
Magnolol Supplementation Alters Serum Parameters, Immune Homeostasis, Amino Acid Profiles, and Gene Expression of Amino Acid Transporters in Growing Pigs

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Growth Performance
2.2. Serum Biochemical Indices
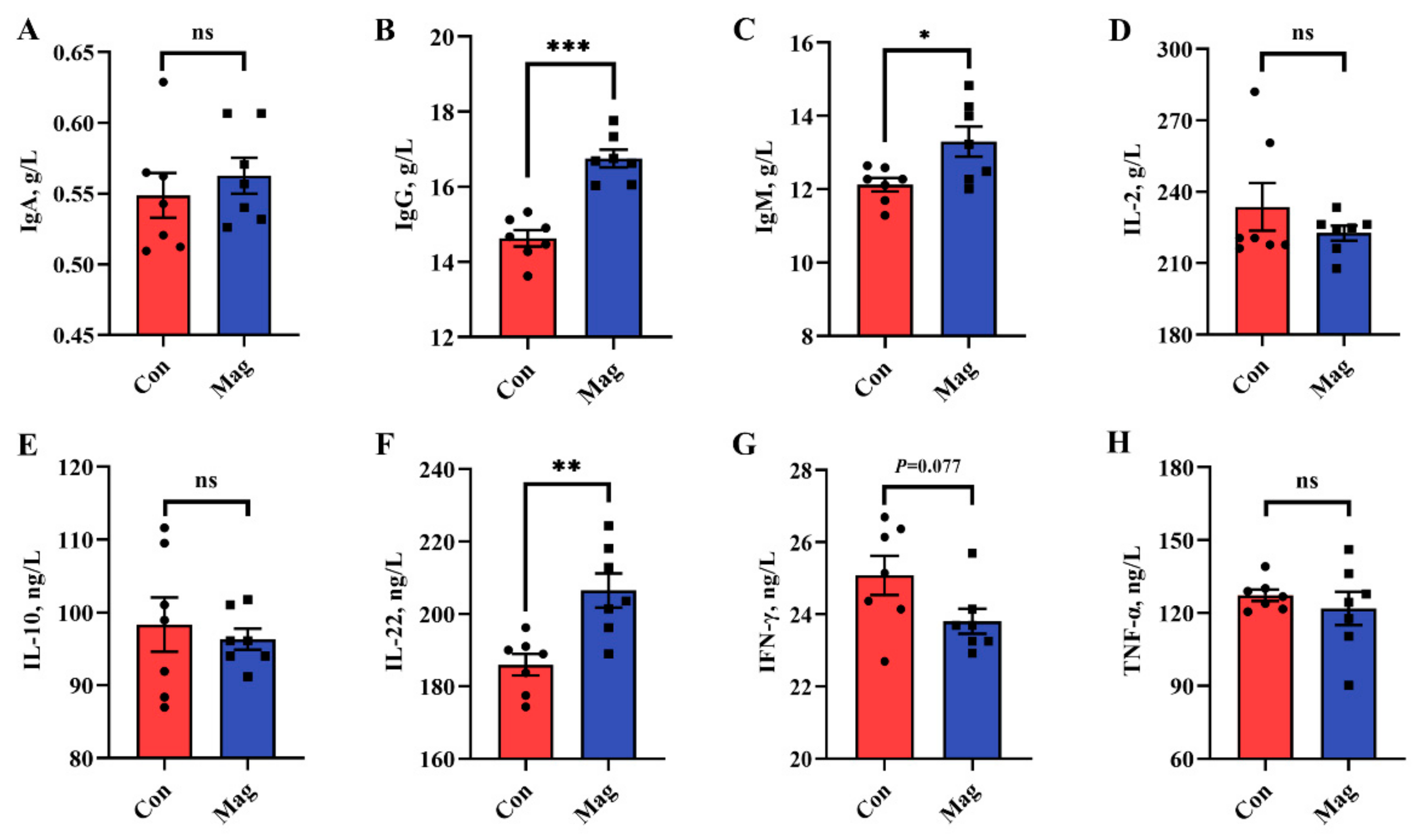
2.3. Immune Responses
2.4. Serum and Dorsal Muscle Amino Acid (AA) Profiles
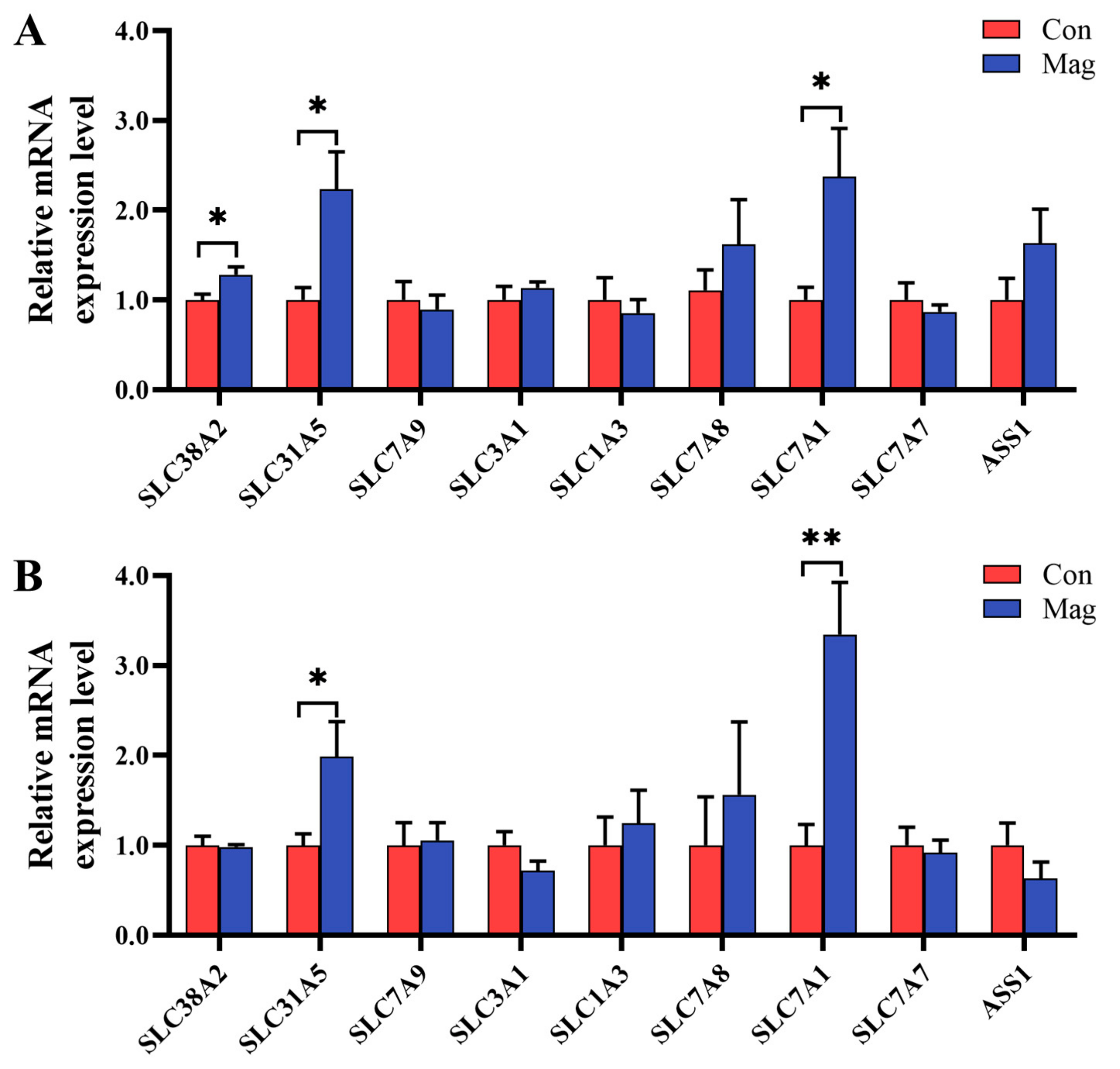
2.5. Gene Expression of AA Transporters in the Jejunum and Ileum
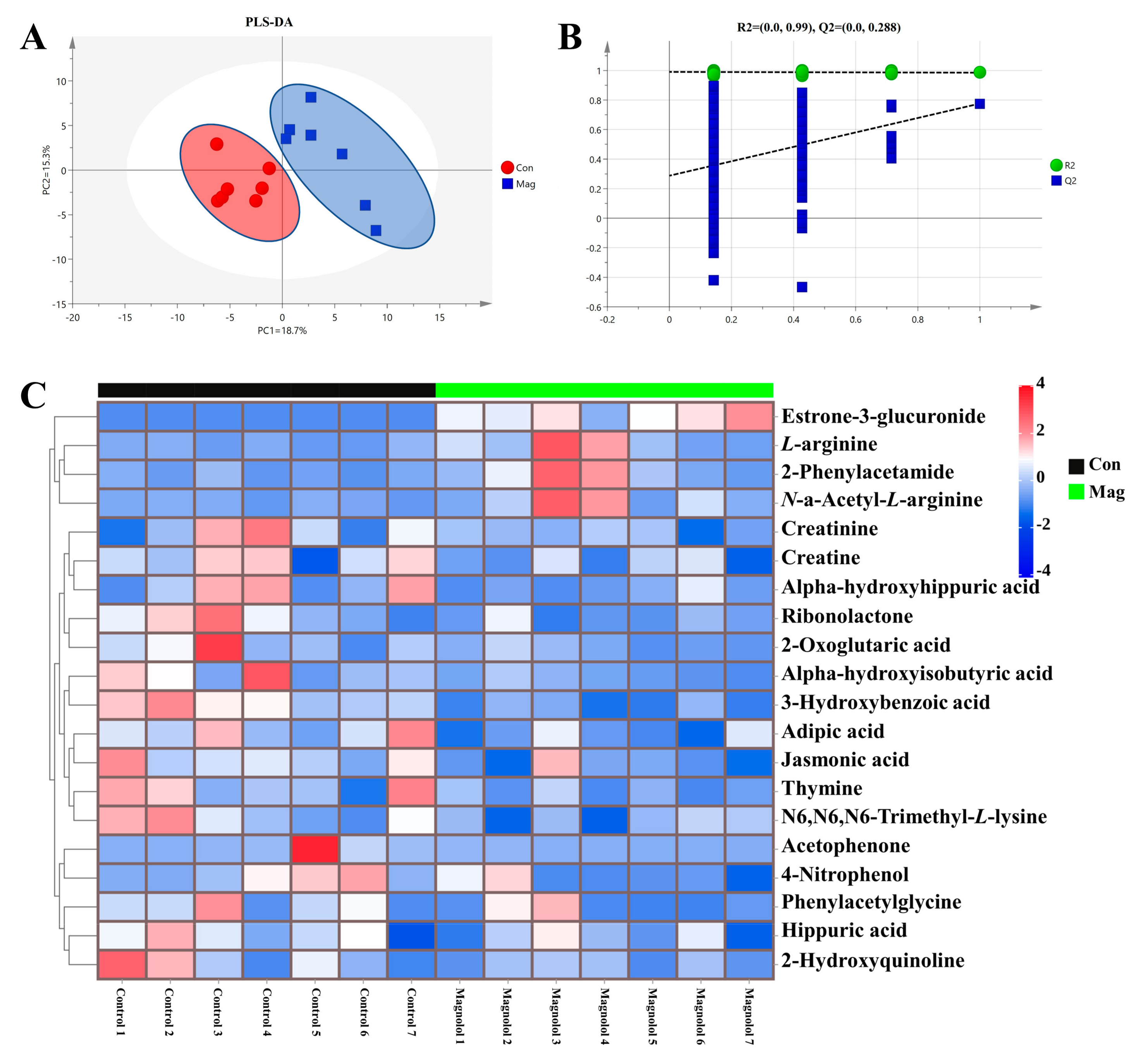
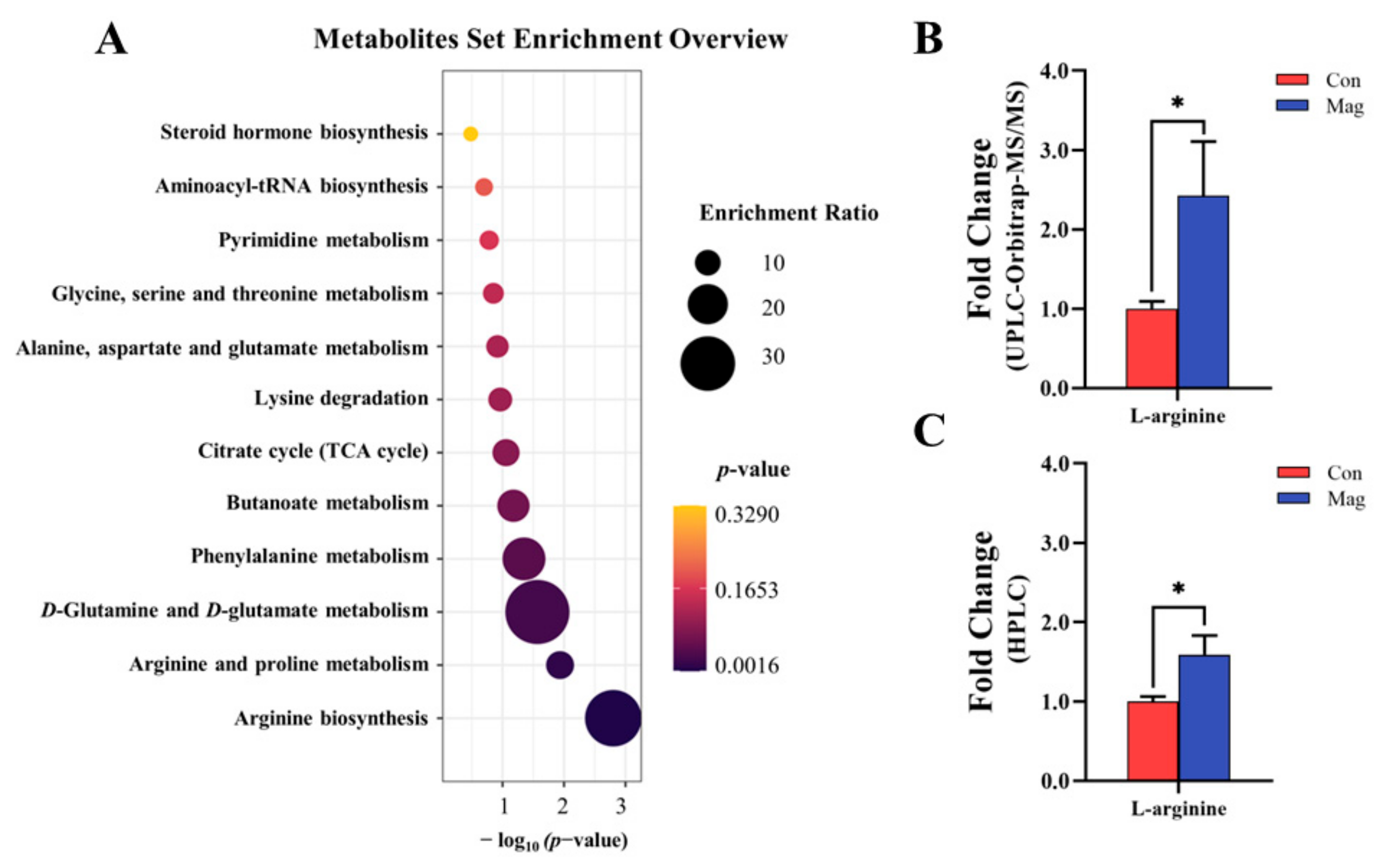
2.6. Serum Metabolic Profiles and Pathway Analysis
3. Discussion
3.1. Effects of Dietary Magnolol Supplementation on the Growth Performance of Growing Pigs
3.2. Effects of Dietary Magnolol Supplementation on Biochemical Indices of Growing Pigs
3.3. Effect of Dietary Magnolol Supplementation on the Immune Response of Growing Pigs
3.4. Effect of Dietary Magnolol Supplementation on the Absorption and Utilization of AAs in Growing Pigs
4. Materials and Methods
4.1. Animal, Diet, and Experimental Design
4.2. Growth Performance
4.3. Sample Collection
4.4. Serum Biochemistry and Immunological Indices
4.5. Free Amino Acids
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.7. Untargeted Serum Metabolomics Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of Antimicrobials in Food Animals and Impact of Transmission of Antimicrobial Resistance on Humans. Biosaf. Health 2021, 3, 32–38. [Google Scholar] [CrossRef]
- Kaiser, R.A.; Taing, L.; Bhatia, H. Antimicrobial Resistance and Environmental Health: A Water Stewardship Framework for Global and National Action. Antibiotics 2022, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, M.L.; Quessy, S.; Letellier, A.; Arsenault, J.; Boulianne, M. Impact of A Drug-free Program on Broiler Chicken Growth Performances, Gut Health, Clostridium perfringens and Campylobacter jejuni Occurrences at the Farm Level. Poult. Sci. 2015, 94, 1791–1801. [Google Scholar] [CrossRef]
- Gaucher, M.-L.; Perron, G.G.; Arsenault, J.; Letellier, A.; Boulianne, M.; Quessy, S. Recurring Necrotic Enteritis Outbreaks in Commercial Broiler Chicken Flocks Strongly Influence Toxin Gene Carriage and Species Richness in the Resident Clostridium Perfringens Population. Front. Microbiol. 2017, 8, 881. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Islam, B.; Akram, M.; Shakil, S.; Ahmad, A.A.; Ali, S.M.; Siddiqui, M.; Khan, A.U. Antimicrobial Activity of Five Herbal Extracts against Multi Drug Resistant (MDR) Strains of Bacteria and Fungus of Clinical Origin. Molecules 2009, 14, 586–597. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential Oils as Alternatives to Antibiotics in Swine Production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Melara, E.G.; Avellaneda, M.C.; Valdivié, M.; García-Hernández, Y.; Aroche, R.; Martínez, Y. Probiotics: Symbiotic Relationship with the Animal Host. Animals 2022, 12, 719. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Peng, Y.; Mu, C.; Zhu, W. Ileum Terminal Antibiotic Infusion Affects Jejunal and Colonic Specific Microbial Population and Immune Status in Growing Pigs. J. Anim. Sci. Biotechnol. 2018, 9, 51. [Google Scholar] [CrossRef]
- Lin, S.; Wang, Z.; Lam, K.L.; Zeng, S.; Tan, B.K.; Hu, J. Role of Intestinal Microecology in the Regulation of Energy Metabolism by Dietary Polyphenols and Their Metabolites. Food Nutr. Res. 2019, 63, 1518. [Google Scholar] [CrossRef]
- Escobar, J.; Frank, J.W.; Suryawan, A.; Nguyen, H.V.; Davis, T.A. Amino Acid Availability and Age Affect the Leucine Stimulation of Protein Synthesis and eIF4F Formation in Muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1615–E1621. [Google Scholar] [CrossRef]
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Parker, D.S.; Armstrong, D.G. Antibiotic Feed Additives and Livestock Production. Proc. Nutr. Soc. 1987, 46, 415–421. [Google Scholar] [CrossRef]
- Yu, M.; Mu, C.; Yang, Y.; Zhang, C.; Su, Y.; Huang, Z.; Yu, K.; Zhu, W. Increases in Circulating Amino Acids with In-feed Antibiotics Correlated with Gene Expression of Intestinal Amino Acid Transporters in Piglets. Amino Acids 2017, 49, 1587–1599. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, D.-C.; Chen, L.-M. Facing a New Challenge: The Adverse Effects of Antibiotics on Gut Microbiota and Host Immunity. Chin. Med. J. 2019, 132, 1135–1138. [Google Scholar] [CrossRef]
- Wang, W.; Li, F.; Duan, Y.; Guo, Q.; Zhang, L.; Yang, Y.; Yin, Y.; Han, M.; Gong, S.; Li, J.; et al. Effects of Dietary Chlorogenic Acid Supplementation Derived from Lonicera Macranthoides Hand-Mazz on Growth Performance, Free Amino Acid Profile, and Muscle Protein Synthesis in a Finishing Pig Model. Oxid. Med. Cell. Longev. 2022, 2022, 6316611. [Google Scholar] [CrossRef]
- Yang, G.; Bibi, S.; Du, M.; Suzuki, T.; Zhu, M.-J. Regulation of the Intestinal Tight Junction by Natural Polyphenols: A Mechanistic Perspective. Crit. Rev. Food Sci. Nutr. 2017, 57, 3830–3839. [Google Scholar] [CrossRef]
- Tretola, M.; Bee, G.; Silacci, P. Gallic Acid Affects Intestinal-epithelial-cell Integrity and Selected Amino-acid Uptake in Porcine In Vitro and Ex Vivo Permeability Models. Brit. J. Nutr. 2021, 126, 492–500. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, W.; Li, Q.; Li, Y.; Yan, Y.; Huang, F.; Wu, X.; Zhou, Q.; Shu, X.; Ruan, Z. Dietary Chlorogenic Acid Regulates Gut Microbiota, Serum-free Amino Acids and Colonic Serotonin Levels in Growing Pigs. Int. J. Food Sci. Nutr. 2018, 69, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Mao, Y.; Wang, S.; Lu, H.; Wang, B.; Ying, L.; Li, Y. Mechanism of Queercetin on Protein Availability in Porcine Intestinal Eptihelial Cells. Chin. J. Anim. Nutr. 2021, 33, 553–562. [Google Scholar]
- Chen, X.; Qiao, T.; Mao, Z.; Jia, G.; Zhao, H.; Liu, G.; Huang, Z. Caffeic Acid Improves Intestinal Barrier Functions by Regulating Colonic Bacteria and Tight Junction Protein Expression and Alleviating Inflammation in Weaning Piglets. Anim. Biotechnol. 2023, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, X.; Zhu, Y.; Wang, K.; Wang, Y. Effect of Magnolol on Cerebral Injury and Blood Brain Barrier Dysfunction Induced by Ischemia-Reperfusion In Vivo and In Vitro. Metab. Brain Dis. 2017, 32, 1109–1118. [Google Scholar] [CrossRef]
- Xie, Q.; Xie, K.; Yi, J.; Song, Z.; Zhang, H.; He, X. The Effects of Magnolol Supplementation on Growth Performance, Meat Quality, Oxidative Capacity, and Intestinal Microbiota in Broilers. Poult. Sci. 2022, 101, 101722. [Google Scholar] [CrossRef]
- Du, E.; Fan, Q.; Zhao, N.; Zhang, W.; Wei, J.; Chen, F.; Huang, S.; Guo, W. Supplemental Magnolol Improves the Antioxidant Capacity and Intestinal Health of Broiler Chickens. Anim. Sci. J. 2021, 92, e13665. [Google Scholar] [CrossRef]
- Peng, W.; Zhou, N.; Song, Z.; Zhang, H.; He, X. Magnolol as a Protective Antioxidant Alleviates Rotenone-Induced Oxidative Stress and Liver Damage through MAPK/mTOR/Nrf2 in Broilers. Metabolites 2023, 13, 84. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, H.; Zhao, N.; Du, E.; Jin, F.; Fan, Q.; Guo, W.; Huang, S.; Wei, J. Effects of Magnolol and Honokiol Blend on Performance, Egg Quality, Hepatic Lipid Metabolism, and Intestinal Morphology of Hens at Late Laying Cycle. Animal 2022, 16, 100532. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Strapazzon, J.V.; Marchiori, M.S.; Molosse, V.L.; Deolindo, G.L.; Fracasso, M.; Copetti, P.M.; Morsch, V.M.; Baldissera, M.D.; Mendes, R.E.; et al. Addition of a Blend Based on Zinc Chloride and Lignans of Magnolia in the Diet of Broilers to Substitute for a Conventional Antibiotic: Effects on Intestinal Health, Meat Quality, and Performance. Animals 2022, 12, 3249. [Google Scholar] [CrossRef]
- Xie, N.; Hu, C.; Guo, A.; Liang, H.; Du, P.; Yin, G. Metabolic Regulation of Magnolol on the Nuclear Receptor, Liver X Receptor. Exp. Ther. Med. 2015, 9, 1827–1830. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Li, Y.; Tian, Q.; Li, Z.; Rong, T.; Ma, X.; Tian, Z.; Cui, Y.; Yu, M. Effects of Magnolol on Nutrient Apparent Digestibility, Serum Biochemical Indices, Main Fceal Microorganism Numbers and Their Metabolite Contents of Weaned Piglets. Chin. J. Anim. Nutr. 2022, 34, 4919–4931. [Google Scholar]
- Li, Y.; Li, F.; Duan, Y.; Guo, Q.; Wang, W.; Wen, C.; Huang, X.; Yin, Y. The Protein and Energy Metabolic Response of Skeletal Muscle to the Low-Protein Diets in Growing Pigs. J. Agric. Food Chem. 2017, 65, 8544–8551. [Google Scholar] [CrossRef]
- Lin, Q.; Liu, Y.; Peng, S.; Liu, C.; Lv, T.; Liao, L.; Li, Y.; Wang, Y.; Fan, Z.; Wu, W.; et al. Magnolol Additive Improves Growth Performance of Linwu Ducklings by Modulating Antioxidative Status. PLoS ONE 2022, 16, e0259896. [Google Scholar] [CrossRef]
- Menci, R.; Khelil-Arfa, H.; Blanchard, A.; Biondi, L.; Bella, M.; Priolo, A.; Luciano, G.; Natalello, A. Effect of Dietary Magnolia Bark Extract Supplementation in Finishing Pigs on the Oxidative Stability of Meat. J. Anim. Sci. Biotechnol. 2022, 13, 89. [Google Scholar] [CrossRef]
- Simitzis, P.E.; Symeon, G.K.; Charismiadou, M.A.; Bizelis, J.A.; Deligeorgis, S.G. The Effects of Dietary Oregano Oil Supplementation on Pig Meat Characteristics. Meat Sci. 2010, 84, 670–676. [Google Scholar] [CrossRef]
- Yang, Y.; Li, F.; Guo, Q.; Wang, W.; Zhang, L.; Yin, Y.; Gong, S.; Han, M.; Yin, Y. Effects of Different Supplemental Levels of Eucommia ulmoides Leaf Extract in the Diet on Carcass Traits and Lipid Metabolism in Growing–Finishing Pigs. Front. Vet. Sci. 2022, 8, 828165. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.; Burrin, D.G.; Henry, J.; Yu, H.; Jahoor, F.; Reeds, P.J. Dietary Amino Acids Are the Preferential Source of Hepatic Protein Synthesis in Piglets. J. Nutr. 1998, 128, 1517–1524. [Google Scholar] [CrossRef]
- Li, X.; Xiong, X.; Wu, X.; Liu, G.; Zhou, K.; Yin, Y. Effects of Stocking Density on Growth Performance, Blood Parameters and Immunity of Growing Pigs. Anim. Nutr. 2020, 6, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-H.; Onufer, E.J.; Huang, L.-H.; Sprung, R.W.; Davidson, W.S.; Czepielewski, R.S.; Wohltmann, M.; Sorci-Thomas, M.G.; Warner, B.W.; Randolph, G.J. Enterically Derived High-density Lipoprotein Restrains Liver Injury through the Portal Vein. Science 2021, 373, eabe6729. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, H.; Weng, M.; Jiang, S.; Gao, J. Diagnostic and Clinical Significance of Serum Levels of D-Lactate and Diamine Oxidase in Patients with Crohn’s Disease. Gastroent. Res. Pract. 2019, 2019, 8536952. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Han, X.; Tang, S.; Li, C.; Xiao, W.; Tan, Z. Magnolol and Honokiol Attenuate Apoptosis of Enterotoxigenic Escherichia Coli-Induced Intestinal Epithelium by Maintaining Secretion and Absorption Homeostasis and Protecting Mucosal Integrity. Med. Sci. Monit. 2018, 24, 3348–3356. [Google Scholar] [CrossRef]
- Khan, S.R.; van der Burgh, A.C.; Peeters, R.P.; van Hagen, P.M.; Dalm, V.A.S.H.; Chaker, L. Determinants of Serum Immunoglobulin Levels: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 1103. [Google Scholar] [CrossRef]
- Ding, X.; Zhong, X.; Yang, Y.; Zhang, G.; Si, H. Citric Acid and Magnolol Ameliorate Clostridium Perfringens Challenge in Broiler Chickens. Animals 2023, 13, 577. [Google Scholar] [CrossRef]
- Huang, H.; Su, L.; Sun, Y.; Li, B.; Deng, J.; Cao, Z. Dietary Magnolol Inclusion Improves the Antioxidant and Immune Responses, and Resistance to Aeromonas Hydrophila in Genetically Improved Farmed Tilapia (Oreochromis niloticus). Aquacult. Rep. 2022, 23, 101017. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Moghadam, M.S.; Dawood, M.A.O.; Hoseinifar, S.H. Lactobacillus Fermentum and/or Ferulic Acid Improved the Immune Responses, Antioxidative Defence and Resistance against Aeromonas Hydrophila in Common Carp (Cyprinus carpio) Fingerlings. Fish Shellfish Immunol. 2019, 94, 916–923. [Google Scholar] [CrossRef]
- Sahu, S.; Das, B.K.; Pradhan, J.; Mohapatra, B.C.; Mishra, B.K.; Sarangi, N. Effect of Magnifera Indica Kernel as a Feed Additive on Immunity and Resistance to Aeromonas Hydrophila in Labeo Rohita Fingerlings. Fish Shellfish Immunol. 2007, 23, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Lin, H.; Ge, X.; Niu, J.; Wang, J.; Wang, Y.; Chen, L.; Huang, Z.; Yu, W.; Tan, X. The Effects of Dietary Soybean Isoflavones on Growth, Innate Immune Responses, Hepatic Antioxidant Abilities and Disease Resistance of Juvenile Golden Pompano Trachinotus Ovatus. Fish Shellfish Immunol. 2015, 43, 158–166. [Google Scholar] [CrossRef]
- Sabat, R.; Ouyang, W.; Wolk, K. Therapeutic Opportunities of the IL-22–IL-22R1 System. Nat. Rev. Drug Discovery 2014, 13, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Yi, T.; Lu, T.T.; Ghilardi, N. The Role of IL-22 in Intestinal Health and Disease. J. Exp. Med. 2020, 217, e20192195. [Google Scholar] [CrossRef]
- Pennino, D.; Bhavsar, P.K.; Effner, R.; Avitabile, S.; Venn, P.; Quaranta, M.; Marzaioli, V.; Cifuentes, L.; Durham, S.R.; Cavani, A.; et al. IL-22 Suppresses IFN-γ–Mediated Lung Inflammation in Asthmatic Patients. J. Allergy Clin. Immun. 2013, 131, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Kong, R.; Han, W.; Bao, W.; Shi, Y.; Ye, L.; Lu, J. Honokiol Alleviates Ulcerative Colitis by Targeting PPAR-γ–TLR4–NF-κB Signaling and Suppressing Gasdermin-D-mediated Pyroptosis In Vivo and In Vitro. Int. Immunopharmacol. 2022, 111, 109058. [Google Scholar] [CrossRef]
- Lee, J.; Jung, E.; Park, J.; Jung, K.; Lee, S.; Hong, S.; Park, J.; Park, E.; Kim, J.; Park, S.; et al. Anti-Inflammatory Effects of Magnolol and Honokiol are Mediated through Inhibition of the Downstream Pathway of MEKK-1 in NF-κB Activation Signaling. Planta Med. 2005, 71, 338–343. [Google Scholar] [CrossRef]
- Wu, G. Functional Amino Acids in Nutrition and Health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef]
- Chen, X.; Luo, X.; Chen, D.; Yu, B.; He, J.; Huang, Z. Arginine Promotes Porcine Type I Muscle Fibres Formation through Improvement of Mitochondrial Biogenesis. Brit. J. Nutr. 2020, 123, 499–507. [Google Scholar] [CrossRef]
- Martí i Líndez, A.-A.; Reith, W. Arginine-dependent Immune Responses. Cell. Mol. Life Sci. 2021, 78, 5303–5324. [Google Scholar] [CrossRef]
- Davis, T.A.; Fiorotto, M.L. Regulation of Muscle Growth in Neonates. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.M. Role of Amino Acid Transporters in Amino Acid Sensing. Am. J. Clin. Nutr. 2013, 99, 223S–230S. [Google Scholar] [CrossRef]
- Kawase, T.; Ohki, R.; Shibata, T.; Tsutsumi, S.; Kamimura, N.; Inazawa, J.; Ohta, T.; Ichikawa, H.; Aburatani, H.; Tashiro, F. PH Domain-only Protein PHLDA3 Is a p53-regulated Repressor of Akt. Cell 2009, 136, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wu, G. L-glutamate Nutrition and Metabolism in Swine. Amino Acids 2018, 50, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Young, V.R.; Ajami, A.M. Glutamate: An Amino Acid of Particular Distinction. J. Nutr. 2000, 130, 892S–900S. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, R.; Knabe, D.A.; Tekwe, C.D.; Dahanayaka, S.; Ficken, M.D.; Fielder, S.E.; Eide, S.J.; Lovering, S.L.; Wu, G. Dietary Supplementation with Monosodium Glutamate is Safe and Improves Growth Performance in Postweaning Pigs. Amino Acids 2013, 44, 911–923. [Google Scholar] [CrossRef]
- Reeds, P.J.; Burrin, D.G.; Jahoor, F.; Wykes, L.; Henry, J.; Frazer, E.M. Enteral Glutamate is Almost Completely Metabolized in First Pass by the Gastrointestinal Tract of Infant Pigs. Am. J. Physiol. 1996, 270 Pt 1, E413–E418. [Google Scholar] [CrossRef]
- Wu, G.; Morris, S.M., Jr. Arginine Metabolism: Nitric Oxide and Beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef]
- Li, M.; Wen, H.; Huang, F.; Wu, M.; Yu, L.; Jiang, M.; Lu, X.; Tian, J. Role of Arginine Supplementation on Muscular Metabolism and Flesh Quality of Pacific White Shrimp (Litopenaeus vannamei) Reared in Freshwater. Front. Nutr. 2022, 9, 980188. [Google Scholar] [CrossRef] [PubMed]
- Pohlenz, C.; Buentello, A.; le J Helland, S.; Gatlin III, D.M. Effects of Dietary Arginine Supplementation on Growth, Protein Optimization and Innate Immune Response of Channel Catfish Ictalurus punctatus (Rafinesque 1818). Aquac. Res. 2014, 45, 491–500. [Google Scholar] [CrossRef]
- Shaffer, P.L.; Goehring, A.; Shankaranarayanan, A.; Gouaux, E. Structure and Mechanism of a Na+-Independent Amino Acid Transporter. Science 2009, 325, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Fairweather, S.J. Amino Acid Transport Across the Mammalian Intestine. Compr. Physiol. 2018, 9, 343–373. [Google Scholar]
- Mastrototaro, L.; Sponder, G.; Saremi, B.; Aschenbach, J.R. Gastrointestinal Methionine Shuttle: Priority Handling of Precious Goods. IUBMB Life 2016, 68, 924–934. [Google Scholar] [CrossRef]
- Chafai, A.; Fromm, M.F.; König, J.; Maas, R. The Prognostic Biomarker L-homoarginine Is A Substrate of the Cationic Amino Acid Transporters CAT1, CAT2A and CAT2B. Sci. Rep. 2017, 7, 4767. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Thompson, M.J.; Atwood, C.S.; Bird, P.H.; Williams, I.H.; Hartmann, P.E. Maintenance of Villus Height and Crypt Depth, and Enhancement of Disaccharide Digestion and Monosaccharide Absorption, in Piglets Fed on Cows’ Whole Milk after Weaning. Br. J. Nutr. 1996, 76, 409–422. [Google Scholar] [CrossRef]
- Yang, K.; Deng, X.; Jian, S.; Zhang, M.; Wen, C.; Xin, Z.; Zhang, L.; Tong, A.; Ye, S.; Liao, P.; et al. Gallic Acid Alleviates Gut Dysfunction and Boosts Immune and Antioxidant Activities in Puppies Under Environmental Stress Based on Microbiome–Metabolomics Analysis. Front. Immunol. 2022, 12, 813890. [Google Scholar] [CrossRef]
- Zhao, L.; Xiao, H.-t.; Mu, H.-x.; Huang, T.; Lin, Z.-s.; Zhong, L.L.D.; Zeng, G.-z.; Fan, B.-m.; Lin, C.-y.; Bian, Z.-x. Magnolol, a Natural Polyphenol, Attenuates Dextran Sulfate Sodium-Induced Colitis in Mice. Molecules 2017, 22, 1218. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, C.; Zheng, M.; Gao, F.; Zhang, J.; Liu, F. Metabolomics Analysis of L-Arginine Induced Gastrointestinal Motility Disorder in Rats Using UPLC-MS after Magnolol Treatment. Front. Pharmacol. 2019, 10, 183. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Deng, D.; Cui, Y.; Chen, W.; Yu, M.; Ma, X. Diet Supplemented with Fermented Okara Improved Growth Performance, Meat Quality, and Amino Acid Profiles in Growing Gigs. Food Sci. Nutr. 2020, 8, 5650–5659. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Jian, S.; Guo, D.; Wen, C.; Xin, Z.; Zhang, L.; Kuang, T.; Wen, J.; Yin, Y.; Deng, B. Fecal Microbiota and Metabolomics Revealed the Effect of Long-term Consumption of Gallic Acid on Canine Lipid Metabolism and Gut Health. Food Chem. X 2022, 15, 100377. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | Con 2 | Mag 3 | p-Value |
|---|---|---|---|
| Initial BW (kg) (day 0) | 25.32 ± 0.04 | 25.31 ± 0.02 | 0.841 |
| final BW (kg) (day 35) | 50.00 ± 1.03 | 51.30 ± 0.76 | 0.334 |
| ADFI, g/d | 1526.14 ± 19.42 | 1521.79 ± 40.50 | 0.907 |
| ADG, g/d | 719.52 ± 18.04 | 742.31 ± 21.67 | 0.444 |
| F:G | 2.13 ± 0.04 | 2.05 ± 0.03 | 0.132 |
| Items 1 | Con 2 | Mag 3 | p-Value |
|---|---|---|---|
| TP, g/L | 60.86 ± 1.62 | 65.79 ± 1.28 * | 0.049 |
| LDL-C, mmol/L | 0.86 ± 0.03 | 0.81 ± 0.05 | 0.394 |
| HDL-C, mmol/L | 0.73 ± 0.03 | 0.83 ± 0.03 | 0.055 |
| ALT, U/L | 47.29 ± 8.14 | 32.82 ± 2.75 | 0.161 |
| AST, U/L | 78.33 ± 18.94 | 41.99 ± 3.77 | 0.107 |
| ALB, g/L | 19.61 ± 0.71 | 22.41 ± 0.81 * | 0.033 |
| GLU, mmol/L | 4.83 ± 0.28 | 5.07 ± 0.22 | 0.530 |
| UREA, mmol/L | 7.74 ± 0.44 | 7.81 ± 0.75 | 0.937 |
| UA, μmol/L | 17.29 ± 1.65 | 15.66 ±1.99 | 0.592 |
| DAO, U/L | 14.47 ± 1.86 | 9.98 ± 0.68 | 0.057 |
| Items | Con 1 | Mag 2 | p-Value |
|---|---|---|---|
| Arginine | 122.40 ± 6.86 | 194.53 ± 27.29 * | 0.035 |
| Histidine | 56.35 ± 2.75 | 93.53 ± 22.60 | 0.180 |
| Isoleucine | 72.55 ± 7.18 | 92.80 ± 15.17 | 0.286 |
| Leucine | 153.83 ± 10.72 | 208.17 ± 25.43 | 0.101 |
| Lysine | 136.24 ± 9.01 | 205.28 ± 50.06 | 0.233 |
| Methionine | 112.29 ± 5.38 | 129.03 ± 5.61 | 0.070 |
| Phenylalanine | 73.04 ± 5.33 | 111.25 ± 26.30 | 0.232 |
| Threonine | 109.64 ± 5.51 | 133.00 ± 28.01 | 0.463 |
| Tryptophan | 42.69 ± 7.31 | 65.17 ± 19.92 | 0.357 |
| Valine | 226.16 ± 6.28 | 254.21 ± 18.24 | 0.212 |
| Alanine | 553.29 ± 56.70 | 618.18 ± 55.30 | 0.463 |
| Aspartate | 34.02 ± 5.76 | 56.09 ± 8.60 | 0.075 |
| Glutamate | 309.12 ± 45.53 | 331.30 ± 28.19 | 0.709 |
| Glycine | 853.82 ± 91.07 | 1081.41 ± 35.42 | 0.120 |
| Serine | 119.25 ± 10.83 | 147.63 ± 13.76 | 0.161 |
| Tyrosine | 63.00 ± 5.26 | 94.68 ± 17.57 | 0.136 |
| Asparagine | 50.39 ± 6.13 | 81.86 ± 17.38 | 0.140 |
| Glutamine | 402.68 ± 21.93 | 483.32 ± 43.07 | 0.148 |
| Proline | 249.14 ± 25.20 | 353.71 ± 35.42 * | 0.048 |
| Cysteine | 83.26 ± 5.25 | 96.85 ± 2.82 | 0.063 |
| Essential Amino Acids 3 | 1105.20 ± 29.84 | 1339.67 ± 130.17 | 0.136 |
| Nonessential Amino Acids 4 | 2468.83 ± 203.00 | 2991.32 ± 234.98 | 0.146 |
| Total Amino Acids 5 | 3574.03 ± 217.64 | 4709.96 ± 575.24 | 0.127 |
| Items | Con 1 | Mag 2 | p-Value |
|---|---|---|---|
| Arginine | 1.81 ± 0.21 | 2.60 ± 0.47 | 0.193 |
| Histidine | 0.76 ± 0.05 | 1.10 ± 0.17 | 0.098 |
| Isoleucine | 1.16 ± 0.12 | 1.33 ± 0.14 | 0.414 |
| Leucine | 2.53 ± 0.19 | 3.93 ± 0.65 | 0.099 |
| Lysine | 2.01 ± 0.31 | 2.64 ± 0.63 | 0.426 |
| Methionine | 1.95 ± 0.19 | 2.43 ± 0.09 | 0.067 |
| Phenylalanine | 2.37 ± 0.19 | 2.77 ± 0.32 | 0.339 |
| Threonine | 1.42 ± 0.15 | 1.81 ± 0.22 | 0.205 |
| Tryptophan | 0.65 ± 0.06 | 0.75 ± 0.04 | 0.231 |
| Valine | 1.92 ± 0.05 | 2.89± 0.77 | 0.288 |
| Alanine | 14.45 ± 0.78 | 17.06 ± 0.93 | 0.070 |
| Aspartate | 2.14 ± 0.21 | 1.98 ± 0.33 | 0.709 |
| Glutamate | 4.71 ± 0.34 | 7.62 ± 0.88 * | 0.015 |
| Glycine | 14.98 ± 1.38 | 13.59 ± 1.09 | 0.481 |
| Serine | 2.20 ± 0.16 | 2.58 ± 0.18 | 0.170 |
| Tyrosine | 2.01 ± 0.12 | 2.64 ± 0.20 * | 0.031 |
| Asparagine | 0.96 ± 0.08 | 1.35 ± 0.23 | 0.173 |
| Glutamine | 21.29 ± 1.71 | 21.32 ± 1.53 | 0.990 |
| Proline | 4.26 ± 0.57 | 4.31 ± 0.32 | 0.953 |
| Cysteine | 0.25 ± 0.03 | 0.35 ± 0.04 | 0.055 |
| Essential Amino Acids 3 | 18.03 ± 0.88 | 21.23 ± 2.81 | 0.348 |
| Nonessential Amino Acids 4 | 62.17 ± 4.44 | 71.55 ± 3.69 | 0.159 |
| Total Amino Acids 5 | 78.80 ± 4.23 | 97.60 ± 6.69 | 0.052 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, Y.; Yu, M.; Tian, Z.; Deng, J.; Ma, X.; Yin, Y. Magnolol Supplementation Alters Serum Parameters, Immune Homeostasis, Amino Acid Profiles, and Gene Expression of Amino Acid Transporters in Growing Pigs. Int. J. Mol. Sci. 2023, 24, 13952. https://doi.org/10.3390/ijms241813952
Liu Y, Li Y, Yu M, Tian Z, Deng J, Ma X, Yin Y. Magnolol Supplementation Alters Serum Parameters, Immune Homeostasis, Amino Acid Profiles, and Gene Expression of Amino Acid Transporters in Growing Pigs. International Journal of Molecular Sciences. 2023; 24(18):13952. https://doi.org/10.3390/ijms241813952
Chicago/Turabian StyleLiu, Yanchen, Yuanfei Li, Miao Yu, Zhimei Tian, Jinping Deng, Xianyong Ma, and Yulong Yin. 2023. "Magnolol Supplementation Alters Serum Parameters, Immune Homeostasis, Amino Acid Profiles, and Gene Expression of Amino Acid Transporters in Growing Pigs" International Journal of Molecular Sciences 24, no. 18: 13952. https://doi.org/10.3390/ijms241813952
APA StyleLiu, Y., Li, Y., Yu, M., Tian, Z., Deng, J., Ma, X., & Yin, Y. (2023). Magnolol Supplementation Alters Serum Parameters, Immune Homeostasis, Amino Acid Profiles, and Gene Expression of Amino Acid Transporters in Growing Pigs. International Journal of Molecular Sciences, 24(18), 13952. https://doi.org/10.3390/ijms241813952