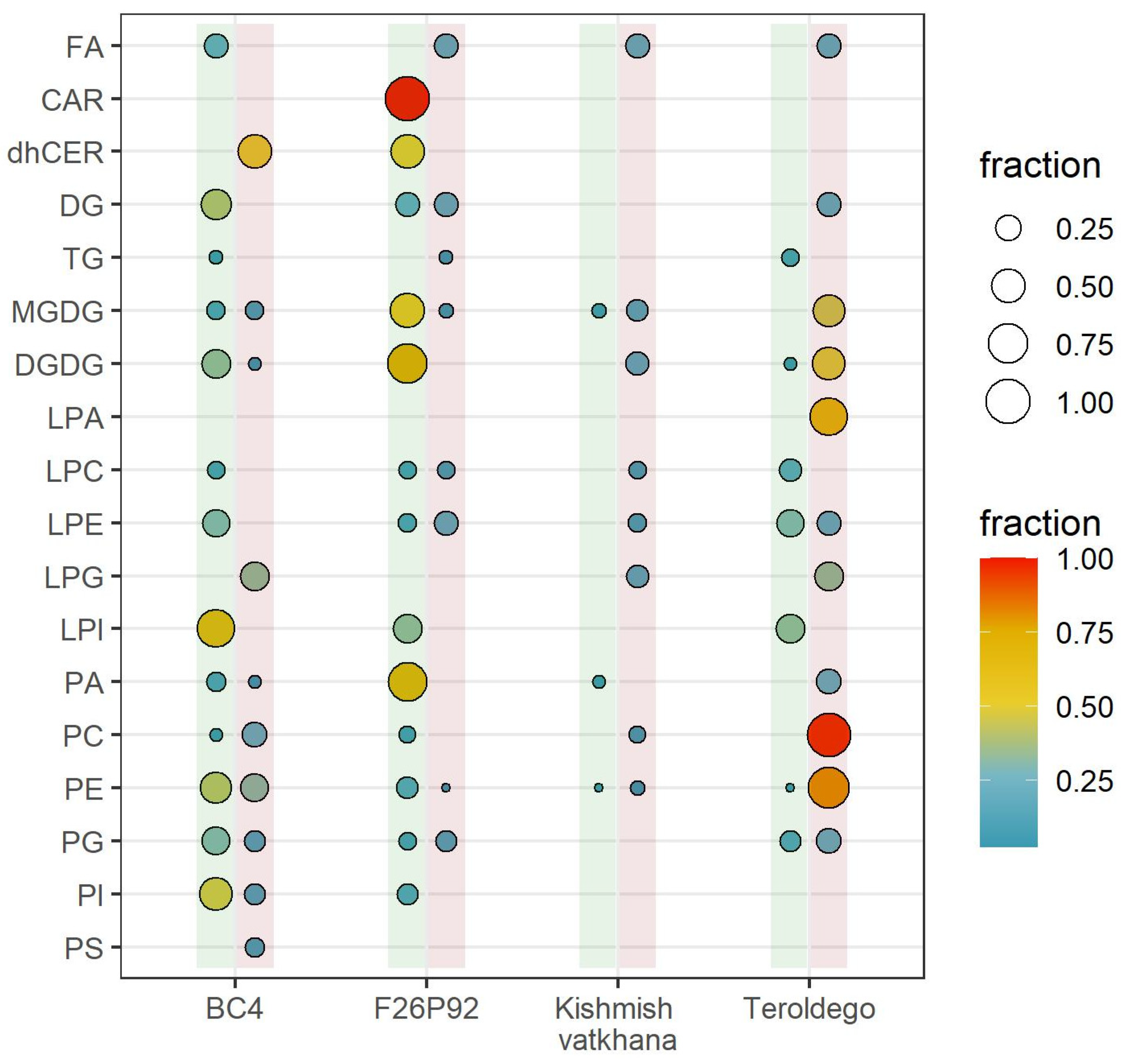
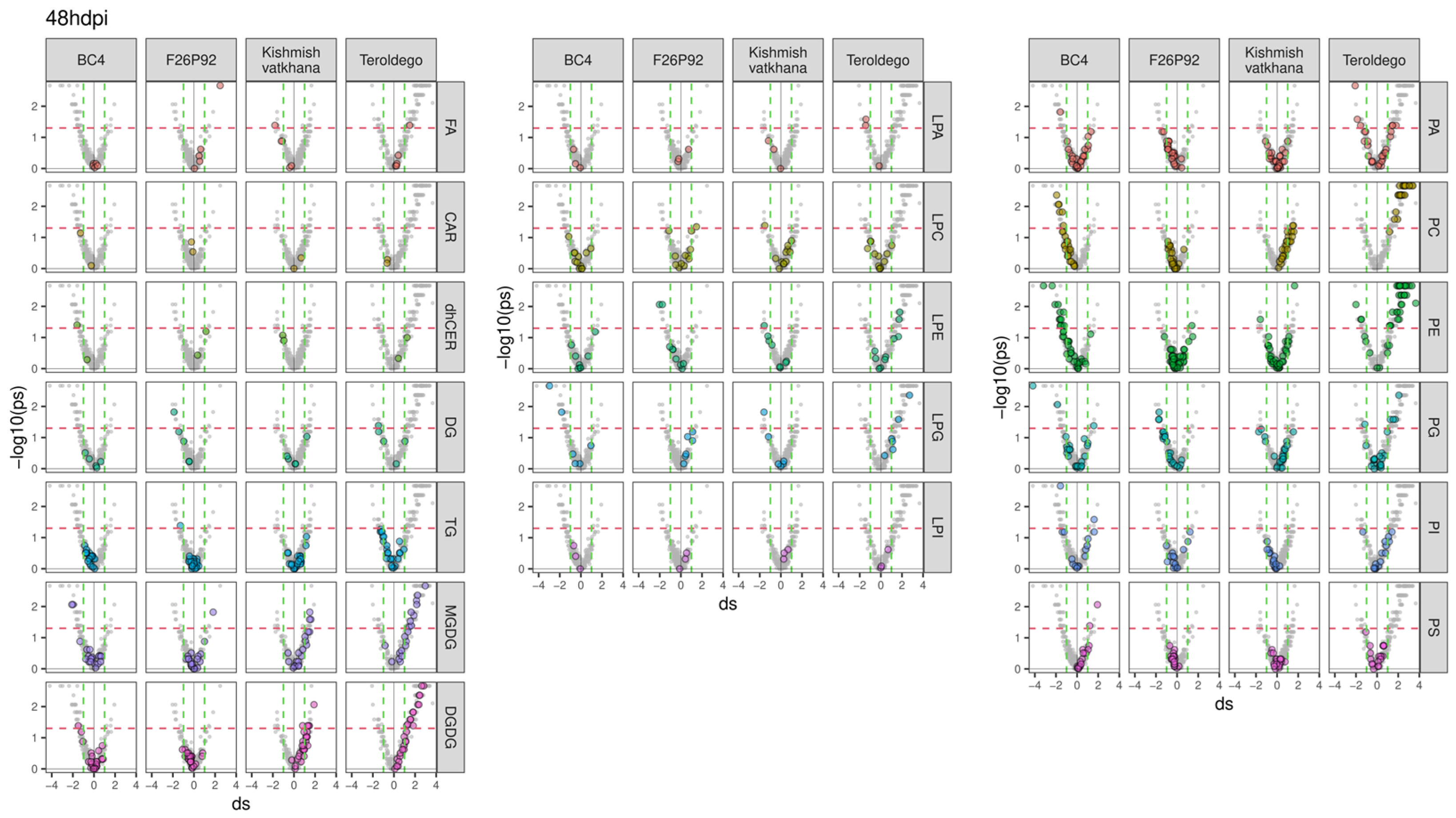
2. Results
2.1. Phenotypic Resistance
2.2. Lipid Modulation of the Grapevine–E. necator Interaction during the First Hours of Infection
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design and Artificial Inoculation
4.3. Disease Assessment
4.4. Lipid Extraction and Analysis
4.5. Data Processing
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CAR | carnitine |
| CER | ceramide |
| DBs | double bonds |
| DG | diacylglycerol |
| DGDG | digalactosyldiacylglycerol |
| dhCER | dihydroceramide |
| ETI | effector-triggered immunity |
| FA | free fatty acid |
| GL | glycerolipid |
| glcCER | glucosyl ceramide |
| glc-dhCER | glucosyldihydroceramide |
| GP | glycerophospholipid |
| HPLC | high-performance liquid chromatography |
| IS | internal standard |
| KMD | Kendrick mass defect |
| LC | liquid chromatography |
| LPA | lyso-glycerophosphate |
| LPC | lyso-glycerophosphocholine |
| LPE | lyso-glycerophosphoethanolamine |
| LPI | lyso-glycerophosphoinositol |
| LPG | lyso-glycerophosphoglycerol |
| MG | monoacylglycerol |
| MGDG | monogalactosyldiacylglycerol |
| MS | mass spectrometry |
| MW | molecular weight |
| nCs | number of carbons |
| OIV | Organisation Internationale de la Vigne et du Vin |
| PA | glycerophosphate |
| PAMPs | pathogen-associated molecular patterns |
| PC | glycerophosphocholine |
| PCA | principal component analysis |
| PCD | programmed cell death |
| PE | glycerophosphoethanolamine |
| PI | glycerophosphoinositol |
| PG | glycerophosphoglycerol |
| PK | polyketide |
| PLD | phospholipase D |
| PR | prenol lipid |
| PS | glycerophosphoserine |
| PTI | pathogen-triggered immunity |
| QTLs | quantitative trait loci |
| QC | quantitative control |
| REN | resistance to Erysiphe necator |
| RUN | resistance to Uncinula necator |
| ROSs | reactive oxygen species |
| RT | retention time |
| SAR | systemic acquired resistance |
| SL | saccharolipid |
| SM | sphingomyelin |
| SP | sphingolipid |
| ST | sterol |
| STD | standard |
| TG | triacylglycerol |
| UPLC | Ultra-high performance liquid chromatography |
References
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification, structures and tools. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef]
- Blanksby, S.J.; Mitchell, T.W. Advances in mass spectrometry for lipidomics. Annu. Rev. Anal. Chem. 2010, 3, 433–465. [Google Scholar] [CrossRef]
- Masuero, D.; Škrab, D.; Chitarrini, G.; Garcia-Aloy, M.; Franceschi, P.; Sivilotti, P.; Guella, G.; Vrhovsek, U. Grape Lipidomics: An Extensive Profiling thorough UHPLC-MS/MS Method. Metabolites 2021, 11, 827. [Google Scholar] [CrossRef]
- Furt, F.; Simon-Plas, F.; Mongrand, S. The Plant Plasma Membrane; Murphy, A.S., Schulz, B., Peer, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–30. [Google Scholar] [CrossRef]
- Cagliari, A.; Margis, R.; dos Santos Maraschin, F.; Turchetto-Zolet, A.C.; Loss, G.; Margis-Pinheiro, M. Biosynthesis of Triacylglycerols (TAGs) in Plants and algae. Int. J. Plant Biol. 2011, 2, e10. [Google Scholar] [CrossRef]
- van Leeuwen, W.; Okrész, L.; Bögre, L.; Munnik, T. Learning the lipid language of plant signalling. Trends Plant Sci. 2004, 9, 378–384. [Google Scholar] [CrossRef]
- Okazaki, Y.; Saito, K. Roles of lipids as signaling molecules and mitigators during stress response in plants. Plant J. Cell Mol. Biol. 2014, 79, 584–596. [Google Scholar] [CrossRef]
- Siebers, M.; Brands, M.; Wewer, V.; Duan, Y.; Hölzl, G.; Dörmann, P. Lipids in plant-microbe interactions. Biochim. Biophys. Acta 2016, 1861, 1379–1395. [Google Scholar] [CrossRef]
- Cavaco, A.R.; Matos, A.R.; Figueiredo, A. Speaking the language of lipids: The cross-talk between plants and pathogens in defence and disease. Cell. Mol. Life Sci. 2021, 78, 4399–4415. [Google Scholar] [CrossRef]
- Huang, P.-Y.; Zimmerli, L. Enhancing crop innate immunity: New promising trends. Front. Plant Sci. 2014, 5, 624. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant–pathogen effectors: Cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B. The black-box of plant apoplast lipidomes. Front. Plant Sci. 2016, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Regente, M.; Monzón, G.C.; de la Canal, L. Phospholipids are present in extracellular fluids of imbibing sunflower seeds and are modulated by hormonal treatments. J. Exp. Bot. 2008, 59, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Ludovici, M.; Ialongo, C.; Reverberi, M.; Beccaccioli, M.; Scarpari, M.; Scala, V. Quantitative profiling of oxylipins through comprehensive LC-MS/MS analysis of Fusarium verticillioides and maize kernels. Food Addit. Contam. Part A 2014, 31, 2026–2033. [Google Scholar] [CrossRef]
- Laureano, G.; Figueiredo, J.; Cavaco, A.R.; Duarte, B.; Caçador, I.; Malhó, R.; Sousa Silva, M.; Matos, A.R.; Figueiredo, A. The interplay between membrane lipids and phospholipase A family members in grapevine resistance against Plasmopara viticola. Sci. Rep. 2018, 8, 14538. [Google Scholar] [CrossRef]
- Müller, V.; Amé, M.V.; Carrari, V.; Gieco, J.; Asis, R. Lipoxygenase Activation in Peanut Seed Cultivars Resistant and Susceptible to Aspergillus parasiticus Colonization. Phytopathology 2014, 104, 1340–1348. [Google Scholar] [CrossRef]
- Suh, J.H.; Niu, Y.S.; Wang, Z.; Gmitter, F.G., Jr.; Wang, Y. Metabolic analysis reveals altered long-chain fatty acid metabolism in the host by huanglongbing disease. J. Agric. Food Chem. 2018, 66, 1296–1304. [Google Scholar] [CrossRef]
- Gadoury, D.M.; Cadle-Davidson, L.; Wilcox, W.F.; Dry, I.B.; Seem, R.C.; Milgroom, M.G. Grapevine powdery mildew (Erysiphe necator): A fascinating system for the study of the biology, ecology and epidemiology of an obligate biotroph. Mol. Plant Pathol. 2012, 13, 1–16. [Google Scholar] [CrossRef]
- Agurto, M.; Schlechter, R.O.; Armijo, G.; Solano, E.; Serrano, C.; Contreras, R.A.; Zúñiga, G.E.; Arce-Johnson, P. RUN1 and REN1 Pyramiding in Grapevine (Vitis vinifera cv. Crimson Seedless) Displays an Improved Defense Response Leading to Enhanced Resistance to Powdery Mildew (Erysiphe necator). Front. Plant Sci. 2017, 8, 758. [Google Scholar] [CrossRef]
- McDonald, B.A.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef]
- Merdinoglu, D.; Schneider, C.; Prado, E.; Wiedemann-Merdinoglu, S.; Mestre, P. Breeding for durable resistance to downy and powdery mildew in grapevine. OENO One 2018, 52, 189–195. [Google Scholar] [CrossRef]
- Ciubotaru, R.M.; Franceschi, P.; Vezzulli, S.; Zulini, L.; Stefanini, M.; Oberhuber, M.; Robatscher, P.; Chitarrini, G.; Vrhovsek, U. Secondary and primary metabolites reveal putative resistance-associated biomarkers against Erysiphe necator in resistant grapevine genotypes. Front. Plant Sci. 2023, 14, 1112157. [Google Scholar] [CrossRef]
- Negrel, L.; Halter, D.; Wiedemann-Merdinoglu, S.; Rustenholz, C.; Merdinoglu, D.; Hugueney, P.; Baltenweck, R. Identification of Lipid Markers of Plasmopara viticola Infection in Grapevine Using a Non-targeted Metabolomic Approach. Front. Plant Sci. 2018, 9, 360. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, A.R.; Maia, M.; Laureano, G.; Duarte, B.; Caçador, I.; Sousa Silva, M.; Matos, A.R.; Figueiredo, A. P-285-Identifying grapevine lipid biomarkers of resistance/susceptibility to Plasmopara viticola; towards a sustainable viticulture. Free Radic. Biol. Med. 2018, 120, 131. [Google Scholar] [CrossRef]
- Della Corte, A.; Chitarrini, G.; Di Gangi, I.M.; Masuero, D.; Soini, E.; Mattivi, F.; Vrhovsek, U. A rapid LC-MS/MS method for quantitative profiling of fatty acids, sterols, glycerolipids, glycerophospholipids and sphingolipids in grapes. Talanta 2015, 140, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Chitarrini, G.; Soini, E.; Riccadonna, S.; Franceschi, P.; Zulini, L.; Masuero, D.; Vecchione, A.; Stefanini, M.; Di Gaspero, G.; Mattivi, F.; et al. Identification of biomarkers for defense response to Plasmopara viticola in a resistant grape variety. Front. Plant Sci. 2017, 8, 1524. [Google Scholar] [CrossRef]
- Laureano, G.; Cavaco, A.R.; Matos, A.R.; Figueiredo, A. Fatty Acid Desaturases: Uncovering Their Involvement in Grapevine Defence against Downy Mildew. Int. J. Mol. Sci. 2021, 22, 5473. [Google Scholar] [CrossRef]
- Viret, O.; Spring, J.L.; Gindro, K. Stilbenes: Biomarkers of grapevine resistance to fungal diseases. OENO One 2018, 52, 235–240. [Google Scholar] [CrossRef]
- Boddy, L. Pathogens of autotrophs. In The Fungi; Watkinson, S.C., Money, N.P., Boddy, L., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 245–292. [Google Scholar]
- Ciubotaru, R.M.; Franceschi, P.; Zulini, L.; Stefanini, M.; Škrab, D.; Rossarolla, M.D.; Robatscher, P.; Oberhuber, M.; Vrhovsek, U.; Chitarrini, G. Mono-Locus and Pyramided Resistant Grapevine Cultivars Reveal Early Putative Biomarkers Upon Artificial Inoculation With Plasmopara viticola. Front. Plant Sci. 2021, 12, 693887. [Google Scholar] [CrossRef]
- Feechan, A.; Kocsis, M.; Riaz, S.; Zhang, W.; Gadoury, D.M.; Walker, M.A.; Dry, I.B.; Reisch, B.; Cadle-Davidson, L. Strategies for RUN1 deployment using RUN2 and REN2 to manage grapevine powdery mildew informed by studies of race specificity. Phytopathology 2015, 105, 1104–1113. [Google Scholar] [CrossRef]
- Pap, D.; Riaz, S.; Dry, I.B.; Jermakow, A.; Tenscher, A.C.; Cantu, D.; Oláh, R.; Walker, M.A. Identification of two novel powdery mildew resistance loci, Ren6 and Ren7, from the wild Chinese grape species Vitis piasezkii. BMC Plant Biol. 2016, 16, 170. [Google Scholar] [CrossRef] [PubMed]
- Zendler, D.; Töpfer, R.; Zyprian, E. Confirmation and fine mapping of the resistance locus Ren9 from the grapevine cultivar ‘Regent’. Plants 2020, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Zuniga, V.; Vidal Valenzuela, Á.; Barba, P.; Espinoza Cancino, C.; Romero-Romero, J.L.; Arce-Johnson, P. Powdery Mildew Resistance Genes in Vines: An Opportunity to Achieve a More Sustainable Viticulture. Pathogens 2022, 11, 703. [Google Scholar] [CrossRef]
- Julius Kühn-Institut-Federal Research Centre for Cultivated Plants (JKI), Institute for Grapevine Breeding-Geilweilerhof (ZR). Available online: www.vivc.de/loci (accessed on 22 October 2022).
- Feechan, A.; Anderson, C.; Torregrosa, L.; Jermakow, A.; Mestre, P.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; Walker, A.R.; Cadle-Davidson, L.; Reisch, B.; et al. Genetic dissection of a TIR-NB-LRR locus from the wild North American grapevine species Muscadinia rotundifolia identifies paralogous genes conferring resistance to major fungal and oomycete pathogens in cultivated grapevine. Plant J. 2013, 76, 661–674. [Google Scholar] [CrossRef]
- Welter, L.J.; Göktürk-Baydar, N.; Akkurt, M.; Maul, E.; Eibach, R.; Töpfer, R.; Zyprian, E.M. Genetic mapping and localization of quantitative trait loci affecting fungal disease resistance and leaf morphology in grapevine (Vitis vinifera L). Mol. Breed. 2007, 20, 359–374. [Google Scholar] [CrossRef]
- Hoffmann, S.; Di Gaspero, G.; Kovács, L.; Howard, S.; Kiss, E.; Galbács, Z.; Testolin, R.; Kozma, P. Resistance to Erysiphe necator in the grapevine ‘Kishmish Vatkana’ is controlled by a single locus through restriction of hyphal growth. Theor. Appl. Genet. 2008, 116, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Zendler, D.; Schneider, P.; Töpfer, R.; Zyprian, E. Fine mapping of Ren3 reveals two loci mediating hypersensitive response against Erysiphe Necator in grapevine. Euphytica 2017, 213, 68. [Google Scholar] [CrossRef]
- Marsh, E.; Alvarez, S.; Hicks, L.M.; Barbazuk, W.B.; Qiu, W.; Kovacs, L.; Schachtman, D. Changes in protein abundance during powdery mildew infection of leaf tissues of Cabernet Sauvignon grapevine (Vitis vinifera L.). Proteomics 2010, 10, 2057–2064. [Google Scholar] [CrossRef]
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; et al. Glycerolipids in photosynthesis: Composition, synthesis and trafficking. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 470–480. [Google Scholar] [CrossRef]
- Mehta, S.; Chakraborty, A.; Roy, A.; Singh, I.K.; Singh, A. Fight Hard or Die Trying: Current Status of Lipid Signaling during Plant–Pathogen Interaction. Plants 2021, 10, 1098. [Google Scholar] [CrossRef]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Dubots, E.; Audry, M.; Yamaryo, Y.; Bastien, O.; Ohta, H.; Breton, C.; Maréchal, E.; Block, M.A. Activation of the Chloroplast Monogalactosyldiacylglycerol Synthase MGD1 by Phosphatidic Acid and Phosphatidylglycerol. J. Biol. Chem. 2010, 285, 6003–6011. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X. Phospholipase D and phosphatidic acid in plant immunity. Plant Sci. 2019, 279, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Feechan, A.; Dry, I. Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease. Hortic. Res. 2015, 2, 15020. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.A.; Kalisch, B.; Hölzl, G.; Schulze, S.; Thiele, J.; Melzer, M.; Rostond, L.R.; Benning, C.; Dörmann, P. Synthesis and transfer of galactolipids in the chloroplast envelope membranes of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 10714–10719. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.C.; Thordal-Christensen, H.; Lipka, V.; Bau, S.; Kombrink, E.; Qiu, J.-L.; Hückelhoven, R.; Stein, M.; Freialdenhoven, A.; Somerville, S.C.; et al. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 2003, 425, 973–977. [Google Scholar] [CrossRef]
- Lipka, V.; Dittgen, J.; Bednarek, P.; Bhat, R.; Wiermer, M.; Stein, M.; Landtag, J.; Brandt, W.; Rosahl, S.; Scheel, D.; et al. Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis. Science 2005, 310, 1180–1183. [Google Scholar] [CrossRef]
- Stein, M.; Dittgen, J.; Sanchez-Rodriquez, C.; Hou, B.-H.; Molina, A.; Schulze-Lefert, P.; Lipka, V.; Somerville, S. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to non-host resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 2006, 18, 731–746. [Google Scholar] [CrossRef]
- Volynkin, V.; Vasylyk, I.; Volodin, V.; Grigoreva, E.; Karzhaev, D.; Lushchay, E.; Ulianich, P.; Volkov, V.; Risovannaya, V.; Blinova, S.; et al. The Assessment of Agrobiological and Disease Resistance Traits of Grapevine Hybrid Populations (Vitis vinifera L. × Muscadinia rotundifolia Michx.) in the Climatic Conditions of Crimea. Plants 2021, 10, 1215. [Google Scholar] [CrossRef]
- Deliere, L.; Miclot, A.S.; Sauris, P.; Rey, P.; Calonnec, A. Efficacy of fungicides with various modes of action in controlling the early stages of an Erysiphe necator-induced epidemic. Pest Manag. Sci. 2010, 66, 1367–1373. [Google Scholar] [CrossRef]
- Valdés-Gómez, H.; Gary, C.; Cartolaro, P.; Lolas-Caneo, M.; Calonnec, A. Powdery mildew development is positively influenced by grapevine vegetative growth induced by different soil management strategies. Crop Prot. 2011, 30, 1168–1177. [Google Scholar] [CrossRef]
- Miclot, A.S.; Wiedemann-Merdinoglu, S.; Duchêne, E.; Merdinoglu, D.; Mestre, P. A standardised method for the quantitative analysis of resistance to grapevine powdery mildew. Eur. J. Plant Pathol. 2012, 133, 483–495. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2020. Available online: https://www.R-project.org/ (accessed on 2 December 2022).
- Sawilowsky, S.S. New effect size rules of thumb. J. Mod. Appl. Stat. Methods 2009, 8, 26. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Resistance- Related Powdery Mildew Loci (Ren/Run) | Resistance Mechanism within the Hosts | Preliminary Leaf Resistance Level | Source of Resistance | References | |||
|---|---|---|---|---|---|---|---|---|
| PCD | ROS | Callose | ||||||
| mono-locus resistance | BC4 | Run1 | yes | yes | yes | total resistance | M. rotundifolia1 | [20,37] |
| “Kishmish vatkana” | Ren1 | yes | yes | yes | partial resistance | V. vinifera2 | [39] | |
| pyramided resistance | F26P92 | Ren3 | yes | yes | yes | partial resistance | V. rupestris3 | [34,38] |
| Ren9 | yes | n.d. | n.d. | partial resistance | V. rupestris3 | [34,40] | ||
| control | “Teroldego” | - | - | - | - | susceptible | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciubotaru, R.M.; Garcia-Aloy, M.; Masuero, D.; Franceschi, P.; Zulini, L.; Stefanini, M.; Oberhuber, M.; Robatscher, P.; Chitarrini, G.; Vrhovsek, U. Semi-Targeted Profiling of the Lipidome Changes Induced by Erysiphe Necator in Disease-Resistant and Vitis vinifera L. Varieties. Int. J. Mol. Sci. 2023, 24, 4072. https://doi.org/10.3390/ijms24044072
Ciubotaru RM, Garcia-Aloy M, Masuero D, Franceschi P, Zulini L, Stefanini M, Oberhuber M, Robatscher P, Chitarrini G, Vrhovsek U. Semi-Targeted Profiling of the Lipidome Changes Induced by Erysiphe Necator in Disease-Resistant and Vitis vinifera L. Varieties. International Journal of Molecular Sciences. 2023; 24(4):4072. https://doi.org/10.3390/ijms24044072
Chicago/Turabian StyleCiubotaru, Ramona Mihaela, Mar Garcia-Aloy, Domenico Masuero, Pietro Franceschi, Luca Zulini, Marco Stefanini, Michael Oberhuber, Peter Robatscher, Giulia Chitarrini, and Urska Vrhovsek. 2023. "Semi-Targeted Profiling of the Lipidome Changes Induced by Erysiphe Necator in Disease-Resistant and Vitis vinifera L. Varieties" International Journal of Molecular Sciences 24, no. 4: 4072. https://doi.org/10.3390/ijms24044072
APA StyleCiubotaru, R. M., Garcia-Aloy, M., Masuero, D., Franceschi, P., Zulini, L., Stefanini, M., Oberhuber, M., Robatscher, P., Chitarrini, G., & Vrhovsek, U. (2023). Semi-Targeted Profiling of the Lipidome Changes Induced by Erysiphe Necator in Disease-Resistant and Vitis vinifera L. Varieties. International Journal of Molecular Sciences, 24(4), 4072. https://doi.org/10.3390/ijms24044072