
Clinical Parameters and Epigenetic Biomarkers of Plaque Vulnerability in Patients with Carotid Stenosis

 , , and
, , and
Abstract
:1. Introduction
Carotid Stenosis: Relevant but Not Sufficient to Identify Vulnerable Plaques
2. Clinical, Haemodynamic and Neuroimaging Parameters for Identifying Stroke Risk Subgroups
2.1. Clinical Considerations in Risk Stratification
2.2. Haemodynamic Considerations in Risk Stratification
2.3. High Risk Features of Atheromatous Plaque in Neuroimaging
3. Epigenetic Biomarkers
3.1. DNA Methylation
3.2. Histone Post-Translational Modifications
3.3. Non-Coding RNAs
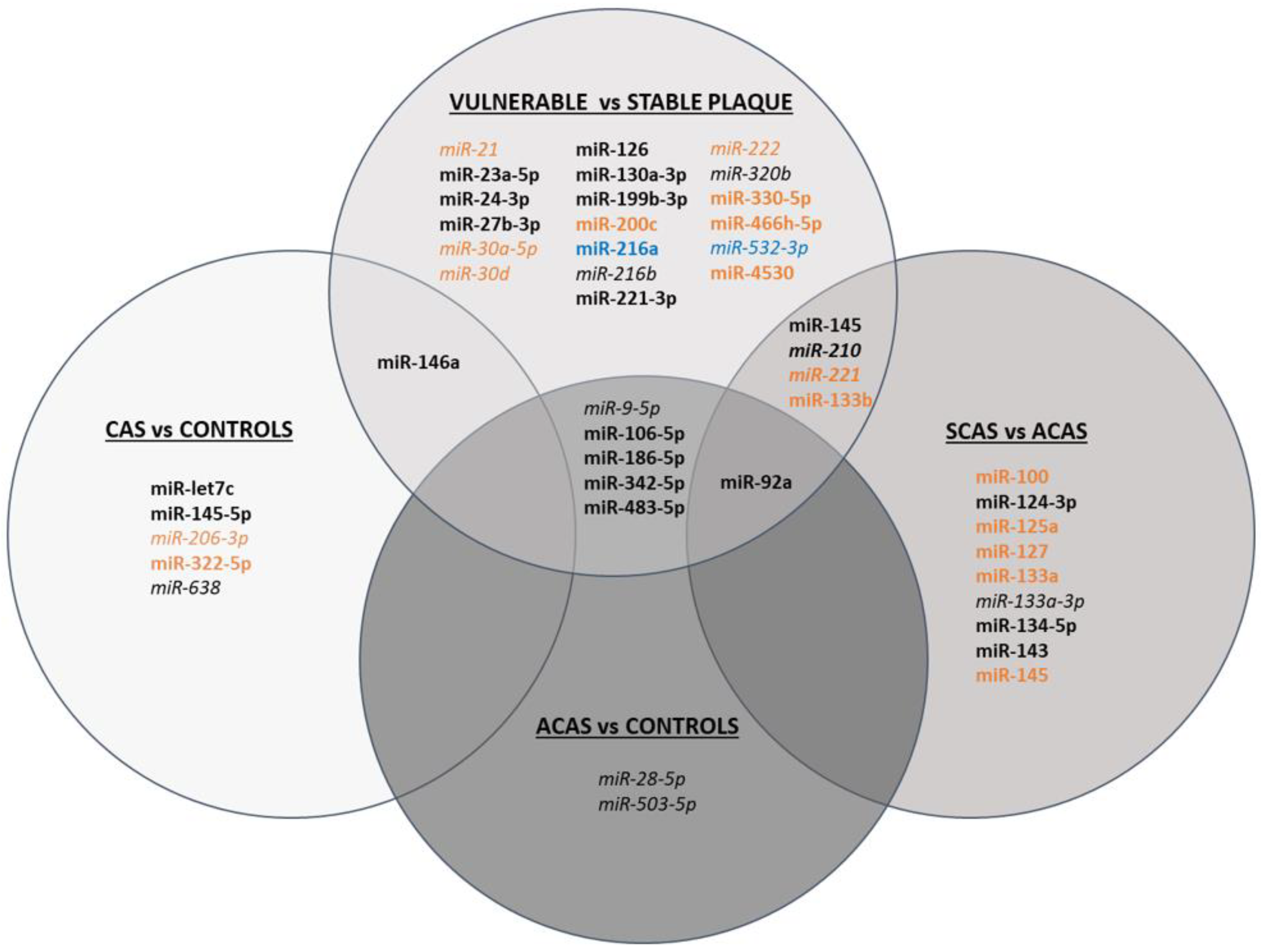
3.3.1. miRNAs
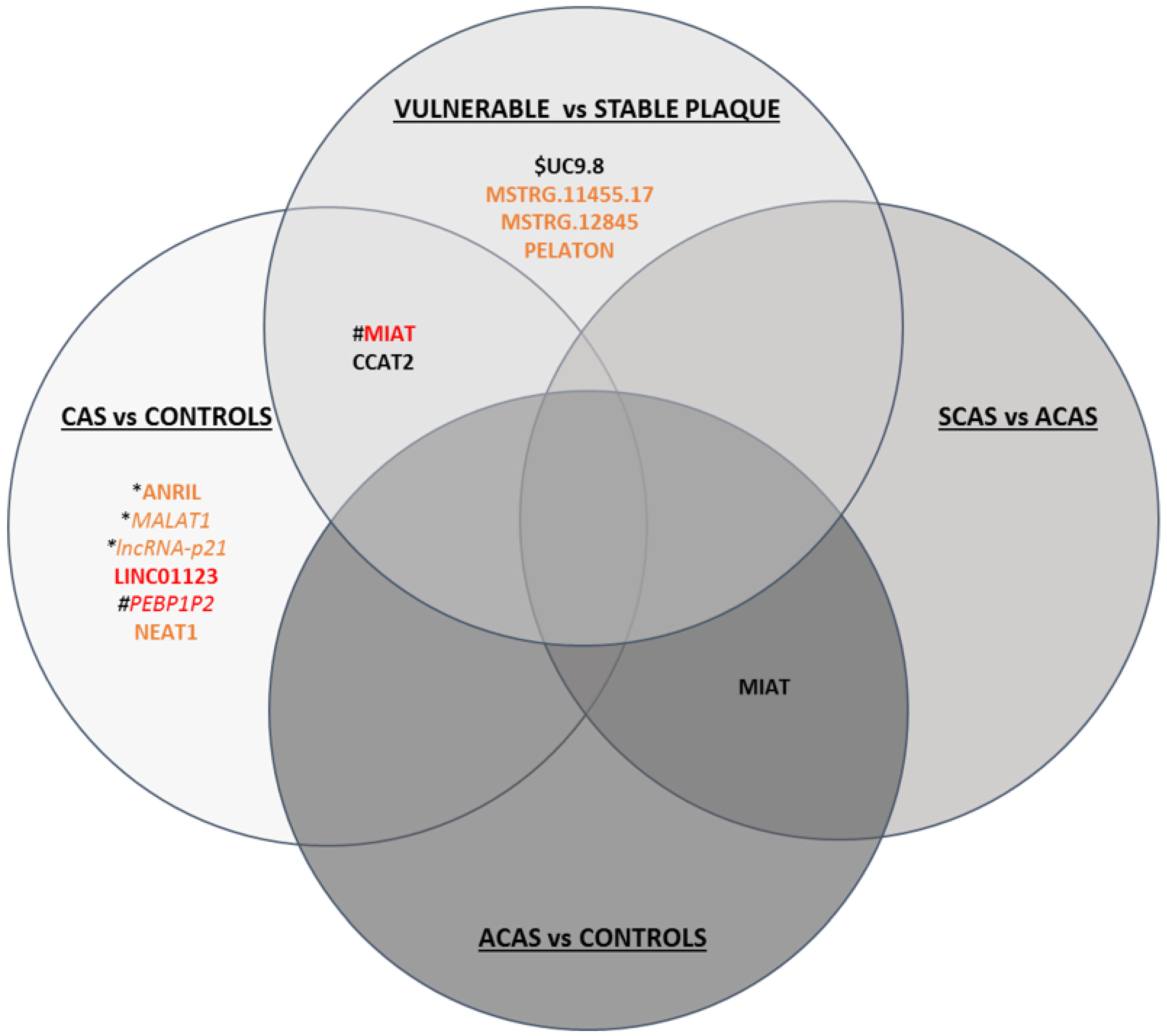
3.3.2. lncRNAs
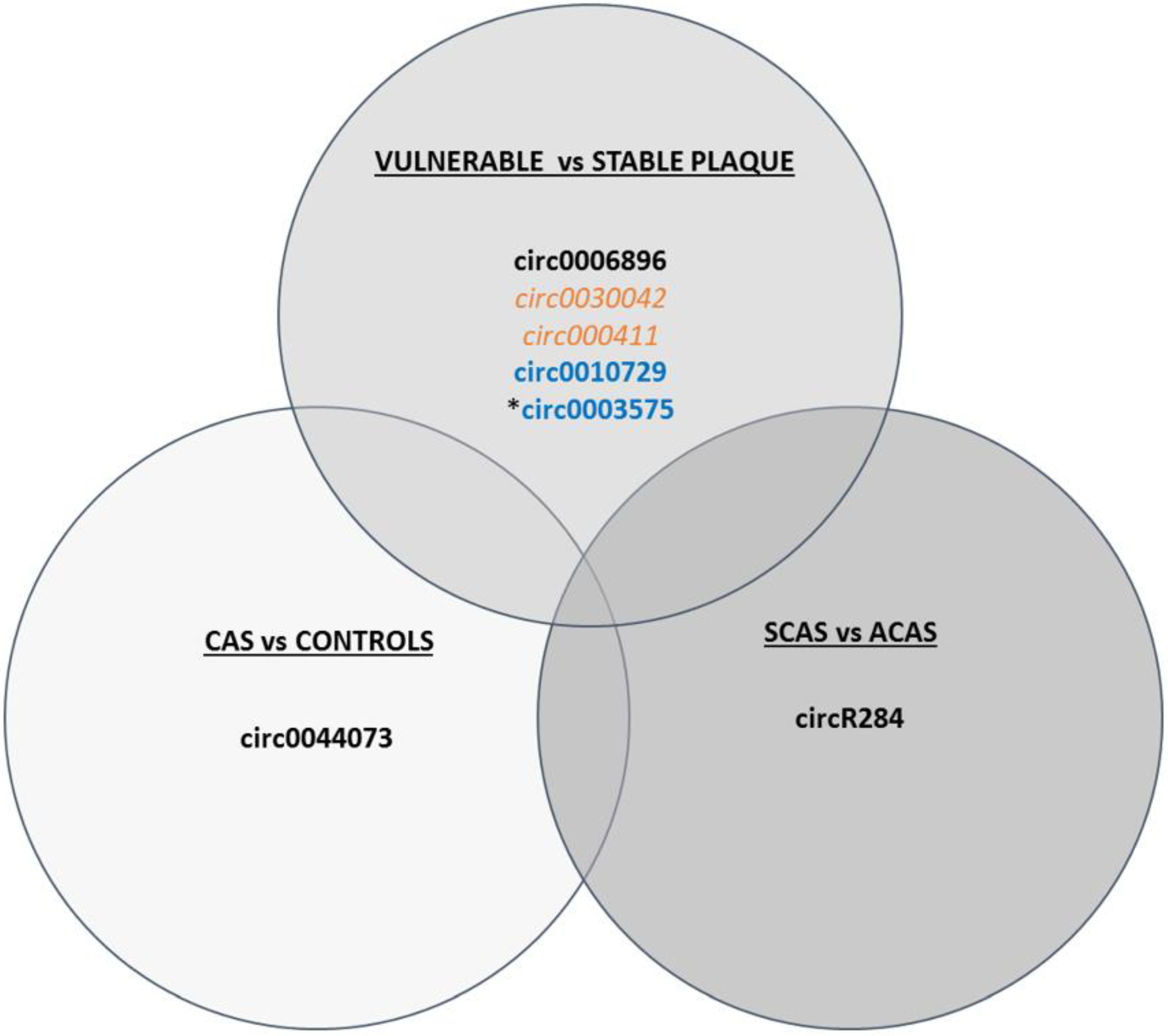
3.3.3. circRNAs
4. Limitations and Perspectives of the Epigenetic Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wafa, H.A.; Wolfe, C.D.A.; Emmett, E.; Roth, G.A.; Johnson, C.O.; Wang, Y. Burden of Stroke in Europe: Thirty-Year Projections of Incidence, Prevalence, Deaths, and Disability-Adjusted Life Years. Stroke 2020, 51, 2418–2427. [Google Scholar] [CrossRef]
- The GBD 2016 Lifetime Risk of Stroke Collaborators. Global, Regional, and Country-Specific Lifetime Risks of Stroke, 1990 and 2016. N. Engl. J. Med. 2018, 379, 2429–2437. [Google Scholar] [CrossRef]
- WHO. The Top 10 Causes of Death 9 December 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 30 March 2022).
- Tabas, I.; García-Cardeña, G.; Owens, G.K. Recent insights into the cellular biology of atherosclerosis. J. Cell Biol. 2015, 209, 13–22. [Google Scholar] [CrossRef]
- Mahmood, S.S.; Levy, D.; Vasan, R.S.; Wang, T.J. The Framinghan Heart Study and the Epidemiology of Cardiovascular Diseases: A Historical Perspective. Lancet 2014, 383, 999–1008. [Google Scholar] [CrossRef] [Green Version]
- Mechtouff, L.; Rascle, L.; Crespy, V.; Canet-Soulas, E.; Nighoghossian, N.; Millon, A. A narrative review of the pathophysiology of ischemic stroke in carotid plaques: A distinction versus a compromise between hemodynamic and embolic mechanism. Ann. Transl. Med. 2021, 9, 1208. [Google Scholar] [CrossRef]
- Barnett, H.J.; Taylor, D.W.; Eliasziw, M.; Fox, A.J.; Ferguson, G.G.; Haynes, R.B.; Rankin, R.N.; Clagett, G.P.; Hachinski, V.C.; Sackett, D.L.; et al. Benefit of carotid endarterectomy in patients with symptomatic moderate or severe stenosis. N. Engl. J. Med. 1998, 339, 1415–1425. [Google Scholar] [CrossRef]
- Halliday, A.; Mansfield, A.; Marro, J.; Peto, C.; Peto, R.; Potter, J.; Thomas, D. Prevention of disabling and fatal strokes by successful carotid endarterectomy in patients without recent neurological symptoms: Randomised controlled trial. Lancet 2004, 363, 1491–1502. [Google Scholar]
- Kleindorfer, D.O.; Towfighi, A.; Chaturvedi, S.; Cockroft, K.M.; Gutierrez, J.; Lombardi-Hill, D.; Kamel, H.; Kernan, W.; Kittner, S.J.; Leira, E.C.; et al. 2021 Guideline for the prevention of stroke in patients with stroke and transient ischemic attack; A guideline from the American Heart Association/American Stroke Association. Stroke 2021, 52, e364–e467. [Google Scholar] [CrossRef]
- Executive Committee for the Asymptomatic Carotid Atherosclerosis Study. Endarterectomy for Asymptomatic Carotid Artery Stenosis. JAMA 1995, 273, 1421–1428. [CrossRef]
- Bonati, L.H.; Kakkos, S.; Berkefeld, J.; de Borst, G.J.; Bulbulia, R.; Halliday, A.; van Heerzeele, I.; Knocar, I.; McCabe, D.J.; Lal, A.; et al. European Stroke Organisation guideline on endarterectomy and stenting for carotid artery stenosis. Eur. Stroke J. 2021, 6, 1–47. [Google Scholar] [CrossRef]
- Marquardt, L.; Geraghty, O.C.; Mehta, Z.; Rothwell, P.M. Low risk of ipsilateral stroke in patients with asymptomatic carotid stenosis on best medical treatment: A prospective, population-based study. Stroke 2010, 41, e11–e17. [Google Scholar] [CrossRef] [Green Version]
- Howard, D.P.J.; Gaziano, L.; Rothwell, P.M. Risk of stroke in relation to degree of asymptomatic carotid stenosis: A population-based cohort study, systematic review, and meta-analysis. Lancet Neurol. 2021, 20, 193–202. [Google Scholar] [CrossRef]
- Migdalski, A.; Jawien, A. New insight into biology, molecular diagnostics and treatment options of unstable carotid atherosclerotic plaque: A narrative review. Ann. Transl. Med. 2021, 9, 1207. [Google Scholar] [CrossRef]
- Naghavi, M.; Libby, P.; Falk, E.; Casscells, S.W.; Litovsky, S.; Rumberger, J.; Badmion, J.J.; Stefanadis, C.; Moreno, P.; Pasterkamp, G.; et al. From Vulnerable Plaque to Vulnerable Patient: A Call for New Definitions and Risk Assessment Strategies: Part II. Circulation 2003, 108, 1772–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puig, N.; Jiménez-Xarrié, E.; Camps-Renom, P.; Benitez, S. Search for reliable circulating biomarkers to predict carotid plaque vulnerability. Int. J. Mol. Sci. 2020, 21, 8236. [Google Scholar] [CrossRef]
- Alsheikh-Ali, A.A.; Kitsios, G.D.; Balk, E.M.; Lau, J.; Ip, S. The Vulnerable Atherosclerotic Plaque: Scope of the Literature. Ann. Intern. Med. 2010, 153, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Finn, A.V.; Nakano, M.; Narula, J.; Kolodgie, F.D.; Virmani, R. Concept of vulnerable/unstable plaque. Arter. Thromb. Vasc. Biol. 2010, 30, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Naghavi, M.; Libby, P.; Falk, E.; Casscells, S.W.; Litovsky, S.; Rumberger, J.; Badmion, J.J.; Stefanadis, C.; Moreno, P.; Pasterkamp, G.; et al. From vulnerable plaque to vulnerable patient: A call for new definitions and risk assessment strategies: Part I. Circulation 2003, 108, 1664–1672. [Google Scholar] [CrossRef]
- Saba, L.; Saam, T.; Jäger, H.R.; Yuan, C.; Hatsukami, T.S.; Saloner, D.; Wasserman, B.A.; Bonati, L.H.; Wintermark, M. Imaging biomarkers of vulnerable carotid plaques for stroke risk prediction and their potential clinical implications. Lancet Neurol. 2019, 18, 559–572. [Google Scholar] [CrossRef]
- Aboyans, V.; Ricco, J.B.; Bartelink, M.L.E.L.; Björck, M.; Brodmann, M.; Cohnert, T.; Collet, J.P.; Czerny, M.; De Carlo, M.; Debus, S.; et al. 2017 ESC Guidelines on the Diagnosis and Treatment of Peripheral Arterial Diseases, in collaboration with the European Society for Vascular Surgery (ESVS). Eur. Heart J. 2018, 39, 763–816. [Google Scholar] [CrossRef] [Green Version]
- Soler-Botija, C.; Gálvez-Montón, C.; Bayés-Genís, A. Epigenetic Biomarkers in Cardiovascular Diseases. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
- Saenger, A.K.; Christenson, R.H. Stroke biomarkers: Progress and challenges for diagnosis, prognosis, differentiation, and treatment. Clin. Chem. 2010, 56, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.; Martorell, J.; Riambau, V. Review of serum biomarkers in carotid atherosclerosis. J. Vasc. Surg. 2020, 71, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Kigka, V.I.; Potsika, V.; Mantzaris, M.; Tsakanikas, V.; Koncar, I.; Fotiadis, D.I. Serum biomarkers in carotid artery disease. Diagnostics 2021, 11, 2143. [Google Scholar] [CrossRef]
- Rothwell, P.M.; Warlow, C.P. Prediction of benefit from carotid endarterectomy in individual patients: A risk-modelling study. Lancet 1999, 353, 2105–2110. [Google Scholar] [CrossRef]
- North American Symptomatic Carotid Endarterectomy Trial Collaborators; Barnett, H.J.M.; Taylor, D.W.; Haynes, R.B.; Sackett, D.L.; Peerless, S.J.; Ferguson, G.G.; Fox, A.J.; Rankin, R.N.; Hachinski, V.C.; et al. Beneficial effect of carotid endarterectomy in symptomatic patients with high-grade carotid stenosis. N. Engl. J. Med. 1991, 325, 445–453. [Google Scholar]
- European Carotid Surgery Trialists Collaborative Group. Randomised trial of endarterectomy for recently symptomatic carotid stenosis: Final results of the MRC European Carotid Surgery Trial (ECST). Lancet 1998, 351, 1379–1387. [Google Scholar] [CrossRef]
- Caballero, P.E.J.; Segura Martín, T. Valores de normalidad de la reactividad vasomotora cerebral mediante el test de apnea voluntaria. Rev. Neurol. 2006, 43, 598–602. [Google Scholar]
- King, A.; Serena, J.; Bornstein, N.M.; Markus, H.S. Does impaired cerebrovascular reactivity predict stroke risk in asymptomatic carotid stenosis? A prospective substudy of the asymptomatic carotid emboli study. Stroke 2011, 42, 1550–1555. [Google Scholar] [CrossRef] [Green Version]
- Kleiser, B.; Widder, B. Course of Carotid Artery Occlusions with Impaired Cerebrovascular Reactivity. Stroke 1992, 23, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, M.; Gerds, T.A.; Grabiak, D.; Zimmermann, P.R.; Roth, M.; Guschlbauer, B.; Timmer, J.; Czosnyka, M.; Weiller, C.; Hetzel, A. Cerebral dysautoregulation and the risk of ischemic events in occlusive carotid artery disease. J. Neurol. 2008, 255, 1182–1189. [Google Scholar] [CrossRef]
- Klijn, C.J.; Kappelle, L.J.; Van Huffelen, A.C.; Visser, G.H.; Algra, A.; Tulleken, C.A.; van Gijn, J. Recurrent ischemia in symptomatic carotid occlusion Prognostic value of hemodynamic factors. Neurology 2000, 55, 1806–1812. [Google Scholar] [CrossRef]
- Amin-Hanjani, S.; Pandey, D.K.; Rose-Finnell, L.; Du, X.; Richardson, D.J.; Thulborn, K.R.; Elkind, M.S.; Zipfel, G.J.; Liebeskind, D.S.; Silver, F.L.; et al. Effect of hemodynamics on stroke risk in symptomatic atherosclerotic vertebrobasilar occlusive disease. JAMA Neurol. 2016, 73, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Markus, H.; Neuroscience, C.; George, S.; Markus, H.S.; King, A.; Shipley, M.; Topakian, R.; Cullinane, M.; Reihill, S.; Bornstein, N.M.; et al. Asymptomatic embolisation for prediction of stroke in the Asymptomatic Carotid Emboli Study (ACES): A prospective observational study. Lancet Neurol. 2010, 9, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Baradaran, H.; Foster, T.; Harrie, P.; Mcnally, J.S.; Alexander, M.; Pandya, A.; Anzai, Y.; Gupta, A. Carotid artery plaque characteristics: Current reporting practices on CT angiography. Neuroradiology 2021, 63, 1013–1018. [Google Scholar] [CrossRef]
- Coutinho, J.M.; Derkatch, S.; Potvin, A.R.J.; Tomlinson, G.; Kiehl, T.-R.; Silver, F.L.; Mandell, D.M. Nonstenotic carotid plaque on CT angiography in patients with cryptogenic stroke. Am. Acad. Neurol. 2016, 87, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Kesavabhotla, K.; Baradaran, H.; Kamel, H.; Pandya, A.; Giambrone, A.E.; Wright, D.; Pain, K.J.; Mtui, E.E.; Suri, J.S.; et al. Plaque echolucency and stroke risk in asymptomatic carotid stenosis: A systematic review and meta-analysis. Stroke 2015, 46, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, Y.; Yoshida, K.; Fukuda, H.; Handa, A.; Chin, M.; Yamagata, S. Asymptomatic Carotid T1-High-Intense Plaque as a Risk Factor for a Subsequent Cerebrovascular Ischemic Event. Cerebrovasc. Dis. 2017, 43, 250–256. [Google Scholar] [CrossRef]
- Sitzer, M.; Müller, W.; Siebler, M.; Hort, W.; Kniemeyer, H.-W.; Jäncke, L.; Steinmetz, H. Plaque Ulceration and Lumen Thrombus Are the Main Sources of Cerebral Microemboli in High-grade Internal Carotid Artery Stenosis. Stroke 1995, 26, 1231–1233. [Google Scholar] [CrossRef]
- Van Damme, H.; Vivario, M.; Boniver, J.; Limet, R. Histologic Characterization of Carotid Plaques. Cardiovasc. Pathol. 1994, 3, 9–17. [Google Scholar] [CrossRef]
- Carbone, F.; Montecucco, F.; Xu, S.; Banach, M.; Jamialahmadi, T.; Sahebkar, A. Epigenetics in atherosclerosis: Key features and therapeutic implications. Expert Opin. Ther. Targets 2020, 24, 719–721. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Yang, M.; Yang, H.; Xu, N.; Fan, X.; Liu, G.; Jiang, X.; Fan, J.; Zhang, L.; et al. DNA methylome profiling reveals epigenetic regulation of lipoprotein-associated phospholipase A2 in human vulnerable atherosclerotic plaque. Clin. Epigenetics 2021, 13, 161. [Google Scholar] [CrossRef]
- Zaina, S.; Gonçalves, I.; Carmona, F.J.; Gomez, A.; Heyn, H.; Mollet, I.G.; Moran, S.; Varol, N.; Esteller, M. DNA methylation dynamics in human carotid plaques after cerebrovascular events. Arter. Thromb. Vasc. Biol. 2015, 35, 1835–1842. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, B.; Irvin, M.R.; Sha, J.; Zhi, D.; Aslibekyan, S.; Absher, D.; Tiwari, H.K.; Kabagambe, E.K.; Ordovas, J.M.; Arnett, D.K. Epigenome-wide association study of fasting measures of glucose, insulin, and homa-ir in the genetics of lipid lowering drugs and diet network study. Diabetes 2014, 63, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Dick, K.J.; Nelson, C.P.; Tsaprouni, L.; Sandling, J.K.; Aïssi, D.; Wahl, S.; Meduri, E.; Morange, P.E.; Gagnon, F.; Grallert, H.; et al. DNA methylation and body-mass index: A genome-wide analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Zaina, S. Unraveling the DNA methylome of atherosclerosis. Curr. Opin. Lipidol. 2014, 25, 148–153. [Google Scholar] [CrossRef]
- Zaina, S.; Heyn, H.; Carmona, F.J.; Varol, N.; Sayols, S.; Condom, E.; Ramírez-Ruz, J.; Gomez, A.; Gonçalves, I.; Moran, S.; et al. DNA methylation map of human atherosclerosis. Circ. Cardiovasc. Genet. 2014, 7, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.Z.; Jiménez, J.M.; Ou, K.; McCormick, M.E.; Zhang, L.D.; Davies, P.F. Hemodynamic disturbed flow induces differential DNA methylation of endothelial Kruppel-like factor 4 promoter in vitro and in vivo. Circ. Res. 2014, 115, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Choi, B.G.; Jelinek, J.; Kim, D.H.; Lee, S.H.; Cho, K.; Rha, S.H.; Lee, Y.H.; Jin, H.S.; Choi, D.K.; et al. Promoter methylation changes in ALOX12 and AIRE1: Novel epigenetic markers for atherosclerosis. Clin. Epigenetics 2020, 12, 66. [Google Scholar] [CrossRef]
- De Vries, M.R.; Quax, P.H.A. Plaque angiogenesis and its relation to inflammation and atherosclerotic plaque destabilization. Curr. Opin. Lipidol. 2016, 27, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, J.; Yang, F.; Zhou, Y. Lp-PLA2 evaluates the severity of carotid artery stenosis and predicts the occurrence of cerebrovascular events in high stroke-risk populations. J. Clin. Lab. Anal. 2021, 35, e23691. [Google Scholar] [CrossRef]
- Jaiswal, S.; Natarajan, P.; Silver, A.J.; Gibson, C.J.; Bick, A.G.; Shvartz, E.; McConkey, M.; Gupta, N.; Gabriel, S.; Ardissino, D.; et al. Clonal Hematopoiesis and Risk of Atherosclerotic Cardiovascular Disease. N. Engl. J. Med. 2017, 377, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Fuster, J.J.; MacLauchlan, S.; Zuriaga, M.A.; Polackal, M.N.; Ostriker, A.C.; Chakraborty, R.; Wu, C.L.; Sano, S.; Muralidharan, S.; Rius, C.; et al. Clonal hematopoiesis associated with TET2 deficiency accelerates atherosclerosis development in mice. Science 2017, 355, 842–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Jin, Y.; Tang, W.H.; Qin, L.; Zhang, X.; Tellides, G.; Hwa, J.; Yu, J.; Martin, K.A. Ten-eleven translocation-2 (TET2) is a master regulator of smooth muscle cell plasticity. Circulation 2013, 128, 2047–2057. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Kamato, D.; Little, P.J.; Nakagawa, S.; Pelisek, J.; Jin, Z.G. Targeting epigenetics and non-coding RNAs in atherosclerosis: From mechanisms to therapeutics. Pharmacol. Ther. 2019, 196, 15–43. [Google Scholar] [CrossRef]
- Yu, J.; Qiu, Y.; Yang, J.; Bian, S.; Chen, G.; Deng, M.; Kang, H.; Huang, L. DNMT1-PPARγ pathway in macrophages regulates chronic inflammation and atherosclerosis development in mice. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Xie, S.A.; Zhang, T.; Wang, J.; Zhao, F.; Zhang, Y.P.; Yao, W.J.; Hur, S.S.; Yeh, Y.T.; Pang, W.; Zheng, L.S.; et al. Matrix stiffness determines the phenotype of vascular smooth muscle cell in vitro and in vivo: Role of DNA methyltransferase 1. Biomaterials 2018, 155, 203–216. [Google Scholar] [CrossRef]
- Katano, H.; Mase, M. Epigenetic changes in carotid plaques with high calcium scores; DNA methylation and microrna assessments. Atherosclerosis 2021, 331, e93. [Google Scholar] [CrossRef]
- Vlad, M.L.; Manea, S.A.; Lazar, A.G.; Raicu, M.; Muresian, H.; Simionescu, M.; Manea, A. Histone Acetyltransferase-Dependent Pathways Mediate Upregulation of NADPH Oxidase 5 in Human Macrophages under Inflammatory Conditions: A Potential Mechanism of Reactive Oxygen Species Overproduction in Atherosclerosis. Oxid. Med. Cell Longev. 2019, 2019, 3201062. [Google Scholar] [CrossRef] [Green Version]
- Manea, S.A.; Vlad, M.L.; Fenyo, I.M.; Lazar, A.G.; Raicu, M.; Muresian, H.; Simionescu, M.; Manea, A. Pharmacological inhibition of histone deacetylase reduces NADPH oxidase expression, oxidative stress and the progression of atherosclerotic lesions in hypercholesterolemic apolipoprotein E-deficient mice; potential implications for human atherosclerosis. Redox Biol. 2020, 28, 101338. [Google Scholar] [CrossRef] [PubMed]
- Greißel, A.; Culmes, M.; Burgkart, R.; Zimmermann, A.; Eckstein, H.-H.; Zernecke, A.; Pelisek, J. Histone acetylation and methylation significantly change with severity of atherosclerosis in human carotid plaques. Cardiovasc. Pathol. 2016, 25, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Winnik, S.; Gaul, D.S.; Preitner, F.; Lohmann, C.; Weber, J.; Miranda, M.X.; Liu, Y.; van Tits, L.J.; Mateos, J.M.; Brokopp, C.E.; et al. Deletion of Sirt3 does not affect atherosclerosis but accelerates weight gain and impairs rapid metabolic adaptation in LDL receptor knockout mice: Implications for cardiovascular risk factor development. Basic Res. Cardiol. 2014, 109, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazan, H.A.; Hatfield, S.A.; Brug, A.; Brooks, A.J.; Lightell, D.J.; Woods, T.C. Carotid Plaque Rupture Is Accompanied by an Increase in the Ratio of Serum circR-284 to miR-221 Levels. Circ. Cardiovasc. Genet. 2017, 10, e001720. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Li, J.; Cairns, M.J. Identifying miRNAs, targets and functions. Brief. Bioinform. 2014, 15, 336. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Han, J.; Chen, J.; Dong, J.; Xia, Y.; Liu, J.; Jian, Y.; Dai, J.; Lu, J.; Jin, G.; et al. Plasma miRNAs as early biomarkers for detecting hepatocellular carcinoma. Int. J. Cancer 2015, 137, 1679–1690. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Li, Y.; Wang, P.; Yu, P.; Lang, L. Circulating miR-342-5p serves as a diagnostic biomarker in patients with carotid artery stenosis and predicts the occurrence of the cerebral ischemic event. Ir. J. Med. Sci. 2021, 191, 713–718. [Google Scholar] [CrossRef]
- Tafrihi, M.; Hasheminasab, E. MiRNAs: Biology, Biogenesis, their Web-based Tools, and Databases. MicroRNA 2018, 8, 4–27. [Google Scholar] [CrossRef]
- Chen, G.; Gao, J.; Sheng, Y.; Han, X.; Ji, X.; Zhao, M.; Wu, J. Diagnostic value of miR-92a in asymptomatic carotid artery stenosis patients and its ability to predict cerebrovascular events. Diagn. Pathol. 2020, 15, 74. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, W.; Zhang, T.; Zhao, H.; He, S.; Li, B.; Gao, Y.; Pan, W. Diagnostic value of miR-186-5p for carotid artery stenosis and its predictive significance for future cerebral ischemic event. Diagn. Pathol. 2020, 15, 101. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, J.; Jiang, W.; Wang, F. Analysis of the diagnostic and prognostic value of miR-9-5p in carotid artery stenosis. Bosn J. Basic Med. Sci. 2021, 21, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; He, X.Y.; Xu, M. The Role of miRNA-146a and Proinflammatory Cytokines in Carotid Atherosclerosis. Biomed. Res. Int. 2020, 2020, 6657734. [Google Scholar] [CrossRef]
- Luque, A.; Farwati, A.; Krupinski, J.; Aran, J.M. Association between low levels of serum miR-638 and atherosclerotic plaque vulnerability in patients with high-grade carotid stenosis. J. Neurosurg. 2019, 131, 72–79. [Google Scholar] [CrossRef]
- Minin, E.O.Z.; Paim, L.R.; Lopes, E.C.P.; Bueno, L.C.M.; Carvalho-Romano, L.F.R.S.; Marques, E.R.; Vegian, C.; Pio-Magalhaes, J.A.; Coelho-Filho, O.R.; Sposito, A.C.; et al. Association of circulating mir-145-5p and mir-let7c and atherosclerotic plaques in hypertensive patients. Biomolecules 2021, 11, 1840. [Google Scholar] [CrossRef]
- Nie, P.; Yang, F.; Wan, F.; Jin, S.; Pu, J. Analysis of MicroRNAs Associated with Carotid Atherosclerotic Plaque Rupture with Thrombosis. Front. Genet. 2021, 12, 599350. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, Z.; Yang, X.; Fu, R.; Wang, J.; Xu, H. Circulating miR-106b-5p serves as a diagnostic biomarker for asymptomatic carotid artery stenosis and predicts the occurrence of cerebral ischemic events. Vasc. Med. 2020, 25, 436–442. [Google Scholar] [CrossRef]
- Cipollone, F.; Felicioni, L.; Sarzani, R.; Ucchino, S.; Spigonardo, F.; Mandolini, C.; Malatesta, S.; Bucci, M.; Mammarella, C.; Santovito, D.; et al. A unique MicroRNA signature associated with plaque instability in humans. Stroke 2011, 42, 2556–2563. [Google Scholar] [CrossRef] [Green Version]
- Maitrias, P.; Metzinger-Le Meuth, V.; Massy, Z.A.; M’Baya-Moutoula, E.; Reix, T.; Caus, T.; Metzinger, L. MicroRNA deregulation in symptomatic carotid plaque. J. Vasc. Surg. 2015, 62, 1245–1250.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badacz, R.; Przewłocki, T.; Gacoń, J.; Stępień, E.; Enguita, F.J.; Karch, I.; Zmudka, K.; Kablak-Ziembicka, A. Circulating miRNA levels differ with respect to carotid plaque characteristics and symptom occurrence in patients with carotid artery stenosis and provide information on future cardiovascular events. Adv. Interv. Cardiol. 2018, 14, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Grosse, G.M.; Derda, A.A.; Stauss, R.D.; Neubert, L.; Jonigk, D.D.; Kühnel, M.P.; Gabriel, M.M.; Schuppner, R.; Wilhelmi, M.; Bär, C.; et al. Circulating microRNAs in Symptomatic and Asymptomatic Carotid Stenosis. Front. Neurol. 2021, 12, 755827. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Cao, Q.; Meng, M.; Wang, X. MicroRNA-186-5p serves as a diagnostic biomarker in atherosclerosis and regulates vascular smooth muscle cell proliferation and migration. Cell Mol. Biol. Lett. 2020, 25, 27. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Fernández-Hernando, C. From evolution to revolution: MiRNAs as pharmacological targets for modulating cholesterol efflux and reverse cholesterol transport. Pharmacol. Res. 2013, 75, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Jiang, L.; Wang, X. Aberrant expression of miR-483-5p in patients with asymptomatic carotid artery stenosis and its predictive value for cerebrovascular event occurrence. Exp. Ther. Med. 2021, 22, 1101. [Google Scholar] [CrossRef]
- Wang, W.Y.; Zheng, Y.S.; Li, Z.G.; Cui, Y.M.; Jiang, J.C. MiR-92a contributes to the cardiovascular disease development in diabetes mellitusthrough NF-κB and downstream inflammatory pathways. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3070–3079. [Google Scholar]
- Huang, H.T.; Liu, Z.C.; Wu, K.Q.; Gu, S.R.; Lu, T.C.; Zhong, C.J.; Zhou, Y.X. MiR-92a regulates endothelial progenitor cells (EPCs) by targeting GDF11 via activate SMAD2/3/FAK/Akt/eNOS pathway. Ann. Transl. Med. 2019, 7, 563. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Zhang, S.; Yan, S.; Wang, Z.; Wang, C.; Zhang, X. MiR-30e and miR-92a are related to atherosclerosis by targeting ABCA1. Mol. Med. Rep. 2019, 19, 3298–3304. [Google Scholar] [CrossRef] [Green Version]
- Bi, S.; Peng, Q.; Liu, W.; Zhang, C.; Liu, Z. MicroRNA-342-5p activates the Akt signaling pathway by downregulating PIK3R1 to modify the proliferation and differentiation of vascular smooth muscle cells. Exp. Ther. Med. 2020, 20, 239. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, H.; Liang, J.; Li, Y.; Li, X. MicroRNA-503-5p improves carotid artery stenosis by inhibiting the proliferation of vascular smooth muscle cells. Exp. Ther. Med. 2020, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yan, S.; Yuan, Y.; Ji, S.; Guo, L. miR-28-5p improved carotid artery stenosis by regulating vascular smooth muscle cell proliferation and migration. Vascular 2021. [Google Scholar] [CrossRef] [PubMed]
- Knoka, E.; Trusinskis, K.; Mazule, M.; Briede, I.; Crawford, W.; Jegere, S.; Kumsars, I.; Narbute, I.; Sondore, D.; Lejnieks, A.; et al. Circulating plasma microRNA-126, microRNA-145, and microRNA-155 and their association with atherosclerotic plaque characteristics. J. Clin. Transl. Res. 2019, 5, 60–67. [Google Scholar]
- Eken, S.M.; Jin, H.; Chernogubova, E.; Li, Y.; Simon, N.; Sun, C.; Korzunowicz, G.; Busch, A.; Bäcklund, A.; Österholm, C.; et al. MicroRNA-210 enhances fibrous cap stability in advanced atherosclerotic lesions. Circ. Res. 2017, 120, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Dolz, S.; Górriz, D.; Tembl, J.I.; Sánchez, D.; Fortea, G.; Parkhutik, V.; Lago, A. Circulating MicroRNAs as novel biomarkers of stenosis progression in asymptomatic carotid stenosis. Stroke 2017, 48, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Qin, Y.; Zhu, G.; Li, Y.; Xue, J. Low serum miR-320b expression as a novel indicator of carotid atherosclerosis. J. Clin. Neurosci. 2016, 33, 252–258. [Google Scholar] [CrossRef]
- Huang, C.-Q.; Jin, W.-X.; Yu, G.-F. Correlation between carotid atherosclerotic plaque properties and serum levels of lncRNA CCAT2 and miRNA-216b. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7033–7038. [Google Scholar] [PubMed]
- Feng, B.; Chakrabarti, S. miR-320 Regulates Glucose-Induced Gene Expression in Diabetes. ISRN Endocrinol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Ye, Z.; Ming, C.; Luo, S.; Ying, X.; Chen, S.; Mao, L.; Li, Y.; Jin, H.; Yu, C.; et al. MicroRNA-23a-5p promotes atherosclerotic plaque progression and vulnerability by repressing ATP-binding cassette transporter A1/G1 in macrophages. J. Mol. Cell Cardiol. 2018, 123, 139–149. [Google Scholar] [CrossRef]
- Jin, H.; Li, D.Y.; Chernogubova, E.; Sun, C.; Busch, A.; Eken, S.M.; Saliba-Gustafsson, P.; Winter, H.; Winski, G.; Raaz, U.; et al. Local Delivery of miR-21 Stabilizes Fibrous Caps in Vulnerable Atherosclerotic Lesions. Mol. Ther. 2018, 26, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Kumric, M.; Borovac, J.A.; Martinovic, D.; Kurir, T.T.; Bozic, J. Circulating biomarkers reflecting destabilization mechanisms of coronary artery plaques: Are we looking for the impossible? Biomolecules 2021, 11, 881. [Google Scholar] [CrossRef] [PubMed]
- Bazan, H.A.; Hatfield, S.A.; O’Malley, C.B.; Brooks, A.J.; Lightell, D.; Woods, T.C. Acute Loss of MIR-221 and MIR-222 in the Atherosclerotic Plaque Shoulder Accompanies Plaque Rupture. Stroke 2015, 46, 3285–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Sun, Y.; Han, T.; Zhu, J.; Xie, Y.; Wang, S.; Wu, Y.; Fan, Y.; Sun, X.; Zhou, J.; et al. Upregulation of miR-330-5p is associated with carotid plaque’s stability by targeting Talin-1 in symptomatic carotid stenosis patients. BMC Cardiovasc. Disord. 2019, 19, 149. [Google Scholar] [CrossRef] [PubMed]
- Magenta, A.; Sileno, S.; D’Agostino, M.; Persiani, F.; Beji, S.; Paolini, A.; Camili, D.; Platone, A.; Capogrossi, M.C.; Fugiuele, S. Atherosclerotic plaque instability in carotid arteries: MiR-200c as a promising biomarker. Clin. Sci. 2018, 132, 2423–2436. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, J.; Chen, Y.; Zhang, S.; Feng, C.; Hou, Z.; Cai, J.; Wang, Y.; Hui, R.; Lv, B.; et al. MicroRNA-216a promotes M1 macrophages polarization and atherosclerosis progression by activating telomerase via the Smad3/NF-κB pathway. Biochim. Biophys. Acta -Mol. Basis Dis. 2019, 1865, 1772–1781. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Cao, Y.; Li, H.; Hu, Z.; Zhang, H.; Zhang, L.; Su, W.; Xu, Y.; Liang, L.; Melgiri, N.D.; et al. miR-532-3p-CSF2RA Axis as a Key Regulator of Vulnerable Atherosclerotic Plaque Formation. Can. J. Cardiol. 2020, 36, 1782–1794. [Google Scholar] [CrossRef]
- Katano, H.; Nishikawa, Y.; Yamada, H.; Yamada, K.; Mase, M. Differential Expression of microRNAs in Severely Calcified Carotid Plaques. J. Stroke Cerebrovasc. Dis. 2018, 27, 108–117. [Google Scholar] [CrossRef]
- Vasuri, F.; Ciavarella, C.; Fittipaldi, S.; Pini, R.; Vacirca, A.; Gargiulo, M.; Faggioli, G.; Pasquinelli, G. Different histological types of active intraplaque calcification underlie alternative miRNA-mRNA axes in carotid atherosclerotic disease. Virchows Arch. 2020, 476, 307–316. [Google Scholar] [CrossRef]
- Arslan, S.; Berkan, Ö.; Lalem, T.; Özbilüm, N.; Göksel, S.; Korkmaz, Ö.; Çetin, N.; Devaux, Y. Long non-coding RNAs in the atherosclerotic plaque. Atherosclerosis 2017, 266, 176–181. [Google Scholar] [CrossRef]
- Ye, Z.M.; Yang, S.; Xia, Y.P.; Hu, R.T.; Chen, S.; Li, B.W.; Chen, S.L.; Luo, X.Y.; Mao, L.; Li, Y.; et al. LncRNA MIAT sponges miR-149-5p to inhibit efferocytosis in advanced atherosclerosis through CD47 upregulation. Cell Death Dis. 2019, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Fasolo, F.; Jin, H.; Winski, G.; Chernogubova, E.; Pauli, J.; Winter, H.; Li, D.; Glukha, N.; Bauer, S.; Metschl, S.; et al. Long Noncoding RNA MIAT Controls Advanced Atherosclerotic Lesion Formation and Plaque Destabilization. Circulation 2021, 144, 1567–1583. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Cai, J.; Han, Y.; Chen, J.; Huang, Z.P.; Chen, C.; Cai, Y.; Huang, H.; Yang, Y.; Liu, Y.; et al. LincRNA-p21 regulates neointima formation, vascular smooth muscle cell proliferation, apoptosis, and atherosclerosis by enhancing p53 activity. Circulation 2014, 130, 1452–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; He, F.; Liang, B.; Jing, Y.; Zhang, P.; Liu, W.; Zhu, B.; Dou, D. LincRNA-p21 alleviates atherosclerosis progression through regulating the miR-221/SIRT1/Pcsk9 axis. J. Cell. Mol. Med. 2021, 25, 9141–9153. [Google Scholar] [CrossRef]
- Weng, G.; Gu, M.; Zhang, Y.; Zhao, G.; Gu, Y. LINC01123 promotes cell proliferation and migration via regulating miR-1277-5p/KLF5 axis in ox-LDL-induced vascular smooth muscle cells. J. Mol. Histol. 2021, 52, 943–953. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lian, Z.; Yang, Y.; Wang, Z.; Fu, X.; Liu, Y.; Li, M.; Tian, J.; Yu, T.; Xin, H. Long Non-coding RNA PEBP1P2 Suppresses Proliferative VSMCs Phenotypic Switching and Proliferation in Atherosclerosis. Mol. Ther. Nucleic Acids 2020, 22, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Vlachogiannis, N.I.; Sachse, M.; Georgiopoulos, G.; Zormpas, E.; Bampatsias, D.; Delialis, D.; Bonini, F.; Glayfos, G.; Sigala, F.; Stamatelopoulos, K.; et al. Adenosine-to-inosine Alu RNA editing controls the stability of the pro-inflammatory long noncoding RNA NEAT1 in atherosclerotic cardiovascular disease. J. Mol. Cell. Cardiol. 2021, 160, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, E.J.; van Solingen, C.; Scacalossi, K.R.; Ouimet, M.; Afonso, M.S.; Prins, J.; Koelwyn, G.J.; Sharma, M.; Ramkhelawon, B.; Carpenter, S.; et al. The long noncoding RNA CHROME regulates cholesterol homeostasis in primates. Nat. Metab. 2019, 1, 98–110. [Google Scholar] [CrossRef]
- Fan, Z.; Zhang, Y.; Xiao, D.; Ma, J.; Liu, H.; Shen, L.; Zhang, M.; He, B. Long noncoding RNA UC.98 stabilizes atherosclerotic plaques by promoting the proliferation and adhesive capacity in murine aortic endothelial cells. Acta Biochim. Biophys. Sin. 2020, 52, 141–149. [Google Scholar] [CrossRef]
- Bao, M.H.; Zhang, R.Q.; Huang, X.S.; Zhou, J.; Guo, Z.; Xu, B.F.; Liu, R. Transcriptomic and Proteomic Profiling of Human Stable and Unstable Carotid Atherosclerotic Plaques. Front. Genet. 2021, 12, 2129. [Google Scholar] [CrossRef]
- Hung, J.; Scanlon, J.P.; Mahmoud, A.D.; Rodor, J.; Ballantyne, M.; Fontaine, M.; Temmerman, L.; Kaczynski, J.; Connor, K.L.; Bhushan, R.; et al. Novel plaque enriched long noncoding RNA in atherosclerotic macrophage regulation (PELATON). Arter. Thromb. Vasc. Biol. 2020, 40, 697–713. [Google Scholar] [CrossRef]
- Rayner, K.J. Leading the long noncoding RNA pack: PELATON in human atherosclerosis. Arter. Thromb. Vasc. Biol. 2020, 40, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Li, X.; Li, C.; Li, Y.; He, Y.; Li, F.; Ling, L. Intracerebral hemorrhage alters circular RNA expression profiles in the rat brain. Am. J. Transl. Res. 2020, 12, 4160–4174. [Google Scholar] [PubMed]
- Yu, F.; Zhang, Y.; Wang, Z.; Gong, W.; Zhang, C. Hsa_circ_0030042 regulates abnormal autophagy and protects atherosclerotic plaque stability by targeting eIF4A3. Theranostics 2021, 11, 5404–5417. [Google Scholar] [CrossRef] [PubMed]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Ji, H.; Zhang, H.; Yang, J.; Guo, R.; Wang, J. circANRIL reduces vascular endothelial injury, oxidative stress and inflammation in rats with coronary atherosclerosis. Exp. Ther. Med. 2020, 20, 2245–2251. [Google Scholar] [CrossRef]
- Li, C.Y.; Ma, L.; Yu, B. Circular RNA hsa_circ_0003575 regulates oxLDL induced vascular endothelial cells proliferation and angiogenesis. Biomed. Pharmacother. 2017, 95, 1514–1519. [Google Scholar] [CrossRef]
- Shen, L.; Hu, Y.; Lou, J.; Yin, S.; Wang, W.; Wang, Y.; Xia, Y.; Wu, W. CircRNA-0044073 is upregulated in atherosclerosis and increases the proliferation and invasion of cells by targeting miR-107. Mol. Med. Rep. 2019, 49, 3923–3932. [Google Scholar] [CrossRef]
- Wen, Y.; Chun, Y.; Qing Lian, Z.; Wei Yong, Z.; Mei Lan, Y.; Huan, L.; Xi, C.Y.; Juan, L.S.; Qing, Z.W.; Jia, C.; et al. circRNA-0006896-miR1264-DNMT1 axis plays an important role in carotid plaque destabilization by regulating the behavior of endothelial cells in atherosclerosis. Mol. Med. Rep. 2021, 23, 311. [Google Scholar] [CrossRef]
- Dang, R.Y.; Liu, F.L.; Li, Y. Circular RNA hsa_circ_0010729 regulates vascular endothelial cell proliferation and apoptosis by targeting the miR-186/HIF-1α axis. Biochem. Biophys. Res. Commun. 2017, 490, 104–110. [Google Scholar] [CrossRef]
- Zhao, C.; Lv, Y.; Duan, Y.; Li, G.; Zhang, Z. Circulating non-coding RNAs and Cardiovascular Diseases. Adv. Exp. Med. Biol. 2020, 1229, 357–367. [Google Scholar]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- De Gonzalo-Calvo, D.; Vea, A.; Bär, C.; Fiedler, J.; Couch, L.S.; Brotons, C.; Llorente-Cortes, V.; Thum, T. Circulating non-coding RNAs in biomarker-guided cardiovascular therapy: A novel tool for personalized medicine? Eur. Heart J. 2019, 40, 1643–1650. [Google Scholar] [CrossRef]
- Garciá-Giménez, J.L.; Mena-Mollá, S.; Beltrán-Garciá, J.; Sanchis-Gomar, F. Challenges in the analysis of epigenetic biomarkers in clinical samples. Clin. Chem. Lab. Med. 2017, 55, 1474–1477. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.B.; Feinberg, M.W. Long Noncoding RNAs in Atherosclerosis and Vascular Injury: Pathobiology, Biomarkers, and Targets for Therapy Jacob. Arter. Thromb. Vasc. Biol. 2020, 40, 2002–2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yuan, Y.; Cho, J.H.; McClarty, S.; Baxter, D.; Galas, D.J. Comparing the MicroRNA spectrum between serum and plasma. PLoS ONE 2012, 7, e41561. [Google Scholar] [CrossRef]
- Sriram, H.; Khanka, T.; Kedia, S.; Tyagi, P.; Ghogale, S.; Deshpande, N.; Chatterjee, G.; Rajpal, S.; Patkar, N.V.; Subramanian, P.G.; et al. Improved protocol for plasma microrna extraction and comparison of commercial kits. Biochem. Med. 2021, 31, 467–475. [Google Scholar] [CrossRef]
- Marabita, F.; De Candia, P.; Torri, A.; Tegnér, J.; Abrignani, S.; Rossi, R.L. Normalization of circulating microRNA expression data obtained by quantitative real-time RT-PCR. Brief. Bioinform. 2016, 17, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Gubern, C.; Hurtado, O.; Rodríguez, R.; Morales, J.R.; Romera, V.G.; Moro, M.A.; Lizasoain, I.; Serena, J.; Mallolas, J. Validation of housekeeping genes for quantitative real-time PCR in in-vivo and in-vitro models of cerebral ischaemia. BMC Mol. Biol. 2009, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Which is the accurate data normalization strategy for microRNA quantification? Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Faraldi, M.; Gomarasca, M.; Banfi, G.; Lombardi, G. Free Circulating miRNAs Measurement in Clinical Settings: The Still Unsolved Issue of the Normalization. Adv. Clin. Chem. 2018, 87, 113–139. [Google Scholar] [PubMed]
- Wan, N.; Jiang, Y.; Huang, J.; Oueslati, R.; Eda, S.; Wu, J.; Lin, X. Rapid and sensitive detection of mirna based on ac electrokinetic capacitive sensing for point-of-care applications. Sensors 2021, 21, 3985. [Google Scholar] [CrossRef] [PubMed]
- Zanchetta, G.; Carzaniga, T.; Vanjur, L.; Casiraghi, L.; Tagliabue, G.; Morasso, C.; Bellini, T.; Buscaglia, M. Design and optimization of a rapid, multiplex miRNA assay without washing steps. Proceedings 2020, 60, 7040. [Google Scholar]
- Ranjan, S.; Jain, S.; Bhargava, A.; Shandilya, R.; Srivastava, R.K.; Mishra, P.K. Lateral flow assay-based detection of long non-coding RNAs: A point-of-care platform for cancer diagnosis. J. Pharm. Biomed. Anal. 2021, 204, 114285. [Google Scholar] [CrossRef]
- Shandilya, R.; Kumari, R.; Singh, R.D.; Chouksey, A.; Bhargava, A.; Goryacheva, I.Y.; Mishra, P.K. Gold based nano-photonic approach for point-of-care detection of circulating long non-coding RNAs. Nanomed. Nanotechnol. Biol. Med. 2021, 36, 102413. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, M.; Cao, J.; Liang, Y.; Tu, T.; Hu, J.; Li, T.; Cai, Y.; Li, S.; Liu, B.; et al. An integrated electrochemical POCT platform for ultrasensitive circRNA detection towards hepatocellular carcinoma diagnosis. Biosens. Bioelectron. 2021, 192, 113500. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Greater Net Benefit if Revascularisation Performed | Lower Net Benefit if Revascularisation Performed |
|---|---|
| ICA stenosis >70% | ICA stenosis 50–69% |
| Male | Female |
| <70 years old | >75 years old |
| Diabetic | Not diabetic |
| Stroke | TIA or asymptomatic |
| Hemispheric symptoms | Amaurosis fugax |
| Plaque characteristics: hypoechoic, irregular surface, ulcerated, high volume, progression | Calcified plaque |
| Decreased CVR. Poor collateral circulation | Good collateral circulation |
| MES+ | MES− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carballo-Perich, L.; Puigoriol-Illamola, D.; Bashir, S.; Terceño, M.; Silva, Y.; Gubern-Mérida, C.; Serena, J. Clinical Parameters and Epigenetic Biomarkers of Plaque Vulnerability in Patients with Carotid Stenosis. Int. J. Mol. Sci. 2022, 23, 5149. https://doi.org/10.3390/ijms23095149
Carballo-Perich L, Puigoriol-Illamola D, Bashir S, Terceño M, Silva Y, Gubern-Mérida C, Serena J. Clinical Parameters and Epigenetic Biomarkers of Plaque Vulnerability in Patients with Carotid Stenosis. International Journal of Molecular Sciences. 2022; 23(9):5149. https://doi.org/10.3390/ijms23095149
Chicago/Turabian StyleCarballo-Perich, Laia, Dolors Puigoriol-Illamola, Saima Bashir, Mikel Terceño, Yolanda Silva, Carme Gubern-Mérida, and Joaquín Serena. 2022. "Clinical Parameters and Epigenetic Biomarkers of Plaque Vulnerability in Patients with Carotid Stenosis" International Journal of Molecular Sciences 23, no. 9: 5149. https://doi.org/10.3390/ijms23095149
APA StyleCarballo-Perich, L., Puigoriol-Illamola, D., Bashir, S., Terceño, M., Silva, Y., Gubern-Mérida, C., & Serena, J. (2022). Clinical Parameters and Epigenetic Biomarkers of Plaque Vulnerability in Patients with Carotid Stenosis. International Journal of Molecular Sciences, 23(9), 5149. https://doi.org/10.3390/ijms23095149