
The SbbHLH041–SbEXPA11 Module Enhances Cadmium Accumulation and Rescues Biomass by Increasing Photosynthetic Efficiency in Sorghum


 and
and 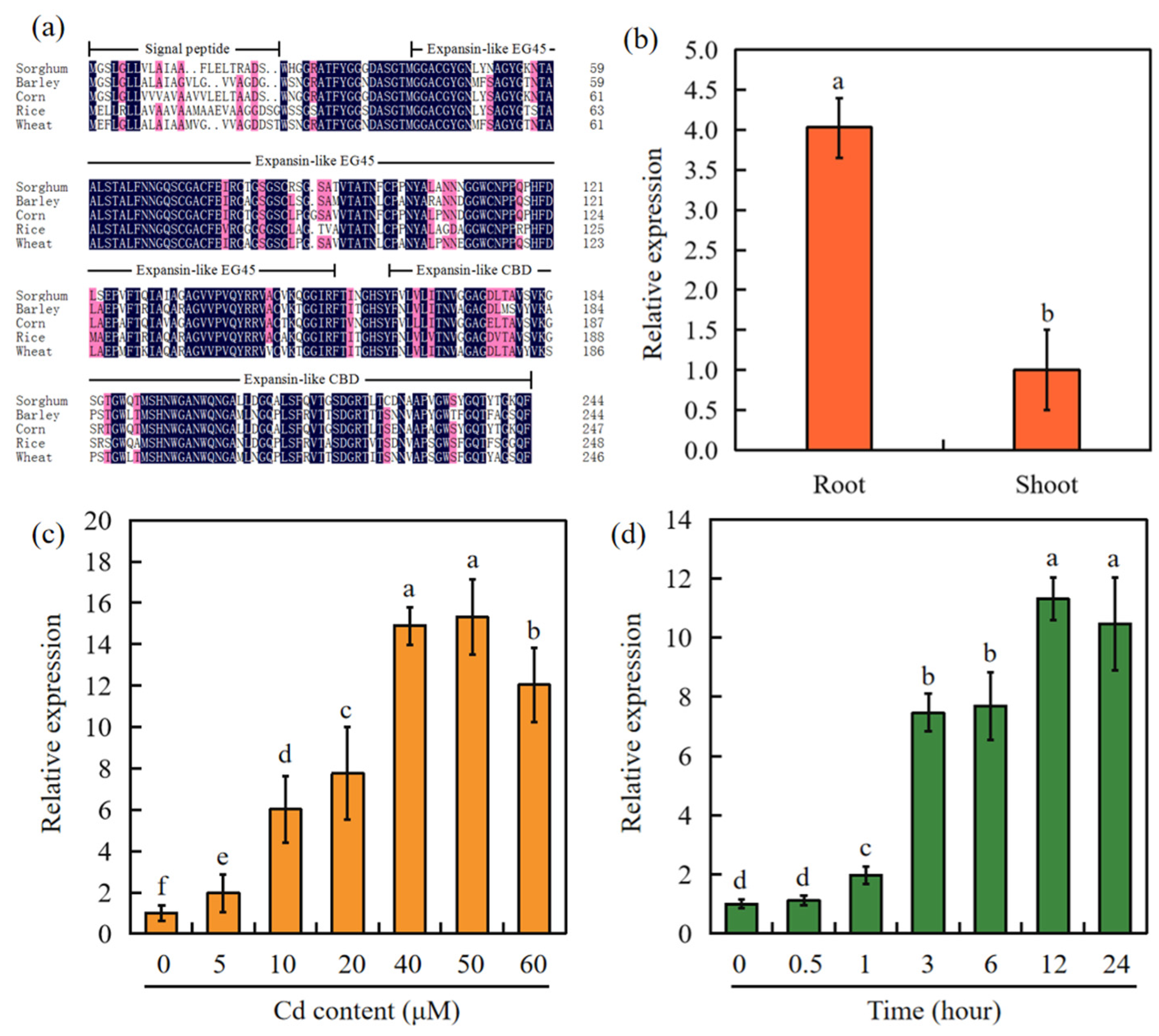{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cloning and Expression of SbEXPA11 in Sorghum
2.2. The SbbHLH041 TF Upregulates SbEXPA11 Expression by Binding Directly to Its Promoter
2.3. Generation and Identification of SbEXPA11-Overexpressing Sorghum Lines
2.4. Overexpression of SbEXPA11 Increases Cd Uptake and Long-Distance Transport
2.5. Overexpression of SbEXPA11 Reduces Cd Damage in Transgenic Sorghum
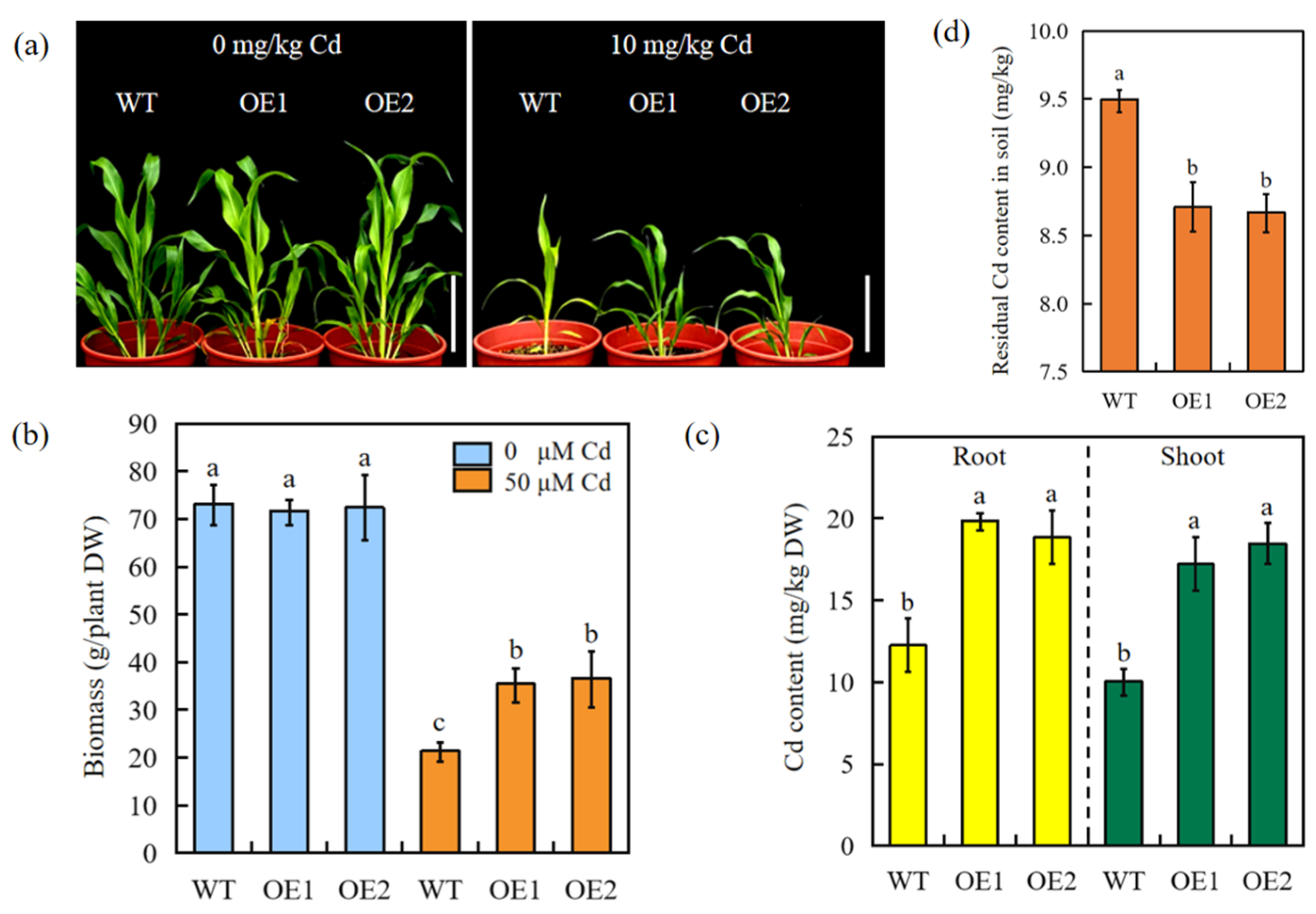
2.6. Overexpression of SbEXPA11 Increases Phytoremediation Capacity
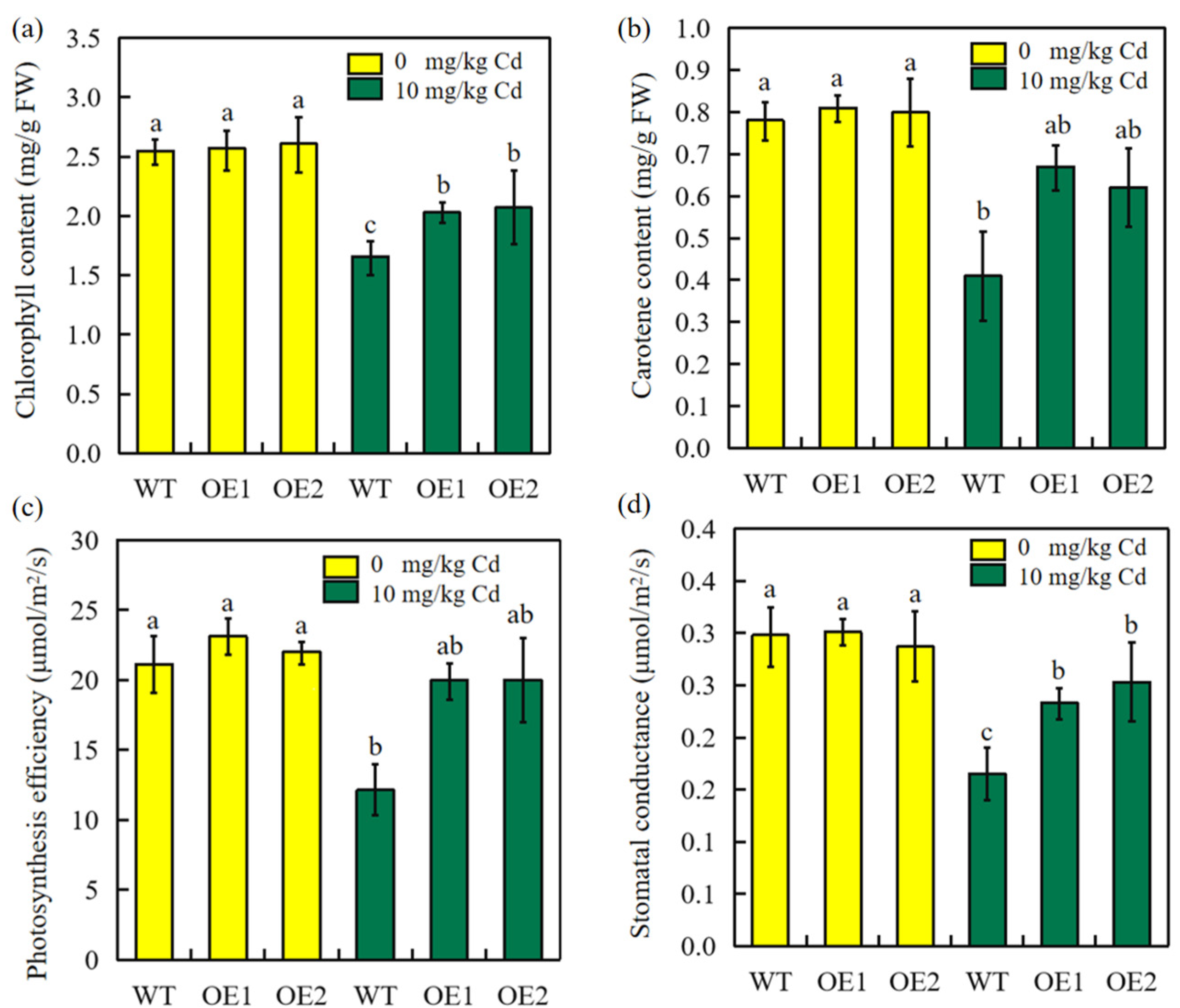
2.7. Overexpression of SbEXPA11 Rescues Photosynthetic Pigments and Efficiency, Stomatal Conductance, and Biomass Production
3. Discussion
3.1. The SbbHLH041 TF Activates SbEXPA11 Expression by Directly Binding to the E-Box in Its Promoter
3.2. Overexpression of SbEXPA11 Increases Cd Accumulation by Increasing Uptake and Long-Distance Transport
3.3. SbEXPA11 Mediates Tolerance to Cd by Improving ROS Scavenging
3.4. Overexpression of SbEXPA11 Rescues Biomass Production by Increasing Photosynthetic Efficiency
3.5. Perspective
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. RNA Extraction and RT–qPCR
4.3. Characterization of SbEXPA11 in Sorghum
4.4. Genetic Transformation
4.5. Measurement of Cd, MDA, and Proline Content, Electrolyte Leakage, ROS Content, and Antioxidant Enzyme Activities
4.6. Measurement of Chlorophyll and Carotene Content
4.7. Measurement of Photosynthetic Efficiency, Stomatal Conductance, and Biomass Production
4.8. Yeast One-Hybrid (Y1H) Assay
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Statistical Analyses
4.11. Primers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muslu, T.; Akpinar, B.A.; Biyiklioglu-Kaya, S.; Yuce, M.; Budak, H. Comparative analysis of coding and non-coding features within insect tolerance loci in wheat with their homologs in cereal genomes. Int. J. Mol. Sci. 2021, 16, 12349. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, N.C.; Bean, E.; Miles, A.M.; de Carvalho, D.U.O.G.; Alemán, M.A.R.; Carvalho, J.S.; Gregory, L.; Ganda, E.A. Cross-sectional study of dairy cattle metagenomes reveals increased antimicrobial resistance in animals farmed in a heavy metal contaminated environment. Front. Microbiol. 2020, 11, 590325. [Google Scholar] [CrossRef] [PubMed]
- Asim, S.; Hussain, A.; Murad, W.; Hamayun, M.; Iqbal, A.; Rehman, H.; Tawab, A.; Irshad, M.; Alataway, A.; Dewidar, A.Z.; et al. Endophytic Fusarium oxysporum GW controlling weed and an effective biostimulant for wheat growth. Front. Plant Sci. 2022, 13, 922343. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Rajpoot, R.; Pandey, P.; Rani, A.; Dubey, R.S. Cadmium alters mitochondrial membrane potential, inhibits electron transport chain activity and induces callose deposition in rice seedlings. J. Plant Growth Regul. 2018, 37, 335–344. [Google Scholar] [CrossRef]
- Rogowska, A.; Pączkowski, C.; Szakiel, A. Modulation of steroid and triterpenoid metabolism in Calendula officinalis plants and hairy root cultures exposed to cadmium stress. Int. J. Mol. Sci. 2022, 23, 5640. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Lampis, S.; Santi, C.; Ciurli, A.; Andreolli, M.; Vallini, G. Promotion of arsenic phytoextraction efficiency in the fern Pteris vittata by the inoculation of As-resistant bacteria: A soil bioremediation perspective. Front. Plant Sci. 2015, 6, 80. [Google Scholar] [CrossRef]
- Laloum, D.; Magen, S.; Soroka, Y.; Avin-Wittenberg, T. Exploring the contribution of autophagy to the excess-sucrose response in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 3891. [Google Scholar] [CrossRef]
- Noh, S.A.; Lee, H.S.; Kim, Y.S.; Paek, K.H.; Shin, J.S.; Bae, J.M. Down–regulation of the IbEXP1 gene enhanced storage root development in sweet potato. J. Exp. Bot. 2013, 64, 129–142. [Google Scholar] [CrossRef]
- Cho, H.T.; Cosgrove, D.J. Regulation of root hair initiation and expansin gene expression in Arabidopsis. Plant Cell 2002, 14, 3237–3253. [Google Scholar] [CrossRef]
- Kwasniewski, M.; Szarejko, I. Molecular cloning and characterization of β–expansin gene related to root hair formation in barley. Plant Physiol. 2006, 141, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zeng, J.; Cao, F.; Ahmed, I.M.; Zhang, G.; Vincze, E.; Wu, F. HvEXPB7, a novel β–expansin gene revealed by the root hair transcriptome of Tibetan wild barley, improves root hair growth under drought stress. J. Exp. Bot. 2015, 66, 7405–7419. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ren, Y.; Zhang, G.; An, J.; Yang, J.; Wang, Y.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improves oxidative stress tolerance in transgenic Arabidopsis plants. Plant Physiol. Biochem. 2018, 124, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, H.; Yang, R.; Xu, X.; Liu, X.; Xu, J. Over–expression of PttEXPA8 gene showed various resistances to diverse stresses. Int. J. Biol. Macromol. 2019, 130, 50–57. [Google Scholar] [CrossRef]
- Zhang, H.; Ding, Y.; Zhi, J.; Li, X.; Liu, H.; Xu, J. Over–expression of the poplar expansin gene PtoEXPA12 in tobacco plants enhanced cadmium accumulation. Int. J. Biol. Macromol. 2018, 116, 676–682. [Google Scholar] [CrossRef]
- Ren, Y.; Chen, Y.; An, J.; Zhao, Z.; Zhang, G.; Wang, Y.; Wang, W. Wheat expansin gene TaEXPA2 is involved in conferring plant tolerance to Cd toxicity. Plant Sci. 2018, 270, 245–256. [Google Scholar] [CrossRef]
- Khan, I.; Asaf, S.; Jan, R.; Bilal, S.; Khan, A.L.; Kim, K.M.; Al–Harrasi, A. Genome–wide annotation and expression analysis of WRKY and bHLH transcriptional factor families reveal their involvement under cadmium stress in tomato (Solanum lycopersicum L.). Front. Plant Sci. 2023, 14, 1100895. [Google Scholar] [CrossRef]
- Verma, S.; Gazara, R.K.; Verma, P.K. Transcription factor repertoire of necrotrophic fungal phytopathogen Ascochyta rabiei: Predominance of MYB transcription factors as potential regulators of secretome. Front. Plant Sci. 2017, 8, 1037. [Google Scholar] [CrossRef]
- Jung, S.E.; Bang, S.W.; Kim, S.H.; Seo, J.S.; Yoon, H.B.; Kim, Y.S.; Kim, J.K. Overexpression of OsERF83, a vascular tissue-specific transcription factor gene, confers drought tolerance in rice. Int. J. Mol. Sci. 2021, 22, 7656. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Fukuoka, H.; Arao, T.; Ohyama, A.; Nunome, T.; Miyatake, K.; Negoro, S. Gene expression analysis in cadmium-stressed roots of a low cadmium-accumulating solanaceous plant, Solanum torvum. J. Exp. Bot. 2010, 61, 423–437. [Google Scholar] [CrossRef]
- Sapara, K.K.; Khedia, J.; Agarwal, P.; Gangapur, D.R.; Agarwal, P.K. SbMYB15 transcription factor mitigates cadmium and nickel stress in transgenic tobacco by limiting uptake and modulating antioxidative defence system. Funct. Plant Biol. 2019, 46, 702–714. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; Tang, H.; Du, X.; He, F.; Ren, M.; Bao, Y. TaSWEET14 confers low cadmium accumulation in wheat and is regulated by TaMYB41. Environ. Exp. Bot. 2022, 201, 104992. [Google Scholar] [CrossRef]
- Jia, Z.; Li, M.; Wang, H.; Zhu, B.; Gu, L.; Du, X.; Ren, M. TaWRKY70 positively regulates TaCAT5 enhanced Cd tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2021, 190, 104591. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; He, F.; Du, X.; Ren, M.; Bao, Y. The TaWRKY22–TaCOPT3D pathway governs cadmium uptake in wheat. Int. J. Mol. Sci. 2022, 23, 10379. [Google Scholar] [CrossRef]
- Wang, H.; Liao, S.; Li, M.; Wei, J.; Zhu, B.; Gu, L.; Li, L.H.; Du, X. TmNAS3 from Triticum monococum directly regulated by TmbHLH47 increases Fe content of wheat grain. Gene 2022, 811, 146096. [Google Scholar] [CrossRef]
- Du, X.; He, F.; Zhu, B.; Ren, M.; Tang, H. NAC transcription factors from Aegilops markgrafii reduce cadmium concentration in transgenic wheat. Plant Soil 2020, 449, 39–50. [Google Scholar] [CrossRef]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant Physiol. 2008, 146, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kim, H.E.; Kim, S. Two different domain architectures generate structural and functional diversity among bZIP genes in the Solanaceae family. Front. Plant Sci. 2022, 13, 967546. [Google Scholar] [CrossRef] [PubMed]
- Dellagi, A.; Segond, D.; Rigault, M.; Fagard, M.; Simon, C.; Saindrenan, P.; Expert, D. Microbial siderophores exert a subtle role in Arabidopsis during infection by manipulating the immune response and the iron status. Plant Physiol. 2009, 150, 1687–1696. [Google Scholar] [CrossRef]
- Long, T.A.; Tsukagoshi, H.; Busch, W.; Lahner, B.; Salt, D.E.; Benfey, P.N. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell 2010, 22, 2219–2236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.S.P.; Xu, J. The R2R3–MYB transcription factor MYB49 regulates cadmium accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.L.; Wang, Y.Y.; Xia, L.Z. The maize bHLH transcription factor bHLH105 confers manganese tolerance in transgenic tobacco. Plant Sci. 2019, 280, 97–109. [Google Scholar] [CrossRef]
- Aono, A.H.; Pimenta, R.J.G.; Garcia, A.L.B.; Correr, F.H.; Hosaka, G.K.; Carrasco, M.M.; Cardoso-Silva, C.B.; Mancini, M.C.; Sforça, D.A.; dos Santos, L.B.; et al. The wild sugarcane and sorghum kinomes: Insights into expansion, diversification, and expression patterns. Front. Plant Sci. 2021, 12, 668623. [Google Scholar] [CrossRef] [PubMed]
- Carretero–Paulet, L.; Galstyan, A.; Roig–Villanova, I.; Martínez–García, J.F.; Bilbao–Castro, J.R.; Robertson, D.L. Genome–wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis, poplar, rice, moss, and algae. Plant Physiol. 2010, 153, 1398–1412. [Google Scholar] [CrossRef]
- Strygina, K.V.; Khlestkina, E.K. Myc–like transcriptional factors in wheat: Structural and functional organization of the subfamily I members. BMC Plant Biol. 2019, 19, 50. [Google Scholar] [CrossRef]
- Wang, H.; Zuo, D.; Zhu, B.; Du, X.; Gu, L. TaMYC8 regulates TaERF6 and inhibits ethylene synthesis to confer Cd tolerance in wheat. Environ. Exp. Bot. 2022, 198, 104854. [Google Scholar] [CrossRef]
- Głazowska, S.; Baldwin, L.; Mravec, J.; Bukh, C.; Hansen, T.H.; Jensen, M.M.; Fangel, J.U.; Willats, W.G.T.; Glasius, M.; Felby, C.; et al. The impact of silicon on cell wall composition and enzymatic saccharification of Brachypodium distachyon. Biotechnol. Biofuels. 2018, 11, 171. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Rahman, M.A.; Kabir, A.H.; Mandal, A.; Roy, S.K.; Song, Y.; Ji, H.C.; Lee, K.W. Glutathione restores Hg-induced morpho-physiological retardations by inducing phytochelatin and oxidative defense in Alfalfa. Biology 2020, 28, 364. [Google Scholar] [CrossRef]
- Massarioli, A.P.; Sartori, A.G.O.; Juliano, F.F.; Santos, R.C.D.; Ramos, J.P.C.; Lima, L.M.; Alencar, S.M. Optimizing procedures for antioxidant phenolics extraction from skin and kernel of peanuts with contrasting levels of drought tolerance foods. Foods 2022, 11, 449. [Google Scholar] [CrossRef]
- Küpper, H.; Parameswaran, A.; Leitenmaier, B.; Trtílek, M.; Šetlík, I. Cadmium–induced inhibition of photosynthesis and long–term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol. 2007, 175, 655–674. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of cadmium stress on leaf chlorophyll fluorescence and photosynthesis of Elsholtzia argyi–a cadmium accumulating plant. Int. J. Phytoremediation 2015, 17, 85–92. [Google Scholar] [CrossRef]
- Sorrentino, M.C.; Capozzi, F.; Amitrano, C.; De Tommaso, G.; Arena, C.; Iuliano, M.; Giordano, S.; Spagnuolo, V. Facing metal stress by multiple strategies: Morphophysiological responses of cardoon (Cynara cardunculus L.) grown in hydroponics. Environ. Sci. Pollut. Res. 2021, 28, 37616–37626. [Google Scholar] [CrossRef] [PubMed]
- Benyó, D.; Horváth, E.; Németh, E.; Leviczky, T.; Takács, K.; Lehotai, N.; Feigl, G.; Kolbert, Z.; Ördög, A.; Gallé, R.; et al. Physiological and molecular responses to heavy metal stresses suggest different detoxification mechanism of Populus deltoides and P. x canadensis. J. Plant Physiol. 2016, 201, 62–70. [Google Scholar] [CrossRef]
- Paape, T.; Heiniger, B.; Santo Domingo, M.; Clear, M.R.; Lucas, M.M.; Pueyo, J.J. Genome-wide association study reveals complex genetic architecture of cadmium and mercury accumulation and tolerance traits in Medicago truncatula. Front. Plant Sci. 2022, 12, 806949. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Jhanji, S. Effect of soil cadmium on growth, photosynthesis and quality of Raphanus sativus and Lactuca sativa. J. Environ. Biol. 2016, 37, 993–997. [Google Scholar] [PubMed]
- Migocka, M.; Papierniak, A.; Kosatka, E.; Klobus, G. Comparative study of the active cadmium efflux systems operating at the plasma membrane and tonoplast of cucumber root cells. J. Exp. Bot. 2011, 62, 4903–4916. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Kondo, E.; Sugiura, D.; Terashima, I.; Suzuki, Y.; Makino, A. Enhanced leaf photosynthesis as a target to increase grain yield: Insights from transgenic rice lines with variable Rieske Fes protein content in the cytochrome b6/f complex. Plant Cell Environ. 2016, 39, 80–87. [Google Scholar] [CrossRef]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Fujikura, U.; Olas, J.J.; Xue, G.P.; Balazadeh, S.; Mueller-Roeber, B. GROWTH-REGULATING FACTOR 9 negatively regulates Arabidopsis leaf growth by controlling ORG3 and restricting cell proliferation in leaf primordia. PLoS Genet. 2018, 14, e1007484. [Google Scholar] [CrossRef]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix–loop–helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real–time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Che, P.; Anand, A.; Wu, E.; Sander, J.D.; Simon, M.K.; Zhu, W.; Sigmund, A.; Zastrow–Hayes, G.; Miller, M.; Liu, D.; et al. Developing a flexible, high-efficiency Agrobacterium–mediated sorghum transformation system with broad application. Plant Biotech. J. 2018, 16, 1388–1395. [Google Scholar] [CrossRef] [PubMed]
- Ror, S.; Panwar, S.L. Sef1-regulated iron regulon responds to mitochondria-dependent iron-sulfur cluster biosynthesis in Candida albicans. Front. Microbiol. 2019, 10, 1528. [Google Scholar] [CrossRef] [PubMed]
- Perinban, S.; Orsat, V.; Lyew, D.; Raghavan, V. Effect of plasma activated water on Escherichia coli disinfection and quality of kale and spinach. Food Chem. 2022, 397, 133793. [Google Scholar] [CrossRef]
- Stefanov, M.A.; Rashkov, G.D.; Yotsova, E.K.; Borisova, P.B.; Dobrikova, A.G.; Apostolova, E.L. Different sensitivity levels of the photosynthetic apparatus in Zea mays L. and Sorghum bicolor L. under salt stress. Plants 2021, 10, 1469. [Google Scholar] [CrossRef]
- Wilkins, O.; Hafemeister, C.; Plessis, A.; Holloway-Phillips, M.M.; Pham, G.M.; Nicotra, A.B.; Gregorio, G.B.; Jagadish, S.V.; Septiningsih, E.M.; Bonneau, R.; et al. EGRINs (Environmental Gene Regulatory Influence Networks) in rice that function in the response to water Deficit, high temperature, and agricultural environments. Plant Cell. 2016, 28, 2365–2384. [Google Scholar] [CrossRef]
- Cheong, M.S.; Kirik, A.; Kim, J.G.; Frame, K.; Kirik, V.; Mudgett, M.B. AvrBsT acetylates Arabidopsis ACIP1, a protein that associates with microtubules and is required for immunity. PLoS Pathog. 2014, 10, e1003952. [Google Scholar] [CrossRef]
- Azeez, A.; Zhao, Y.C.; Singh, R.K.; Yordanov, Y.S.; Dash, M.; Miskolczi, P.; Stojkovič, K.; Strauss, S.H.; Bhalerao, R.P.; Busov, V.B. EARLY BUD-BREAK 1 and EARLY BUD-BREAK 3 control resumption of poplar growth after winter dormancy. Nat. Commun. 2021, 18, 1123. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Yu, J.; Zhu, B.; Gu, L.; Wang, H.; Du, X.; Zeng, T.; Tang, H. The SbbHLH041–SbEXPA11 Module Enhances Cadmium Accumulation and Rescues Biomass by Increasing Photosynthetic Efficiency in Sorghum. Int. J. Mol. Sci. 2023, 24, 13061. https://doi.org/10.3390/ijms241713061
Wang H, Yu J, Zhu B, Gu L, Wang H, Du X, Zeng T, Tang H. The SbbHLH041–SbEXPA11 Module Enhances Cadmium Accumulation and Rescues Biomass by Increasing Photosynthetic Efficiency in Sorghum. International Journal of Molecular Sciences. 2023; 24(17):13061. https://doi.org/10.3390/ijms241713061
Chicago/Turabian StyleWang, Huinan, Junxing Yu, Bin Zhu, Lei Gu, Hongcheng Wang, Xuye Du, Tuo Zeng, and Heng Tang. 2023. "The SbbHLH041–SbEXPA11 Module Enhances Cadmium Accumulation and Rescues Biomass by Increasing Photosynthetic Efficiency in Sorghum" International Journal of Molecular Sciences 24, no. 17: 13061. https://doi.org/10.3390/ijms241713061
APA StyleWang, H., Yu, J., Zhu, B., Gu, L., Wang, H., Du, X., Zeng, T., & Tang, H. (2023). The SbbHLH041–SbEXPA11 Module Enhances Cadmium Accumulation and Rescues Biomass by Increasing Photosynthetic Efficiency in Sorghum. International Journal of Molecular Sciences, 24(17), 13061. https://doi.org/10.3390/ijms241713061