
Spaceflight-Induced Gene Expression Profiles in the Mouse Brain Are Attenuated by Treatment with the Antioxidant BuOE
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
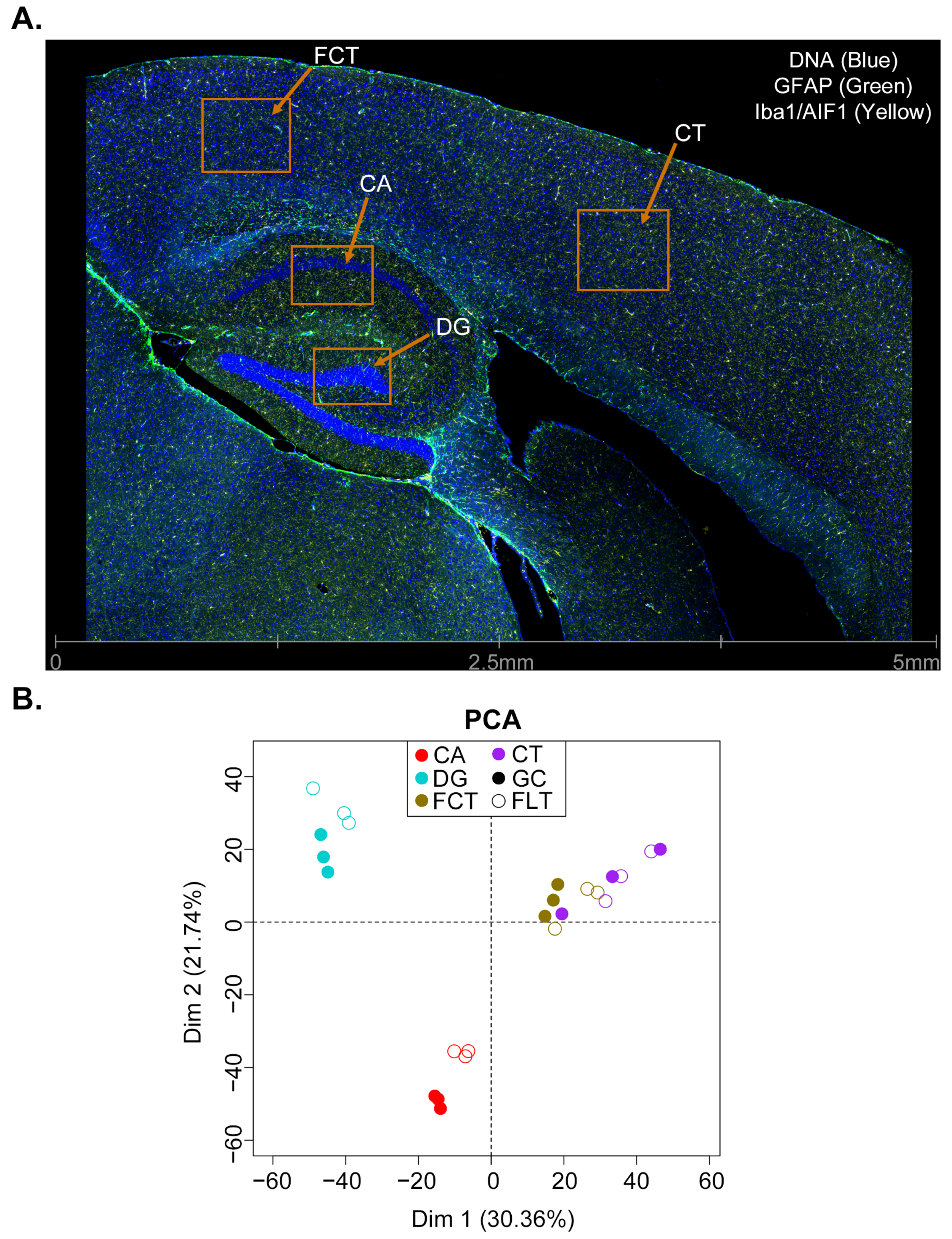
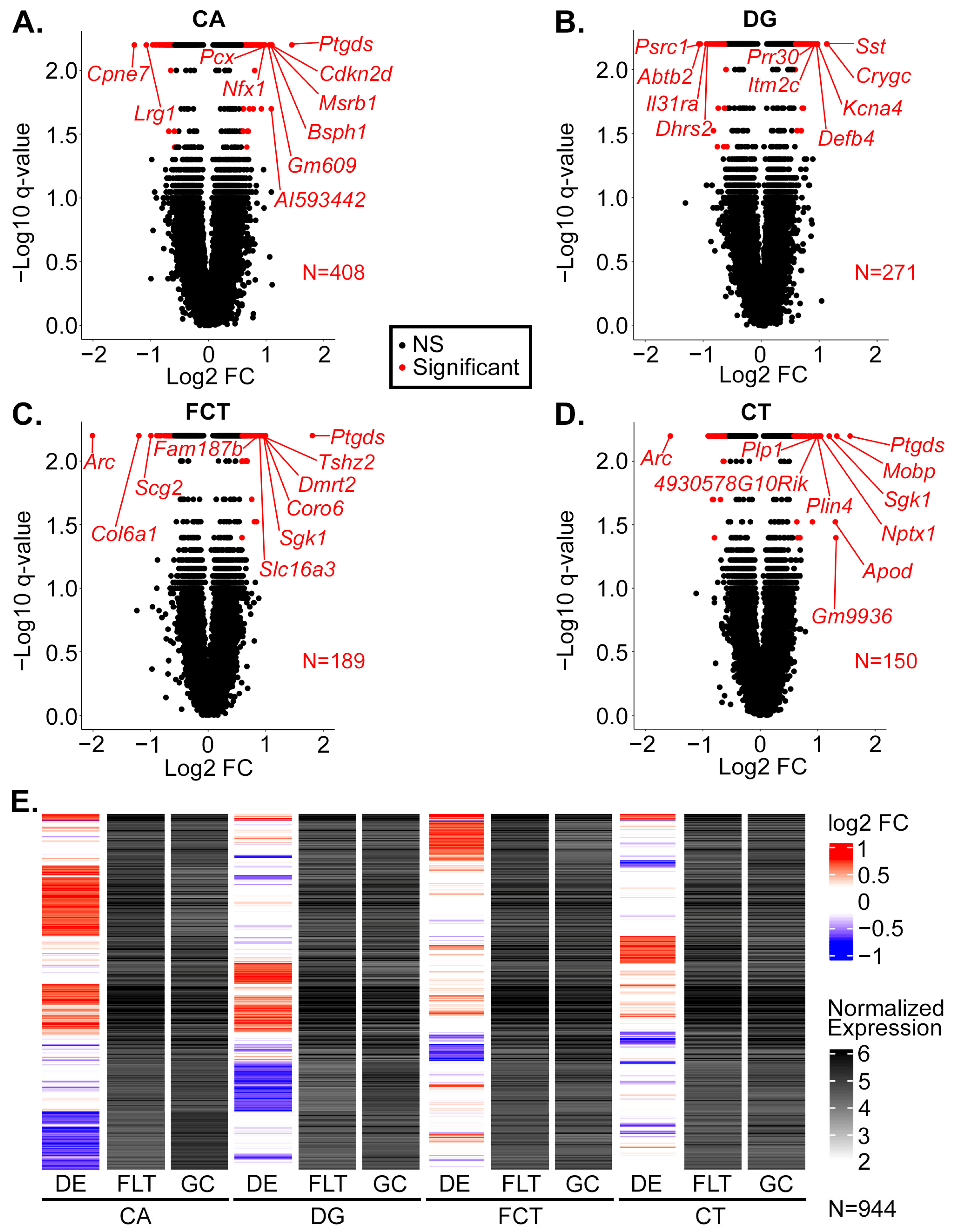
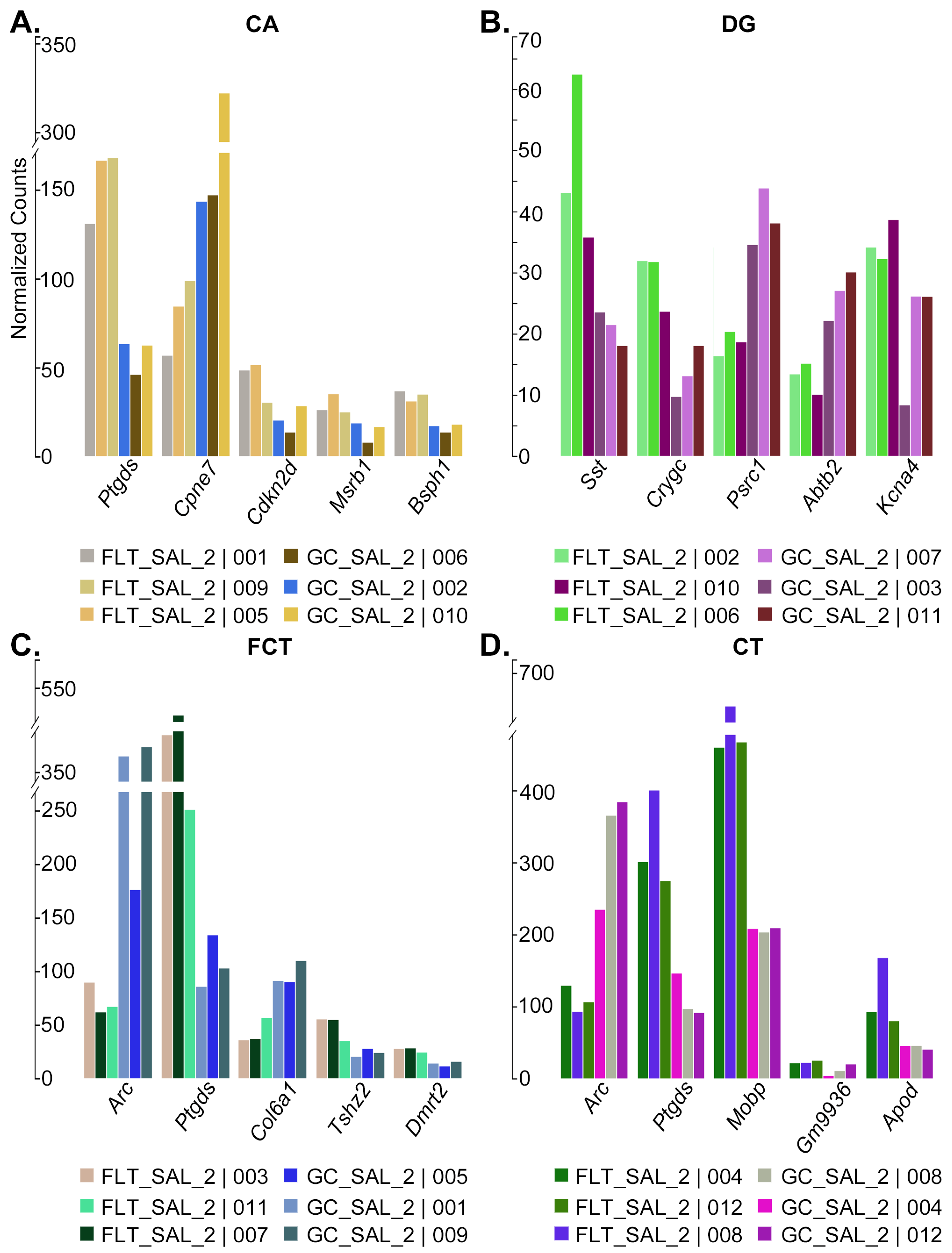
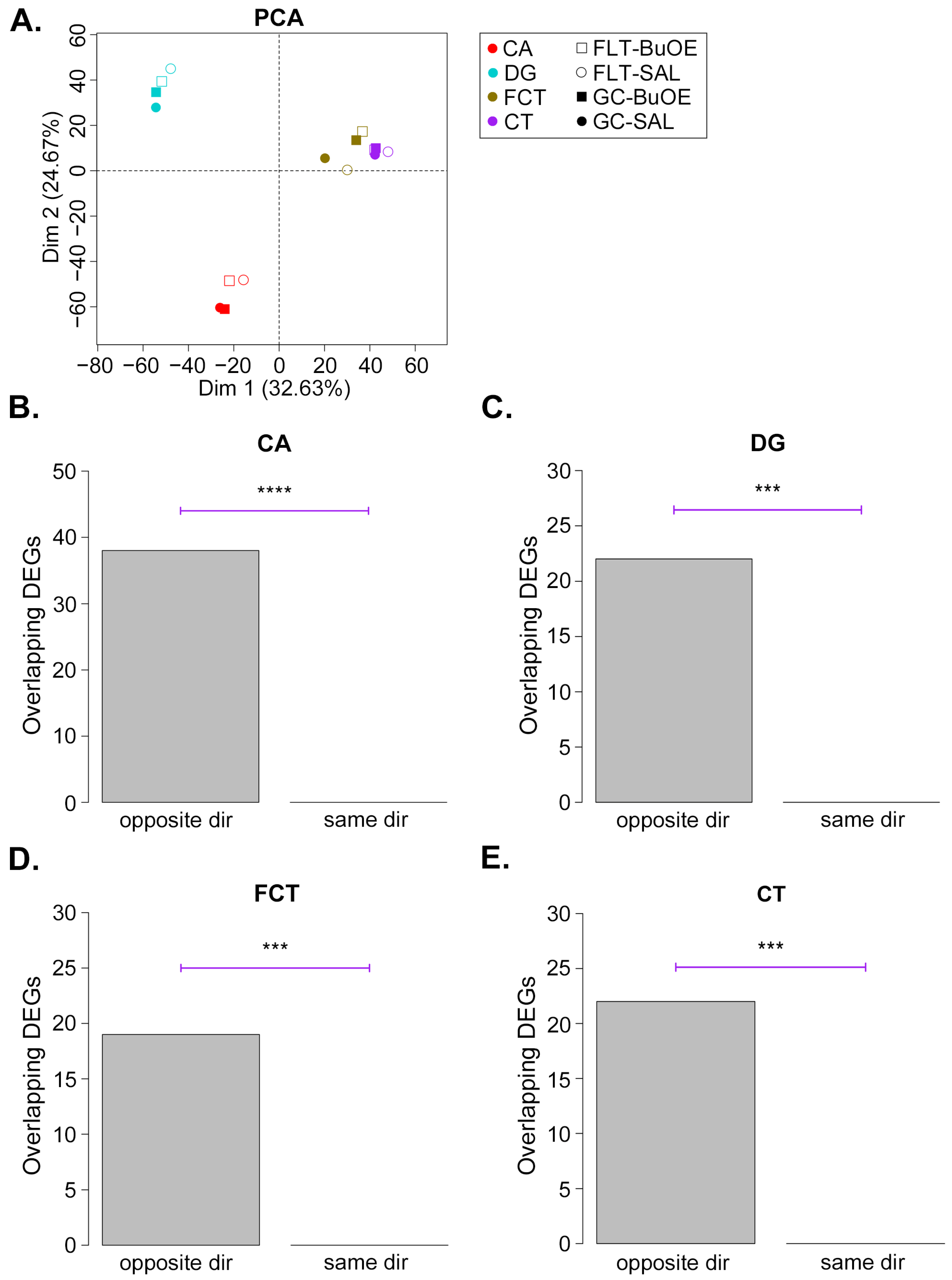
2.1. Gene Expression Profiling of CA, DG, CT, and FCT Samples Subjected to Spaceflight
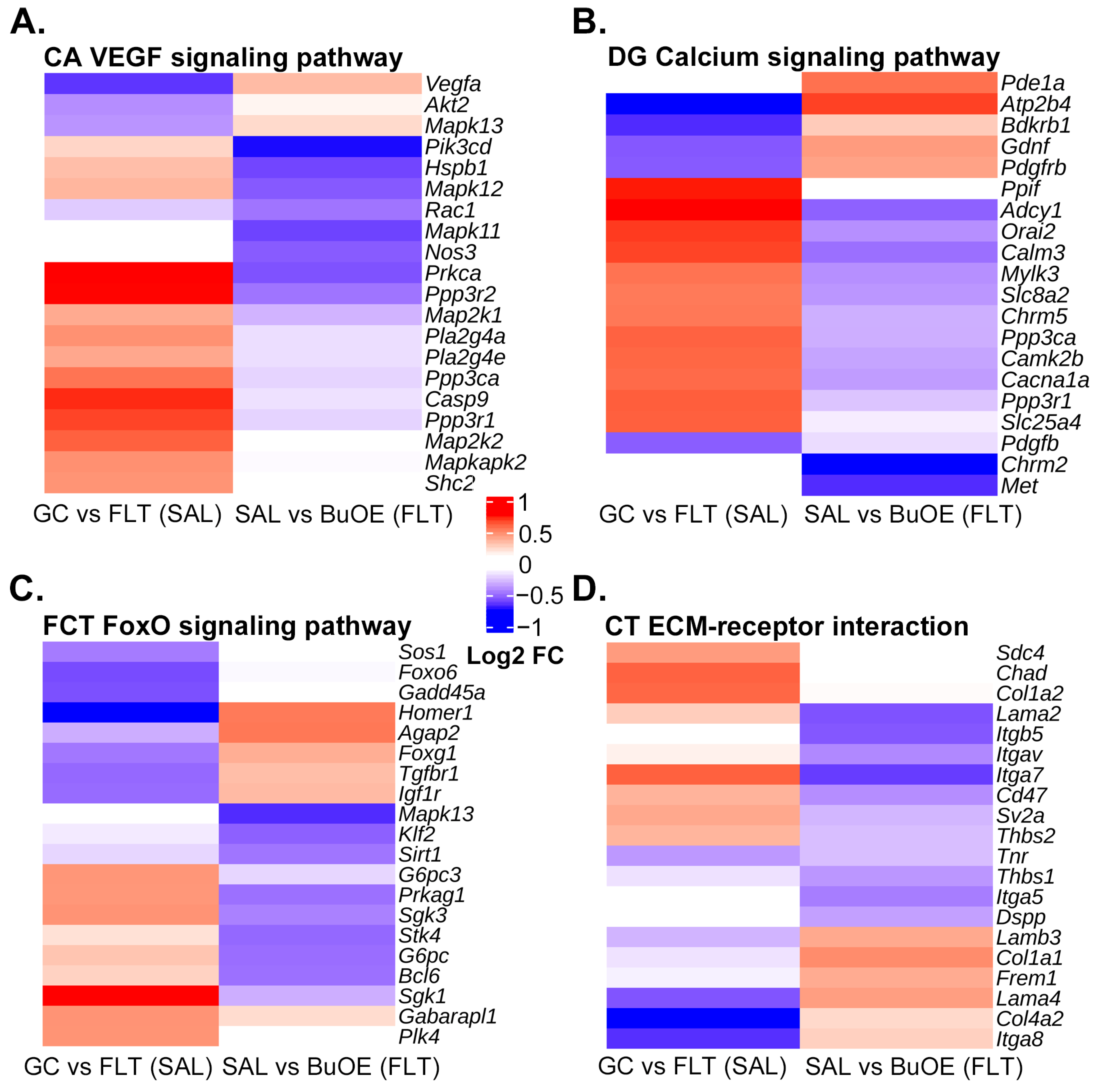
2.2. BuOE Attenuates Spaceflight-Induced Transcriptional Changes in the CA, DG, FCT, and CT
3. Discussion
4. Materials and Methods
4.1. Spaceflight and Mouse Groups
4.2. GeoMx DSP
4.3. GeoMx DSP Data Analysis
4.4. Association of DEG’s with Neurological Disease
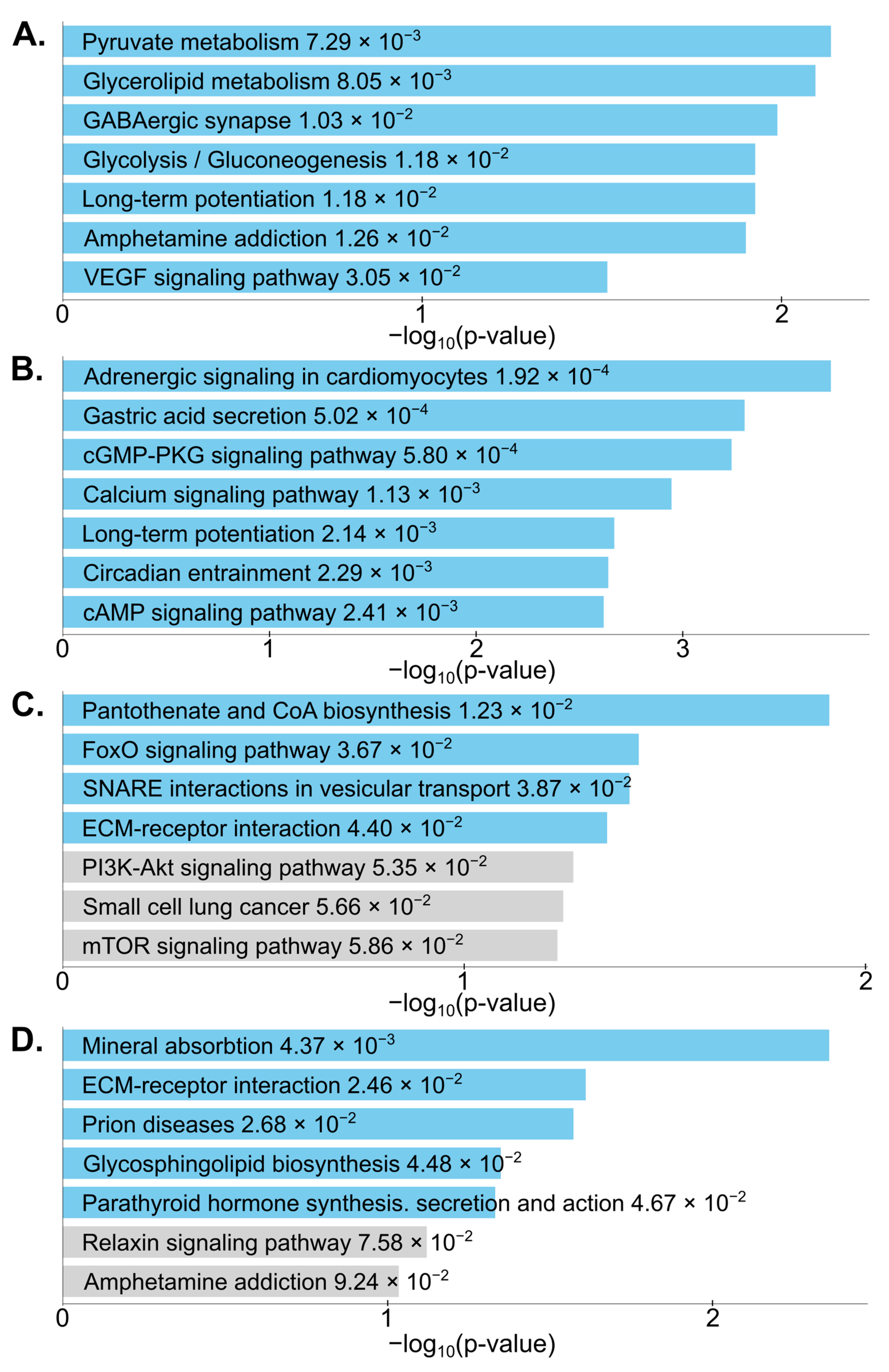
4.5. Pathway Analysis
4.6. Principal Component Analysis
4.7. Statistical Comparison of Spaceflight-Induced and BuOE-Induced DEG Overlaps
4.8. Permutation Testing of Spaceflight-Induced and BuOE-Induced DEG Overlaps
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crucian, B.E.; Stowe, R.P.; Pierson, D.L.; Sams, C.F. Immune system dysregulation following short- vs long-duration spaceflight. Aviat. Space Environ. Med. 2008, 79, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Fowler, B.; Meehan, S.; Singhal, A. Perceptual-motor performance and associated kinematics in space. Hum. Factors 2008, 50, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.B.; Hoover, M.; Southward, E.F. Cognitive performance during prismatic displacement as a partial analogue of “space fog”. Aviat. Space Environ. Med. 2009, 80, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Villasana, L.E.; Rosenthal, R.A.; Doctrow, S.R.; Pfankuch, T.; Zuloaga, D.G.; Garfinkel, A.M.; Raber, J. Effects of alpha-lipoic acid on associative and spatial memory of sham-irradiated and 56Fe-irradiated C57BL/6J male mice. Pharmacol. Biochem. Behav. 2013, 103, 487–493. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Haley, G.E.; Yeiser, L.; Olsen, R.H.; Davis, M.J.; Johnson, L.A.; Raber, J. Early effects of whole-body (56)Fe irradiation on hippocampal function in C57BL/6J mice. Radiat. Res. 2013, 179, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Maroso, M.; Syage, A.; Allen, B.D.; Angulo, M.C.; Soltesz, I.; Limoli, C.L. Persistent nature of alterations in cognition and neuronal circuit excitability after exposure to simulated cosmic radiation in mice. Exp. Neurol. 2018, 305, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Tidmore, A.; Dutta, S.M.; Fesshaye, A.S.; Russell, W.K.; Duncan, V.D.; Britten, R.A. Space Radiation-Induced Alterations in the Hippocampal Ubiquitin-Proteome System. Int. J. Mol. Sci. 2021, 22, 7713. [Google Scholar] [CrossRef]
- Parihar, V.K.; Allen, B.; Tran, K.K.; Macaraeg, T.G.; Chu, E.M.; Kwok, S.F.; Chmielewski, N.N.; Craver, B.M.; Baulch, J.E.; Acharya, M.M.; et al. What happens to your brain on the way to Mars. Sci. Adv. 2015, 1, e1400256. [Google Scholar] [CrossRef]
- Santucci, D.; Kawano, F.; Ohira, T.; Terada, M.; Nakai, N.; Francia, N.; Alleva, E.; Aloe, L.; Ochiai, T.; Cancedda, R.; et al. Evaluation of gene, protein and neurotrophin expression in the brain of mice exposed to space environment for 91 days. PLoS ONE 2012, 7, e40112. [Google Scholar] [CrossRef]
- Li, K.; Guo, X.; Jin, Z.; Ouyang, X.; Zeng, Y.; Feng, J.; Wang, Y.; Yao, L.; Ma, L. Effect of Simulated Microgravity on Human Brain Gray Matter and White Matter--Evidence from MRI. PLoS ONE 2015, 10, e0135835. [Google Scholar] [CrossRef]
- Mao, X.W.; Sandberg, L.B.; Gridley, D.S.; Herrmann, E.C.; Zhang, G.; Raghavan, R.; Zubarev, R.A.; Zhang, B.; Stodieck, L.S.; Ferguson, V.L.; et al. Proteomic Analysis of Mouse Brain Subjected to Spaceflight. Int. J. Mol. Sci. 2018, 20, 7. [Google Scholar] [CrossRef] [PubMed]
- Dow, R.S. Contribution of electrophysiological studies to cerebellar physiology. J. Clin. Neurophysiol. 1988, 5, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Cassady, K.; Koppelmans, V.; Reuter-Lorenz, P.; De Dios, Y.; Gadd, N.; Wood, S.; Castenada, R.R.; Kofman, I.; Bloomberg, J.; Mulavara, A.; et al. Effects of a spaceflight analog environment on brain connectivity and behavior. Neuroimage 2016, 141, 18–30. [Google Scholar] [CrossRef]
- Koppelmans, V.; Mulavara, A.P.; Seidler, R.D.; De Dios, Y.E.; Bloomberg, J.J.; Wood, S.J. Cortical thickness of primary motor and vestibular brain regions predicts recovery from fall and balance directly after spaceflight. Brain Struct. Funct. 2022, 227, 2073–2086. [Google Scholar] [CrossRef]
- Liao, Y.; Miao, D.; Huan, Y.; Yin, H.; Xi, Y.; Liu, X. Altered regional homogeneity with short-term simulated microgravity and its relationship with changed performance in mental transformation. PLoS ONE 2014, 8, e64931. [Google Scholar] [CrossRef] [PubMed]
- Kokhan, V.S.; Matveeva, M.I.; Bazyan, A.S.; Kudrin, V.S.; Mukhametov, A.; Shtemberg, A.S. Combined effects of antiorthostatic suspension and ionizing radiation on the behaviour and neurotransmitters changes in different brain structures of rats. Behav. Brain Res. 2017, 320, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Matveeva, M.I.; Shtemberg, A.S.; Timoshenko, G.N.; Krasavin, E.A.; Narkevich, V.B.; Klodt, P.M.; Kudrin, V.S.; Bazyan, A.S. The effects of irradiation by C-12 carbon ions on monoamine exchange in several rat brain structures. Neurochem. J. 2013, 7, 303–307. [Google Scholar] [CrossRef]
- Bhattacherjee, A.; Djekidel, M.N.; Chen, R.; Chen, W.; Tuesta, L.M.; Zhang, Y. Cell type-specific transcriptional programs in mouse prefrontal cortex during adolescence and addiction. Nat. Commun. 2019, 10, 4169. [Google Scholar] [CrossRef]
- de Hemptinne, C.; Chen, W.; Racine, C.A.; Seritan, A.L.; Miller, A.M.; Yaroshinsky, M.S.; Wang, S.S.; Gilron, R.; Little, S.; Bledsoe, I.; et al. Prefrontal Physiomarkers of Anxiety and Depression in Parkinson’s Disease. Front. Neurosci. 2021, 15, 748165. [Google Scholar] [CrossRef]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar] [CrossRef]
- Liang, R.; Wang, L.; Sun, S.; Zheng, C.; Yang, J.; Ming, D. Medial prefrontal cortex and hippocampus in mice differently affected by simulate microgravity and social isolation associated with the alternation of emotional and cognitive functions. Life Sci. Space Res. 2022, 33, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Sarkar, S.; Ramesh, V.; Hayes, B.E.; Thomas, R.L.; Wilson, B.L.; Kim, H.; Barnes, S.; Kulkarni, A.; Pellis, N.; et al. Proteomic analysis of mice hippocampus in simulated microgravity environment. J. Proteome Res. 2006, 5, 548–553. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Javed, I.; Liu, Y.; Lu, S.; Peng, G.; Zhang, Y.; Qing, H.; Deng, Y. Effect of Prolonged Simulated Microgravity on Metabolic Proteins in Rat Hippocampus: Steps toward Safe Space Travel. J. Proteome Res. 2016, 15, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Kiffer, F.; Alexander, T.; Anderson, J.E.; Groves, T.; Wang, J.; Sridharan, V.; Boerma, M.; Allen, A.R. Late Effects of (16)O-Particle Radiation on Female Social and Cognitive Behavior and Hippocampal Physiology. Radiat. Res. 2019, 191, 278–294. [Google Scholar] [CrossRef]
- Ohm, T.G. The dentate gyrus in Alzheimer’s disease. Prog. Brain Res. 2007, 163, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.P. Space flight and oxidative stress. Nutrition 2002, 18, 867–871. [Google Scholar] [CrossRef]
- Batinic-Haberle, I.; Reboucas, J.S.; Spasojevic, I. Superoxide dismutase mimics: Chemistry, pharmacology, and therapeutic potential. Antioxid. Redox Signal. 2010, 13, 877–918. [Google Scholar] [CrossRef]
- Weitzel, D.H.; Tovmasyan, A.; Ashcraft, K.A.; Rajic, Z.; Weitner, T.; Liu, C.; Li, W.; Buckley, A.F.; Prasad, M.R.; Young, K.H.; et al. Radioprotection of the brain white matter by Mn(III) n-Butoxyethylpyridylporphyrin-based superoxide dismutase mimic MnTnBuOE-2-PyP5+. Mol. Cancer Ther. 2015, 14, 70–79. [Google Scholar] [CrossRef]
- Mao, X.; Stanbouly, S.; Holley, J.; Pecaut, M.; Crapo, J. Evidence of Spaceflight-Induced Adverse Effects on Photoreceptors and Retinal Function in the Mouse Eye. Int. J. Mol. Sci. 2023, 24, 7362. [Google Scholar] [CrossRef]
- Williams, C.G.; Lee, H.J.; Asatsuma, T.; Vento-Tormo, R.; Haque, A. An introduction to spatial transcriptomics for biomedical research. Genome Med. 2022, 14, 68. [Google Scholar] [CrossRef]
- Stahl, P.L.; Salmen, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Zollinger, D.R.; Lingle, S.E.; Sorg, K.; Beechem, J.M.; Merritt, C.R. GeoMx RNA Assay: High Multiplex, Digital, Spatial Analysis of RNA in FFPE Tissue. Methods Mol. Biol. 2020, 2148, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Christian, K.M.; Song, H.; Ming, G.L. Functions and dysfunctions of adult hippocampal neurogenesis. Ann. Rev. Neurosci. 2014, 37, 243–262. [Google Scholar] [CrossRef] [PubMed]
- McNerlin, C.; Guan, F.; Bronk, L.; Lei, K.; Grosshans, D.; Young, D.W.; Gaber, M.W.; Maletic-Savatic, M. Targeting hippocampal neurogenesis to protect astronauts’ cognition and mood from decline due to space radiation effects. Life Sci. Space Res. 2022, 35, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Curtis, S.B.; Vazquez, M.E.; Wilson, J.W.; Atwell, W.; Kim, M.; Capala, J. Cosmic ray hit frequencies in critical sites in the central nervous system. Adv. Space Res. 1998, 22, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, J.T.; Schafer, S.T.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 2016, 167, 897–914. [Google Scholar] [CrossRef]
- Bond, A.M.; Ming, G.L.; Song, H. Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later. Cell Stem Cell 2015, 17, 385–395. [Google Scholar] [CrossRef]
- Park, A.J.; Harris, A.Z.; Martyniuk, K.M.; Chang, C.Y.; Abbas, A.I.; Lowes, D.C.; Kellendonk, C.; Gogos, J.A.; Gordon, J.A. Reset of hippocampal-prefrontal circuitry facilitates learning. Nature 2021, 591, 615–619. [Google Scholar] [CrossRef]
- Liu, B.; Hinshaw, R.G.; Le, K.X.; Park, M.A.; Wang, S.; Belanger, A.P.; Dubey, S.; Frost, J.L.; Shi, Q.; Holton, P.; et al. Space-like (56)Fe irradiation manifests mild, early sex-specific behavioral and neuropathological changes in wildtype and Alzheimer’s-like transgenic mice. Sci. Rep. 2019, 9, 12118. [Google Scholar] [CrossRef]
- Raber, J.; Rudobeck, E.; Campbell-Beachler, M.; Allen, A.R.; Allen, B.; Rosi, S.; Nelson, G.A.; Ramachandran, S.; Turner, J.; Fike, J.R.; et al. 28Silicon radiation-induced enhancement of synaptic plasticity in the hippocampus of naive and cognitively tested mice. Radiat. Res. 2014, 181, 362–368. [Google Scholar] [CrossRef]
- Rosenstein, J.M.; Krum, J.M.; Ruhrberg, C. VEGF in the nervous system. Organogenesis 2010, 6, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Ermak, G.; Davies, K.J. Calcium and oxidative stress: From cell signaling to cell death. Mol. Immunol. 2002, 38, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Storz, P. Forkhead Homeobox Type O Transcription Factors in the Responses to Oxidative Stress. Antioxid. Redox Signal. 2011, 14, 593–605. [Google Scholar] [CrossRef]
- Martins, S.G.; Zilhao, R.; Thorsteinsdottir, S.; Carlos, A.R. Linking Oxidative Stress and DNA Damage to Changes in the Expression of Extracellular Matrix Components. Front. Genet. 2021, 12, 673002. [Google Scholar] [CrossRef] [PubMed]
- Larina, I.M.; Nichiporuk, I.A.; Veselova, O.M.; Vasilieva, G.Y.; Popova, I.A. Shifts in metabolism and its regulation under the effect of spaceflight factors. Aviakosm. Ekolog. Med. 2013, 47, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Jonscher, K.R.; Alfonso-Garcia, A.; Suhalim, J.L.; Orlicky, D.J.; Potma, E.O.; Ferguson, V.L.; Bouxsein, M.L.; Bateman, T.A.; Stodieck, L.S.; Levi, M.; et al. Spaceflight Activates Lipotoxic Pathways in Mouse Liver. PLoS ONE 2016, 11, e0152877. [Google Scholar] [CrossRef]
- Uruno, A.; Saigusa, D.; Suzuki, T.; Yumoto, A.; Nakamura, T.; Matsukawa, N.; Yamazaki, T.; Saito, R.; Taguchi, K.; Suzuki, M.; et al. Nrf2 plays a critical role in the metabolic response during and after spaceflight. Commun. Biol. 2021, 4, 1381. [Google Scholar] [CrossRef]
- Stroud, J.E.; Gale, M.S.; Zwart, S.R.; Heer, M.; Smith, S.M.; Montina, T.; Metz, G.A.S. Longitudinal metabolomic profiles reveal sex-specific adjustments to long-duration spaceflight and return to Earth. Cell Mol. Life Sci. 2022, 79, 578. [Google Scholar] [CrossRef]
- Overbey, E.G.; da Silveira, W.A.; Stanbouly, S.; Nishiyama, N.C.; Roque-Torres, G.D.; Pecaut, M.J.; Zawieja, D.C.; Wang, C.; Willey, J.S.; Delp, M.D.; et al. Spaceflight influences gene expression, photoreceptor integrity, and oxidative stress-related damage in the murine retina. Sci. Rep. 2019, 9, 13304. [Google Scholar] [CrossRef]
- Brainard, G.C.; Barger, L.K.; Soler, R.R.; Hanifin, J.P. The development of lighting countermeasures for sleep disruption and circadian misalignment during spaceflight. Curr. Opin. Pulm. Med. 2016, 22, 535–544. [Google Scholar] [CrossRef]
- Gundel, A.; Polyakov, V.V.; Zulley, J. The alteration of human sleep and circadian rhythms during spaceflight. J. Sleep Res. 1997, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Iyer, J.; Mhatre, S.D.; Gilbert, R.; Bhattacharya, S. Multi-system responses to altered gravity and spaceflight: Insights from Drosophila melanogaster. Neurosci. Biobehav. Rev. 2022, 142, 104880. [Google Scholar] [CrossRef] [PubMed]
- Mallis, M.M.; DeRoshia, C.W. Circadian rhythms, sleep, and performance in space. Aviat. Space Environ. Med. 2005, 76, B94–B107. [Google Scholar] [PubMed]
- Flynn-Evans, E.E.; Barger, L.K.; Kubey, A.A.; Sullivan, J.P.; Czeisler, C.A. Circadian misalignment affects sleep and medication use before and during spaceflight. NPJ Microgravity 2016, 2, 15019. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.A.; Kent, B.A.; Grant, L.K.; Clark, T.; Hanifin, J.P.; Barger, L.K.; Czeisler, C.A.; Brainard, G.C.; St Hilaire, M.A.; Lockley, S.W. Effects of dynamic lighting on circadian phase, self-reported sleep and performance during a 45-day space analog mission with chronic variable sleep deficiency. J. Pineal Res. 2022, 73, e12826. [Google Scholar] [CrossRef] [PubMed]
- Shephard, A.M.; Brown, N.S.; Snell-Rood, E.C. Anthropogenic Zinc Exposure Increases Mortality and Antioxidant Gene Expression in Monarch Butterflies with Low Access to Dietary Macronutrients. Environ. Toxicol. Chem. 2022, 41, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C.; Sullivan, D.W., Jr.; Spasojevic, I.; Mujer, C.V.; Spainhour, C.B.; Crapo, J.D. Nonclinical Safety and Toxicokinetics of MnTnBuOE-2-PyP5+ (BMX-001). Int. J. Toxicol. 2016, 35, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Batinic-Haberle, I.; Tovmasyan, A.; Roberts, E.R.; Vujaskovic, Z.; Leong, K.W.; Spasojevic, I. SOD therapeutics: Latest insights into their structure-activity relationships and impact on the cellular redox-based signaling pathways. Antioxid. Redox Signal. 2014, 20, 2372–2415. [Google Scholar] [CrossRef]
- Prokop, S.; Miller, K.R.; Labra, S.R.; Pitkin, R.M.; Hoxha, K.; Narasimhan, S.; Changolkar, L.; Rosenbloom, A.; Lee, V.M.; Trojanowski, J.Q. Impact of TREM2 risk variants on brain region-specific immune activation and plaque microenvironment in Alzheimer’s disease patient brain samples. Acta Neuropathol. 2019, 138, 613–630. [Google Scholar] [CrossRef]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef]
- Beechem, J.M. High-Plex Spatially Resolved RNA and Protein Detection Using Digital Spatial Profiling: A Technology Designed for Immuno-oncology Biomarker Discovery and Translational Research. Methods Mol. Biol. 2020, 2055, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Pinero, J.; Bravo, A.; Queralt-Rosinach, N.; Gutierrez-Sacristan, A.; Deu-Pons, J.; Centeno, E.; Garcia-Garcia, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Chambers, J.M.; Hastie, T. Analysis of Variance; Designed Experiments. In Statistical Models in S; Wadsworth & Brooks/Cole Advanced Books & Software: Pacific Grove, CA, USA, 1992. [Google Scholar]
- Storey, J.D. The positive false discovery rate: A Bayesian interpretation and the q-value. Ann. Stat. 2003, 31, 2013–2035. [Google Scholar] [CrossRef]
- Husson, F.O.; Lê, S.B.; Pageès, J.R.M. Exploratory Multivariate Analysis by Example Using R; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremsky, I.; Ali, S.; Stanbouly, S.; Holley, J.; Justinen, S.; Pecaut, M.; Crapo, J.; Mao, X. Spaceflight-Induced Gene Expression Profiles in the Mouse Brain Are Attenuated by Treatment with the Antioxidant BuOE. Int. J. Mol. Sci. 2023, 24, 13569. https://doi.org/10.3390/ijms241713569
Kremsky I, Ali S, Stanbouly S, Holley J, Justinen S, Pecaut M, Crapo J, Mao X. Spaceflight-Induced Gene Expression Profiles in the Mouse Brain Are Attenuated by Treatment with the Antioxidant BuOE. International Journal of Molecular Sciences. 2023; 24(17):13569. https://doi.org/10.3390/ijms241713569
Chicago/Turabian StyleKremsky, Isaac, Samir Ali, Seta Stanbouly, Jacob Holley, Stephen Justinen, Michael Pecaut, James Crapo, and Xiaowen Mao. 2023. "Spaceflight-Induced Gene Expression Profiles in the Mouse Brain Are Attenuated by Treatment with the Antioxidant BuOE" International Journal of Molecular Sciences 24, no. 17: 13569. https://doi.org/10.3390/ijms241713569
APA StyleKremsky, I., Ali, S., Stanbouly, S., Holley, J., Justinen, S., Pecaut, M., Crapo, J., & Mao, X. (2023). Spaceflight-Induced Gene Expression Profiles in the Mouse Brain Are Attenuated by Treatment with the Antioxidant BuOE. International Journal of Molecular Sciences, 24(17), 13569. https://doi.org/10.3390/ijms241713569