
Nutritional Management of Thyroiditis of Hashimoto


 , , , and
, , , and
Abstract
:1. Introduction
2. Nutritional Factors Linked to TH Etiopathogenesis
2.1. Nutritional Deficits or Excess
2.2. Nutritional Elements Generating Intoxication
2.3. Cyanotoxins and Thyroid Function
2.4. Diets Favoring Inflammation and Gut Dysbiosis and Their Role in Thyroiditis of Hashimoto
2.5. Proinflammatory Nutrients Favoring Clinical Manifestation of Endocrine and Metabolic Disease
3. The Role of Thyroid–Gut Axis and the Influence of Gut Microbiota on TH
4. Endocrine and Immune Regulators Involved in Adaptation: Vitamin D and Melatonin
4.1. Essential Effects of Vitamin D and Melatonin on Body Physiology and Cell Function
4.2. Deficits of Vitamin D and Melatonin Linked to Autoimmune Predisposition
5. Nutritional Treatment Strategies of Thyroiditis of Hashimoto
5.1. Anti-Inflammatory Nutrients
5.2. Adaptive Responses Involved in the Control of Immune Reactions
5.3. Beneficial Effects of Vitamin D and Melatonin as Antioxidants
5.4. Diets Aimed at Liver Detoxication and Cleansing
5.5. Ecological Dietary Regimen for TH
6. Perspectives for Nutritional Management of Thyroiditis of Hashimoto
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, S.; Rayman, M.P. Multiple Nutritional Factors and the Risk of Hashimoto’s Thyroiditis. Thyroid 2017, 27, 597–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okayasu, I.; Hara, Y.; Nakamura, K.; Rose, N.R. Racial and age-related differences in incidence and severity of focal autoimmune thyroiditis. Am. J. Clin. Pathol. 1994, 101, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Krysiak, R.; Szkróbka, W.; Okopień, B. The Effect of Gluten-Free Diet on Thyroid Autoimmunity in Drug-Naïve Women with Hashimoto’s Thyroiditis: A Pilot Study. Exp. Clin. Endocrinol. Diabetes 2019, 127, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Ihnatowicz, P.; Drywień, M.; Wątor, P.; Wojsiat, J. The importance of nutritional factors and dietary management of Hashimoto’s thyroiditis. Ann. Agric. Environ. Med. 2020, 27, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Caturegli, P.; De Remigis, A.; Rose, N.R. Hashimoto thyroiditis: Clinical and diagnostic criteria. Autoimmun. Rev. 2014, 13, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Subekti, I.; Pramono, L.A. Current diagnosis and management of Graves’ disease. Acta Med. Indones. 2018, 50, 177–182. [Google Scholar] [PubMed]
- Wiersinga, W.M. Clinical Relevance of Environmental Factors in the Pathogenesis of Autoimmune Thyroid Disease. Endocrinol. Metab. 2016, 31, 213–222. [Google Scholar] [CrossRef]
- Weetman, A.P. An update on the pathogenesis of Hashimoto’s thyroiditis. J. Endocrinol. Investig. 2021, 44, 883–890. [Google Scholar] [CrossRef]
- Rayman, M.P. Multiple nutritional factors and thyroid disease, with particular reference to autoimmune thyroid disease. Proc. Nutr. Soc. 2019, 78, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Benvenga, S.; Ferrari, S.M.; Elia, G.; Ragusa, F.; Patrizio, A.; Paparo, S.R.; Camastra, S.; Bonofiglio, D.; Antonelli, A.; Fallahi, P. Nutraceuticals in Thyroidology: A Review of in Vitro, and in Vivo Animal Studies. Nutrients 2020, 12, 1337. [Google Scholar] [CrossRef]
- Ceballos, D.; Hernández-Camba, A.; Ramos, L. Diet and microbiome in the beginning of the sequence of gut inflammation. World J. Clin. Cases 2021, 9, 11122–11147. [Google Scholar] [CrossRef] [PubMed]
- Kochman, J.; Jakubczyk, K.; Bargiel, P.; Janda-Milczarek, K. The Influence of Oxidative Stress on Thyroid Diseases. Antioxidants 2021, 10, 1442. [Google Scholar] [CrossRef] [PubMed]
- Cayres, L.C.F.; de Salis, L.V.V.; Rodrigues, G.S.P.; Lengert, A.V.H.; Biondi, A.P.C.; Sargentini, L.D.B.; Brisotti, J.L.; Gomes, E.; de Oliveira, G.L.V. Detection of Alterations in the Gut Microbiota and Intestinal Permeability in Patients With Hashimoto Thyroiditis. Front. Immunol. 2021, 12, 579140. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E.; Wahl, R. Microbiota and Thyroid Interaction in Health and Disease. Trends Endocrinol. Metab. 2019, 30, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Chichova, M.; Tasinov, O.; Shkodrova, M.; Mishonova, M.; Sazdova, I.; Ilieva, B.; Doncheva-Stoimenova, D.; Kiselova-Kaneva, Y.; Raikova, N.; Uzunov, B.; et al. New Data on Cylindrospermopsin Toxicity. Toxins 2021, 13, 41. [Google Scholar] [CrossRef]
- Chen, J.; Bian, R.; Li, J.; Qiu, L.; Lu, B.; Ouyang, X. Chronic exposure to microcystin-LR reduces thyroid hormone levels by activating p38/MAPK and MEK/ERK signal pathway. Ecotoxicol Environ. Saf. 2019, 173, 142–148. [Google Scholar] [CrossRef]
- Chen, L.; Shi, T.; Wang, Y.T.; He, J.; Zhao, X.; Wang, Y.K.; Giesy, J.P.; Chen, F.; Chen, Y.; Tuo, X.; et al. Effects of acute exposure to microcystins on hypothalamic-pituitary-adrenal (HPA), -gonad (HPG) and -thyroid (HPT) axes of female rats. Sci. Total Environ. 2021, 778, 145196. [Google Scholar] [CrossRef]
- Błażewicz, A.; Wiśniewska, P.; Skórzyńska-Dziduszko, K. Selected Essential and Toxic Chemical Elements in Hypothyroidism-A Literature Review (2001–2021). Int. J. Mol. Sci. 2021, 22, 10147. [Google Scholar] [CrossRef]
- Baltaci, A.K.; Mogulkoc, R.; Baltaci, S.B. Review: The role of zinc in the endocrine system. Pak. J. Pharm. Sci. 2019, 32, 231–239. [Google Scholar]
- Kawicka, A.; Regulska-Ilow, B. Metabolic disorders and nutritional status in autoimmune thyroid diseases. Postepy Hig. Med. Dosw. 2015, 69, 80–90. [Google Scholar] [CrossRef]
- Kim, D. The Role of Vitamin D in Thyroid Diseases. Int. J. Mol. Sci. 2017, 18, 1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pałkowska-Goździk, E.; Lachowicz, K.; Rosołowska-Huszcz, D. Effects of Dietary Protein on Thyroid Axis Activity. Nutrients 2017, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azami, M.; Parizad, N.; Sayehmiri, K. Prevalence of hypothyroidism, hypoparathyroidism and thefrequency of regular chelation therapy in patients with thalassemia major in Iran: A systematic review and meta-analysis study. Iran. J. Ped. Hematol. Oncol. 2016, 6, 260–275. [Google Scholar]
- Cayir, A.; Doneray, H.; Kurt, N.; Orbak, Z.; Kaya, A.; Turan, M.I.; Yildirim, A. Thyroid Functions and Trace Elements in Pediatric Patients with Exogenous Obesity. Biol. Trace Elem. Res. 2014, 157, 95–100. [Google Scholar] [CrossRef]
- Memon, N.S.; Kazi, T.G.; Afridi, H.I.; Baig, J.A.; Sahito, O.M.; Baloch, S.; Waris, M. Correlation of Manganese with Thyroid Function in Females Having Hypo- and Hyperthyroid Disorders. Biol. Trace Elem. Res. 2015, 167, 165–171. [Google Scholar] [CrossRef]
- Stojsavljevic, A.; Rovcanin, B.; Jagodic, J.; Radojkovic, D.D.; Paunovic, I.; Gavrovic-Jankulovic, M.; Manojlovic, D. Significance of Arsenic and Lead in Hashimoto’s Thyroiditis Demonstrated on Thyroid Tissue, Blood, and Urine Samples. Environ. Res. 2020, 186, 109538. [Google Scholar] [CrossRef]
- Jain, R.B.; Choi, Y.S. Interacting effects of selected trace and toxic metals on thyroid function. Int. J. Environ. Health Res. 2016, 26, 75–91. [Google Scholar] [CrossRef]
- Benvenga, S.; Marini, H.R.; Micali, A.; Freni, J.; Pallio, G.; Irrera, N.; Squadrito, F.; Altavilla, D.; Antonelli, A.; Ferrari, S.M.; et al. Protective Effects of Myo-Inositol and Selenium on Cadmium-Induced Thyroid Toxicity in Mice. Nutrients 2020, 12, 1222. [Google Scholar] [CrossRef]
- Talebi, S.; Ghaedi, E.; Sadeghi, E.; Mohammadi, H.; Hadi, A.; Clark, C.C.T.; Askari, G. Trace Element Status and Hypothyroidism: A Systematic Review and Meta-Analysis. Biol. Trace Elem. Res. 2020, 197, 1–14. [Google Scholar] [CrossRef]
- Igbokwe, I.O.; Igwenagu, E.; Igbokwe, N.A. Aluminium toxicosis: A review of toxic actions and effects. Interdiscip. Toxicol. 2019, 12, 45–70. [Google Scholar] [CrossRef] [Green Version]
- Genchi, G.; Carocci, A.; Lauria, G.; Sinicropi, M.S.; Catalano, A. Nickel: Human Health and Environmental Toxicology. Int. J. Environ. Res. Public Health 2020, 17, 679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopp, E.; Rettenmeier, A.W. Tin, Toxicity. In Encyclopedia of Metalloproteins; Kretsinger, R.H., Uversky, V.N., Permyakov, E.A., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Chitambar, C.R. Medical Applications and Toxicities of Gallium Compounds. Int. J. Environ. Res. Public Health 2010, 7, 2337–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitto, A.; Polito, F.; Atteritano, M.; Altavilla, D.; Mazzaferro, S.; Marini, H.; Adamo, E.B.; D’Anna, R.; Granese, R.; Corrado, F.; et al. Genistein aglycone does not affect thyroid function: Results from a three-year, randomized, double-blind, placebo-controlled trial. J. Clin. Endocrinol. Metab. 2010, 95, 3067–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, H.; Polito, F.; Adamo, E.B.; Bitto, A.; Squadrito, F.; Benvenga, S. Update on genistein and thyroid: An overall message of safety. Front. Endocrinol. 2012, 3, 94. [Google Scholar] [CrossRef] [Green Version]
- Meriluoto, J.; Spoof, L.; Codd, G.A. (Eds.) Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons: Chichester, UK, 2017. [Google Scholar]
- Mills, M.C.; Evans, M.V.; Lee, S.; Knobloch, T.; Weghorst, C.; Lee, J. Acute cyanotoxin poisoning reveals a marginal effect on mouse gut microbiome composition but indicates metabolic shifts related to liver and gut inflammation. Ecotoxicol. Environ. Saf. 2021, 215, 112126. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 31. [Google Scholar] [CrossRef] [Green Version]
- Arman, T.; Clarke, J.D. Microcystin Toxicokinetics, Molecular Toxicology, and Pathophysiology in Preclinical Rodent Models and Humans. Toxins 2021, 13, 537. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Chen, G.; Wang, M.; Hu, T. Cylindrospermopsin impairs vascular smooth muscle cells by P53-mediated apoptosis due to ROS overproduction. Toxicol. Lett. 2021, 353, 83–92. [Google Scholar] [CrossRef]
- Sazdova, I.; Keremidarska-Markova, M.; Chichova, M.; Uzunov, B.; Georgiev, G.; Mladenov, M.; Schubert, R.; Stoyneva-Gärtner, M.; Gagov, H.S. Review of Cyanotoxicity Studies Based on Cell Cultures. J. Toxicol. 2022, 2022, 5647178. [Google Scholar] [CrossRef]
- Passali, M.; Josefsen, K.; Frederiksen, J.L.; Antvorskov, J.C. Current Evidence on the Efficacy of Gluten-Free Diets in Multiple Sclerosis, Psoriasis, Type 1 Diabetes and Autoimmune Thyroid Diseases. Nutrients 2020, 12, 2316. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.; Ross, V.; Mahadevan, U. Popular exclusionary diets for inflammatory bowel disease: The search for a dietary culprit. Inflamm. Bowel Dis. 2014, 20, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Walters, S.S.; Quiros, A.; Rolston, M.; Grishina, I.; Li, J.; Fenton, A.; DeSantis, T.Z.; Thai, A.; Andersen, G.L.; Papathakis, P.; et al. Analysis of Gut Microbiome and Diet Modification in Patients with Crohn’s Disease. SOJ Microbiol. Infect. Dis. 2014, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olendzki, B.C.; Silverstein, T.D.; Persuitte, G.M.; Ma, Y.; Baldwin, K.R.; Cave, D. An anti-inflammatory diet as treatment for inflammatory bowel disease: A case series report. Nutr. J. 2014, 13, 5. [Google Scholar] [CrossRef] [Green Version]
- Konijeti, G.G.; Kim, N.; Lewis, J.D.; Groven, S.; Chandrasekaran, A.; Grandhe, S.; Diamant, C.; Singh, E.; Oliveira, G.; Wang, X.; et al. Efficacy of the Autoimmune Protocol Diet for Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 2054–2060. [Google Scholar] [CrossRef]
- Maes, M.; Leunis, J.C. Normalization of leaky gut in chronic fatigue syndrome (CFS) is accompanied by a clinical improvement: Effects of age, duration of illness and the translocation of LPS from gram-negative bacteria. Neuroendocrinol. Lett. 2008, 29, 902–910. [Google Scholar]
- Sen, T.; Cawthon, C.R.; Ihde, B.T.; Hajnal, A.; DiLorenzo, P.M.; de La Serre, C.B.; Czaja, K. Diet-driven microbiota dysbiosis is associated with vagal remodeling and obesity. Physiol. Behav. 2017, 173, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Inglis, J.E.; Lin, P.J.; Kerns, S.L.; Kleckner, I.R.; Kleckner, A.S.; Castillo, D.A.; Mustian, K.M.; Peppone, L.J. Nutritional Interventions for Treating Cancer-Related Fatigue: A Qualitative Review. Nutr. Cancer 2019, 71, 21–40. [Google Scholar] [CrossRef]
- Lankinen, M.; Uusitupa, M.; Schwab, U. Nordic Diet and Inflammation-A Review of Observational and Intervention Studies. Nutrients 2019, 11, 1369. [Google Scholar] [CrossRef] [Green Version]
- Kharrazian, D.; Herbert, M.; Vojdani, A. Immunological Reactivity Using Monoclonal and Polyclonal Antibodies of Autoimmune Thyroid Target Sites with Dietary Proteins. J. Thyroid Res. 2017, 2017, 4354723. [Google Scholar] [CrossRef]
- Hargreaves, C.E.; Grasso, M.; Hampe, C.S.; Stenkova, A.; Atkinson, S.; Joshua, G.W.; Wren, B.W.; Buckle, A.M.; Dunn-Walters, D.; Banga, J.P. Yersinia enterocolitica provides the link between thyroid-stimulating antibodies and their germline counterparts in Graves’ disease. J. Immunol. 2013, 190, 5373–5381. [Google Scholar] [CrossRef] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet—Microbe—Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haß, U.; Herpich, C.; Norman, K. Anti-Inflammatory Diets and Fatigue. Nutrients 2019, 11, 2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Kim, W.-L.; Park, B.-H.; Lee, S.-O.; Chae, S.-W. Effect of toxic trace element detoxification, body fat reduction following four-week intake of the Wellnessup diet: A three-arm, randomized clinical trial. Nutr. Metab. 2020, 17, 47. [Google Scholar] [CrossRef]
- Altobelli, E.; Del Negro, V.; Angeletti, P.M.; Latella, G. Low-FODMAP Diet Improves Irritable Bowel Syndrome Symptoms: A Meta-Analysis. Nutrients 2017, 9, 940. [Google Scholar] [CrossRef]
- Rakel, D. The Elimination Diet. In Integrative Medicine, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 86. [Google Scholar]
- Wang, K.; Wei, H.; Zhang, W.; Li, Z.; Ding, L.; Yu, T.; Tan, L.; Liu, Y.; Liu, T.; Wang, H.; et al. Severely low serum magnesium is associated with increased risks of positive anti- thyroglobulin antibody and hypothyroidism: A cross-sectional study. Sci. Rep. 2018, 8, 9904. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.D.; Chen, C.Y.; Knox, N.C.; Marrie, R.A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Van Domselaar, G. A comparative study of the gut microbiota in immune-mediated inflammatory diseases-does a common dysbiosis exist? Microbiome 2018, 6, 221. [Google Scholar] [CrossRef]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef]
- Salazar-Viedma, M.; Vergaño-Salazar, J.G.; Pastenes, L.; D’Afonseca, V. Simulation Model for Hashimoto Autoimmune Thyroiditis Disease. Endocrinology 2021, 162, bqab190. [Google Scholar] [CrossRef]
- Bruun, J.M.; Maersk, M.; Belza, A.; Astrup, A.; Richelsen, B. Consumption of sucrose-sweetened soft drinks increases plasma levels of uric acid in overweight and obese subjects: A 6-month randomised controlled trial. Eur. J. Clin. Nutr. 2015, 69, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Garcia, E.; Schulze, M.B.; Meigs, J.B.; Manson, J.E.; Rifai, N.; Stampfer, M.J.; Willett, W.C.; Hu, F.B. Consumption of trans fatty acids is related to plasma biomarkers of inflammation and endothelial dysfunction. J. Nutr. 2005, 135, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.J.; Kabi, A.; Nickerson, K.P.; McDonald, C. Combinatorial effects of diet and genetics on inflammatory bowel disease pathogenesis. Inflamm. Bowel Dis. 2015, 21, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Hammerling, U.; Bergman Laurila, J.; Grafström, R.; Ilbäck, N.G. Consumption of Red/Processed Meat and Colorectal Carcinoma: Possible Mechanisms Underlying the Significant Association. Crit. Rev. Food Sci. Nutr. 2016, 56, 614–634. [Google Scholar] [CrossRef]
- Oliveira, A.; Rodríguez-Artalejo, F.; Lopes, C. Alcohol intake and systemic markers of inflammation—Shape of the association according to sex and body mass index. Alcohol Alcohol. 2010, 45, 119–125. [Google Scholar] [CrossRef]
- Papier, K.; Hartman, L.; Tong, T.; Key, T.J.; Knuppel, A. Higher Meat Intake Is Associated with Higher Inflammatory Markers, Mostly Due to Adiposity: Results from UK Biobank. J. Nutr. 2022, 152, 183–189. [Google Scholar] [CrossRef]
- Chai, W.; Morimoto, Y.; Cooney, R.V.; Franke, A.A.; Shvetsov, Y.B.; Le Marchand, L.; Haiman, C.A.; Kolonel, L.N.; Goodman, M.T.; Maskarinec, G. Dietary Red and Processed Meat Intake and Markers of Adiposity and Inflammation: The Multiethnic Cohort Study. J. Am. Coll. Nutr. 2017, 36, 378–385. [Google Scholar] [CrossRef]
- Knezevic, J.; Starchl, C.; Tmava Berisha, A.; Amrein, K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020, 12, 1769. [Google Scholar] [CrossRef]
- Virili, C.; Fallahi, P.; Antonelli, A.; Benvenga, S.; Centanni, M. Gut microbiota and Hashimoto’s thyroiditis. Rev. Endocr. Metab. Disord. 2018, 19, 293–300. [Google Scholar] [CrossRef]
- Docimo, G.; Cangiano, A.; Romano, R.M.; Pignatelli, M.F.; Offi, C.; Paglionico, V.A.; Galdiero, M.; Donnarumma, G.; Nigro, V.; Esposito, D.; et al. The Human Microbiota in Endocrinology: Implications for Pathophysiology, Treatment, and Prognosis in Thyroid Diseases. Front. Endocrinol. 2020, 11, 586529. [Google Scholar] [CrossRef]
- Golan, D.; Staun-Ram, E.; Glass-Marmor, L.; Lavi, I.; Rozenberg, O.; Dishon, S.; Barak, M.; Ish-Shalom, S.; Miller, A. The influence of vitamin D supplementation on melatonin status in patients with multiple sclerosis. Brain Behav. Immun. 2013, 32, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Maseeh, A. Vitamin D: The “sunshine” vitamin. J. Pharm. Pharmacother. 2012, 3, 118–126. [Google Scholar] [CrossRef]
- Dupuis, M.L.; Pagano, M.T.; Pierdominici, M.; Ortona, E. The role of vitamin D in autoimmune diseases: Could sex make the difference? Biol. Sex Differ. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Huiberts, L.M.; Smolders, C.H.J.K. Effects of vitamin D on mood and sleep in the healthy population: Interpretations from the serotonergic pathway. Sleep Med. Rev. 2021, 55, 101379. [Google Scholar] [CrossRef]
- Ghareghani, M.; Reiter, R.J.; Zibara, K.; Farhadi, N. Latitude, Vitamin D, Melatonin, and Gut Microbiota Act in Concert to Initiate Multiple Sclerosis: A New Mechanistic Pathway. Front. Immunol. 2018, 9, 2484. [Google Scholar] [CrossRef]
- Sears, B. Anti-inflammatory Diets. J. Am. Coll. Nutr. 2015, 34, 14–21. [Google Scholar] [CrossRef]
- Caesar, R.; Tremaroli, V.; Kovatcheva-Datchary, P.; Cani, P.D.; Bäckhed, F. Crosstalk between Gut Microbiota and Dietary Lipids Aggravates WAT Inflammation through TLR Signaling. Cell Metab. 2015, 22, 658–668. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, C.; Bucci, I.; Di Santo, S.; Rossi, C.; Grassadonia, A.; Mariotti, M.; Piantelli, M.; Monaco, F.; Napolitano, G. Resveratrol inhibits sodium/iodide symporter gene expression and function in rat thyroid cells. PLoS ONE 2014, 9, e107936. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive Compounds and Bioactivities of Ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Berbert, A.A.; Kondo, C.R.; Almendra, C.L.; Matsuo, T.; Dichi, I. Supplementation of fish oil and olive oil in patients with rheumatoid arthritis. Nutrition 2005, 21, 131–136. [Google Scholar] [CrossRef]
- Thornley, S.; Tayler, R.; Sikaris, K. Sugar restriction: The evidence for a drug-free intervention to reduce cardiovascular disease risk. Intern. Med. J. 2012, 5, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Eliaz, I.; Raz, A. Pleiotropic Effects of Modified Citrus Pectin. Nutrients 2019, 11, 2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candelli, M.; Franza, L.; Pignataro, G.; Ojetti, V.; Covino, M.; Piccioni, A.; Gasbarrini, A.; Franceschi, F. Interaction between Lipopolysaccharide and Gut Microbiota in Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 6242. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.; Morley, J.E.; Hershman, J.M. Melatonin and the thyroid. Horm. Metab. Res. 1980, 12, 71–73. [Google Scholar] [CrossRef]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin Inhibits Oxidative Stress and Apoptosis in Cryopreserved Ovarian Tissues via Nrf2/HO-1 Signaling Pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef]
- Bellipanni, G.; Bianchi, P.; Pierpaoli, W.; Bulian, D.; Ilyia, E. Effects of melatonin in perimenopausal and menopausal women: A randomized and placebo controlled study. Exp. Gerontol. 2001, 36, 297–310. [Google Scholar] [CrossRef]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The potential benefits of red beetroot supplementation in health and disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef]
- Guan, Y.S.; He, Q. Plants Consumption and Liver Health. Evid Based Complement. Altern. Med. 2015, 2015, 824185. [Google Scholar] [CrossRef] [Green Version]
- Klimek-Szczykutowicz, M.; Szopa, A.; Ekiert, H. Citrus limon (Lemon) Phenomenon-A Review of the Chemistry, Pharmacological Properties, Applications in the Modern Pharmaceutical, Food, and Cosmetics Industries, and Biotechnological Studies. Plants 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.; Neerajand, S.; Kumar, J. Functional Properties of Pomegranate (Punica granatum L.). Pharm. Innov. 2018, 7, 71–81. Available online: https://www.thepharmajournal.com/archives/2018/vol7issue10/PartB/7-9-41-419.pdf (accessed on 28 March 2022).
- Kawagishi, H.; Fukumoto, Y.; Hatakeyama, M.; He, P.; Arimoto, H.; Matsuzawa, T.; Arimoto, Y.; Suganuma, H.; Inakuma, T.; Sugiyama, K. Liver injury suppressing compounds from avocado (Persea americana). J. Agric. Food Chem. 2001, 49, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, D.J.; Alsherbiny, M.A.; Perera, S.; Low, M.; Basu, A.; Devi, O.A.; Barooah, M.S.; Li, C.G.; Papoutsis, K. The Odyssey of Bioactive Compounds in Avocado (Persea americana) and Their Health Benefits. Antioxidants 2019, 8, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliaz, I.; Hotchkiss, A.T.; Fishman, M.L.; Rode, D. The effect of modified citrus pectin on urinary excretion of toxic elements. Phytother. Res. 2006, 20, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Aggarwal, B.B. Turmeric, the Golden Spice: From Traditional Medicine to Modern Medicine. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011; Chapter 13. [Google Scholar]
- McAnulty, L.S.; Nieman, D.C.; Dumke, C.L.; Shooter, L.A.; Henson, D.A.; Utter, A.C.; Milne, G.; McAnulty, S.R. Effect of blueberry ingestion on natural killer cell counts, oxidative stress, and inflammation prior to and after 2.5 h of running. Appl. Physiol. Nutr Metab. 2011, 36, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Sears, M.E. Chelation: Harnessing and enhancing heavy metal detoxification--a review. Sci. World J. 2013, 18, 219840. [Google Scholar] [CrossRef] [Green Version]
- Mashhadi, N.S.; Ghiasvand, R.; Askari, G.; Hariri, M.; Darvishi, L.; Mofid, M.R. Anti-oxidative and anti-inflammatory effects of ginger in health and physical activity: Review of current evidence. Int. J. Prev. Med. 2013, 1, S36–S42. [Google Scholar]
- Brent, G.A. Environmental exposures and autoimmune thyroid disease. Thyroid 2010, 20, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, Y.M.L.; Silva, W.E.D.; Souza, F.A.L.; Teixeira, V.W.; Teixeira, Á.A.C. Melatonin on hypothyroidism and gonadal development in rats: A review. JBRA Assist. Report. 2020, 24, 498–506. [Google Scholar] [CrossRef]
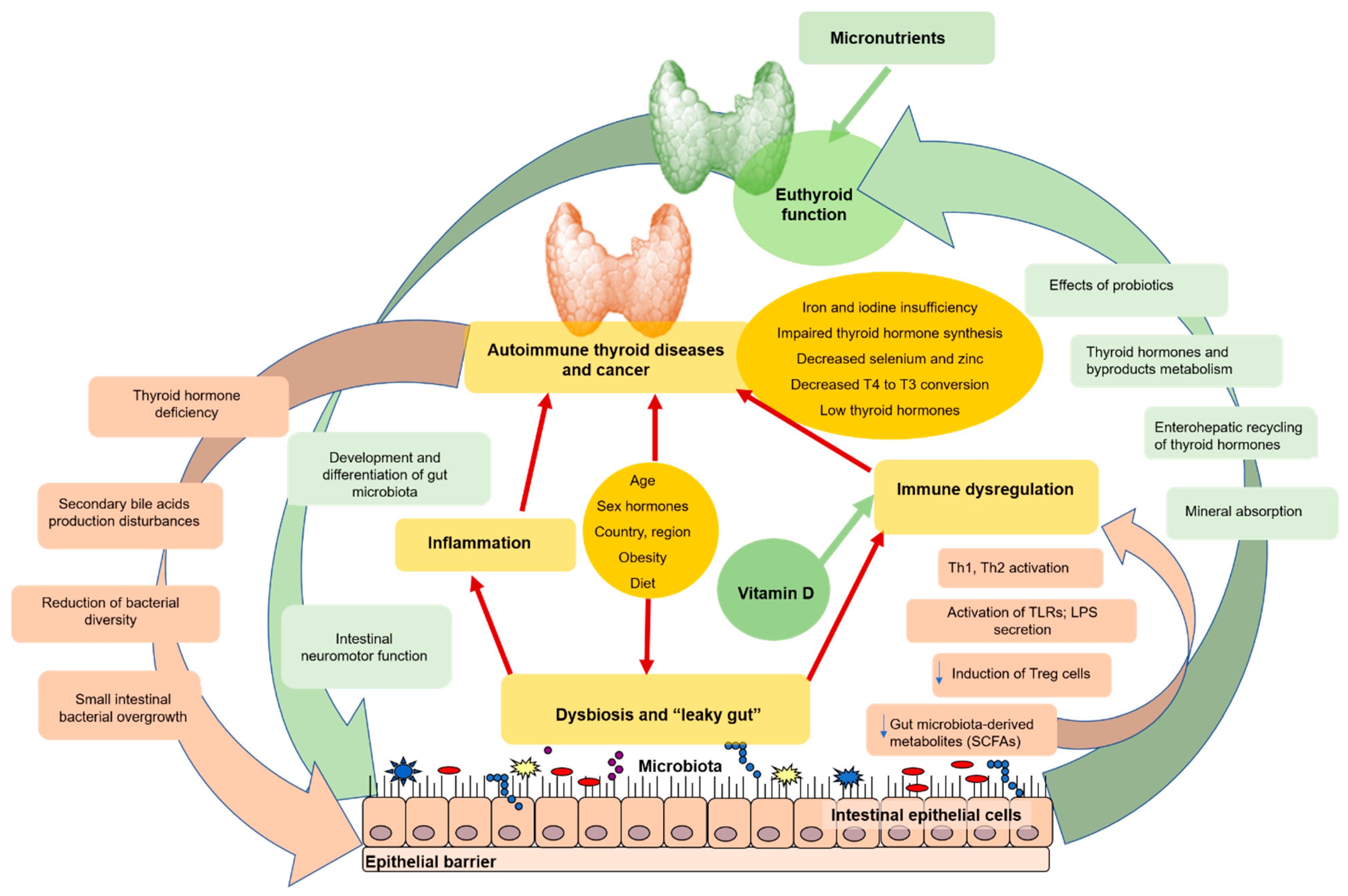

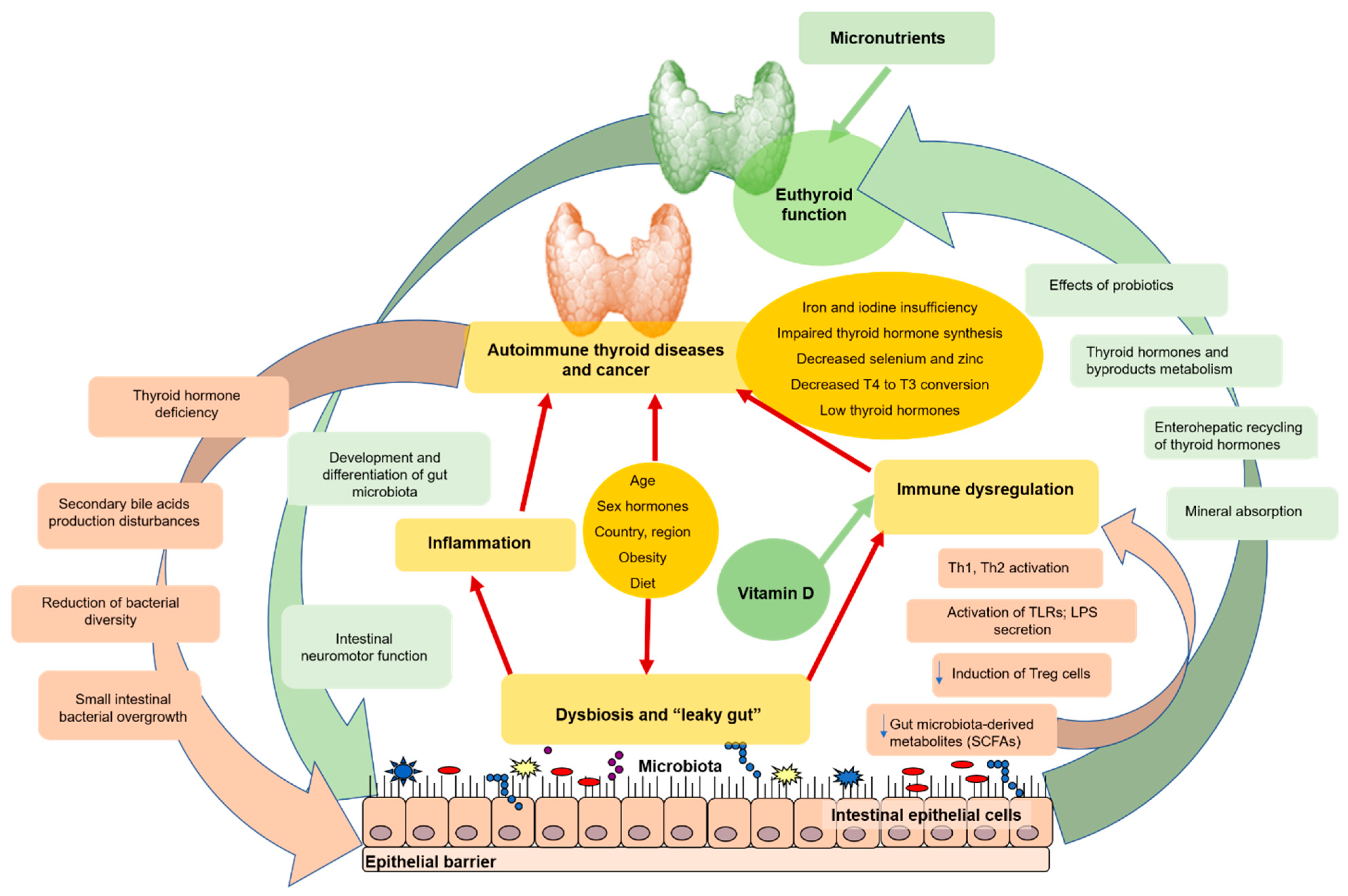{kind=link}
| Nutrients | Foods | Effects | Molecular Targets | Restrictions | Ref. |
|---|---|---|---|---|---|
| Iodine/I | Iodized salt, plum, maize; sea fish, iodized milk, dairy products, chicken eggs | Deficiency of iodine may cause goiter and hypothyroidism. Thyroid peroxidase in the presence of peroxide iodinates tyrosine bound to thyroglobulin | Part of T3 and T4 | Chronic high dietary iodine intake may induce autoimmune thyroiditis | [1,9] |
| Iron/Fe | Meat, animal offal, pumpkin seeds, cocoa and bitter chocolate, sardines, seafood | Necessary for the thyroid peroxidase reaction | Thyroid peroxidase | Chronic high dietary iron has pro-oxidant and cancerogenic effects | [4] |
| Selenium/Se | Brazilian walnut, meat, liver, fish, spinach | Activation or inactivation of T4, antioxidant and anti-inflammatory effects | Thyroid function depends on glutathione peroxidases, deiodinases, and selenoprotein S | High doses of Se are toxic | [4,9] |
| Zinc/Zn | Whole grain cereals, flax seeds, pumpkin seeds, millet, meat, buckwheat | Zn deficiency leads to disturbances in T3 and T4 levels and increases antibody titers against thyroid antigens | Cofactor for over 300 metalloproteins activates deiodinases and is needed for proper T3 receptor signaling, stimulates the synthesis of TBS | Low doses of Zn cause oxidative stress | [17,19] |
| Vitamin D | Fish oil, fatty fish, chicken eggs | Malnutrition of vitamin D correlates to autoimmune diseases | Nuclear vitamin D receptor via vitamin D response element regulates more than 200 human genes | Hypervitaminosis leads to hypercalcemia | [4,21] |
| Proteins | From unprocessed meat, eggs, sea fish | Low-protein content, soy proteins diet, and starvation downregulate HPT axis; malnutrition leads to thyroid gland damage, especially in children | Low-protein diet increases plasma TBG and decreases plasma transthyretin, T3, and pituitary TSH transcript in rats, and increases TSH in humans | N/A | [20,22] |
| Inositol (vitamin B8) | Citrus fruit, cantaloupe, bananas, raisins, and fiber-rich foods | TSH signaling, thyroid cells’ protection; immunomodulatory effect, decreases TSH to normal values in patients with hypothyroidism when applied with selenium | In TSH signaling as part of PIP-3; T3 decreases TPO-Ab, and Tg-Ab increases thyrocytes viability in the presence of H2O2 and cytokines | [10] |
| Nutrients | Foods | Effects | Molecular Targets | Restrictions | Ref. |
|---|---|---|---|---|---|
| Manganese/Mn | Whole grains, clams, oysters, mussels, nuts, soybeans, leafy vegetables and legumes, rice, coffee, tea, black pepper, and other spices | Mn modulates TSH secretion by a dopaminergic mechanism | Part of Mn-SOD | In high doses, pro-antioxidative effect | [24,25] |
| Fluoride/F | Drinking water, fluoride-treated salt | Higher fluoride in drinking water increases hypothyroidism by about twice, iodine deficiency | T3 and T4 iodination | Toxic | [18] |
| Lead/Pb | Polluted air | Increased plasma levels in hypothyroidism, pro-oxidative effects; decreases Se in blood | Thyroid selenoproteins | Toxic | [26] |
| Cadmium/Cd | Smelters, food, burning fossil fuels, plastics, and nickel-cadmium batteries, cigarette smoke, phosphate fertilizers | Oxidative stress and mitochondrial, leading to hypothyroidism and hyperthyroidism; selenium and myo-inositol protect against Cd | MCP-1 and C-X-C motif chemokine 10 expression | Toxic | [18,27,28] |
| Chromium/Cr | High doses from air, foods, or through the skin | Via pleiotropic mechanisms, most of them indirect via insulin, cortisol, Fe, and Se | Oxidation of proteins influences Fe and Se homeostasis | Oxidative changes of proteins | [29] |
| Iron/Fe | Red meat, liver, beans, edamame beans, chickpeas, nuts, dried fruit (apricots), cereals, soybean flour | Facilitates thyroid hormone iodination | Activates thyroid peroxidase | In high doses, pro-antioxidant | [23] |
| Aluminum/Al | Through food, through breathing, and by skin contact | The aluminum ion (Al3+) is harmful. The uptake of aluminum can occur through food, breathing, and skin contact. Long-lasting uptakes of significant aluminum concentrations can damage the central nervous system, leading to dementia, loss of memory, listlessness, severe trembling | AlF4- is a non-specific G-protein activator | In high doses, neurotoxin | [30] |
| Nickel/Ni | Hazelnuts; cocoa and dark chocolate; fruits (almonds, dates, figs, pineapple, plums, raspberries); grains (bran, buckwheat, millet, whole grain bread, oats, brown rice, sesame seeds, sunflower seeds); seafood (shrimps, mussels, oysters, crab, salmon); vegetables (beans, savoy cabbage, leeks, lettuce, lentils, peas, spinach, cabbage); tea from drink dispensers; soya and soya products; peanuts; licorice; baking powder | Contact dermatitis; headaches; gastrointestinal manifestations; respiratory manifestations; lung fibrosis; cardiovascular diseases; lung cancer; nasal cancer; epigenetic effects | Immunotoxic and carcinogenic agent | N/A | [31] |
| Tin/Sn | Tin is present in the air, water, soil, and landfills; it is a normal part of many plants and animals; tin concentrations in foods not packaged in metal cans are minimal; people can be exposed to the tin when consuming food or liquid from tin-lined cans. | Inhalation, oral, or dermal exposure to some organotin compounds has been shown to cause harmful effects in human skin and eye irritation, respiratory irritation, gastrointestinal effects, and neurological problems in humans exposed for a short period to high amounts of organotin compounds. | N/A | Lethal intoxication cases may appear with large amounts but are rare | [32] |
| Gallium/Ga | Found in small amounts in nature and the human body | Acute exposure to gallium (III) chloride can cause throat irritation, difficulty breathing, and chest pain. Its fumes can cause pulmonary edema and partial paralysis. | N/A | N/A | [33] |
| Genistein | Soy and soy foods contain this phytoestrogen belonging to isoflavones | Goitrogenic effect, hypothyroidism | Inhibitor of thyroid peroxidase and sulfotransferase enzymes | Long-term consumption presents a risk for infants and women | [34,35] |
| Diet | Foods | Eliminated Foods or Nutrients | Effects | Restrictions or Remarks | Ref. |
|---|---|---|---|---|---|
| Low-residue diet | Elimination of fruits, vegetables, whole grains, and legumes | Reduced fiber intake | Relieve obstructive symptoms; no effect on inflammation | Long-term use could decrease microbial diversity | [11,44,45] |
| Anti-inflammatory diet | More prebiotic and probiotic foods, n-3 PUFA, wild fish, grass-fed meat, vegetables, fruit, nuts, some saturated fat | Restriction of gluten, lactose, total fat, refined carbohydrates, others | Decrease of HBI or MTLWSI | N/A | [46] |
| Autoimmune diet (modified-Paleo diet) | Meat (grass-fed), fish, vegetables, excluding nightshade vegetables, sweet potatoes, fruit (in small amounts), coconut milk, avocado, olive, coconut oil, dairy-free fermented foods (kefir, kombucha, sauerkraut, kimchi) | Less or elimination of processed food, dairy, grains, refined sugars, legumes, and cereals | Remission of gut inflammation | N/A | [47] |
| Leaky gut diet | Glutamine, N-acetyl-L-cysteine, and zinc | Gluten- and milk-free, low-carb, and low-sugar diet | Reduces gut-derived inflammation; ROS and inflammatory cytokines IL-1β, IL-6, IFN-γ, TNF-α; prevents bacterial translocation via enhancing weakened tight junctions | N/A | [48,49] |
| Mediterranean diet | Fish, lean meat, whole grains, legumes, nuts, dairy, olive oil, vegetables, fruits, and moderate wine consumption | Heavily processed foods, i.e., processed red meats, refined grains, refined/processed/hydrogenated oils, alcohol, butter | Lowers the inflammatory load and simultaneously balances gut microbiota | Diet is rich in omega-3 and omega-9 FA, fibers, complex carbohydrates, minerals, vitamins, and secondary plant metabolites | [50] |
| Nordic diet | High consumption of whole grains and unrefined sugars, fish and lean meat, dairy products, canola or rapeseed oil, and vegetables and fruits like cabbage and berries | Refined carbohydrates and processed foods | Lower blood pressure, normalize cholesterol, lose weight, or maintain a healthy weight. | Causing obesity | [51] |
| Proteins (dietary) | Latex hevein, kamut, soy sauce, gelatin, scallops, cashew, Brazil nut | Reduction of other macronutrients | Cross-reactivity of dietary proteins with monoclonal antibodies against T4, T3, and Tg | The observed immunoreactivity of purified dietary proteins in vitro might have an antithyroid effect in vivo only in the very leaky intestine | [52] |
| Proteins (microbial) | Protein fermented food | N/A | Gastrointestinal pathogen Yersinia enterocolitica have proteins (porins) that mimic thyroid antigens and could lead to autoimmunization and stimulate precursor B cells for TSHR-Ab production | N/A | [53,54] |
| Fatigue reduction and anti-inflammatory diet | Foods rich in antioxidative vitamins, omega-3 fatty acids, and in fibers, polyphenol-rich vegetables | Reduced inflammatory foods | Anti-inflammatory effect, fatigue reduction | N/A | [55] |
| Western diets | Rich in linoleic acid; high ratio of ω-6 to ω-3 FA | Diet is rich in calories | Inflammatory effect | N/A | [56] |
| Wellnessup diet | Organic plant-based diet including various vegetables, fruits, whole grains, nuts, and phytonutrients | Elimination of meat, eggs, fish, dairy products, processed food, refined sugars | It may have several beneficial effects, such as body fat reduction and improving some of the detoxification elements through caloric restriction. | N/A | [57] |
| FODMAP diet | Proteins: beef, chicken, eggs, fish, lamb, pork, prawns, tempeh, and tofu; whole grains starches: white and brown rice, lentils, corn, oats, quinoa, cassava, and potatoes fruits: blueberries, raspberries, pineapple, honeydew melon, cantaloupe, kiwi, limes, guava, starfruit, grapes vegetables: bean sprouts, bell peppers, radishes, bok choy, carrots, celery, eggplant, tomatoes, spinach, cucumber, pumpkin, zucchini nuts: almonds, macadamia nuts, peanuts, pecans, pine nuts, and walnuts seeds: pumpkin, sesame, sunflower seeds, linseeds, dairy, coconut and olive oils, peppermint tea | Oligosaccharides: wheat, rye, nuts, legumes, garlic, artichokes, onion disaccharides: lactose-containing products—milk, ice cream, yogurt, soft cheese, buttermilk, condensed milk, whipped cream monosaccharides: fructose-containing fruits—apples, pears, mango, watermelon, sweeteners—honey, agave nectar, and high-fructose corn syrup polyols: mannitol and sorbitol in apples, pears, cauliflower, stone fruits, mushrooms, and snow peas; xylitol and isomalt in sweeteners, such as those in sugar-free gum and mints | Reduces symptoms of irritable bowel syndrome. May decrease both stomach pain and bloating. Helps manage flatulence, diarrhea, and constipation | N/A | [58] |
| Elimination diet | Common foods in the elimination diet are gluten, dairy products, citrus, soy, peanuts, eggs, corn, tree nuts, beef, refined sugars | Eliminates certain food or group of foods believed to cause an adverse food reaction, often referred to as a “food intolerance.” | May reduce inflammation and allergy symptoms | N/A | [58] |
| Vegan diet | Plant-based foods: fruits, vegetables, soy, legumes, nuts and nut butter, sprouted or fermented plant foods, plant-based dairy alternatives, and whole grains | Excludes meat, poultry, eggs, dairy, and seafood | Associated with improved glycemic control, lower total cholesterol, blood pressure, and BMI. May prevent cancer. | Possible deficiency of some essential amino acids and vitamins | [59] |
| Nutrients | Foods | Effects | Molecular Targets | Ref. |
|---|---|---|---|---|
| Inflammatory proteins | Avocado, orange juice, cooked Brussels sprouts, seaweed, radish, almond, Brazil nut, macadamia nut, mustard seeds, cashew, and others | Immunological reactivity of several dietary proteins to T3-Ab and T4-Ab, one with Tg and 5’ deiodinase. This could be observed in vivo only in case of a combination of improper protein digestion and leaky intestine | Specific T3, T4, Tg, and 5’ deiodinase Ag-Ab immune cross-reactivity for few dietary proteins with similarity to T3 or T4; no binding to TSH-R, TPO, or TBP; inflammatory effect only in impaired digestion and leaky intestine | [52] |
| Gluten | Wheat, rye, barley, and contaminated oat products | Antigen in CD; molecular mimicry with thyroid Tg; effect on gut microbiota and permeability; element deficiency due to CD; gluten-free diet for TH in the absence of CD is not recommended | Anti-Tg Ab can be produced in the comorbidity of CD and TH | [4] |
| Sugar: sucrose, fructose, glucose, high-fructose corn syrup (HFCS) | Refined sugar, candy, chocolate, cookies, sweets, cakes, foods with added sugar, sugary and soft drinks | Increase inflammatory markers and circulating uric acid; associated with metabolic disorders | [64] | |
| Artificial trans fatty acids (FA) | Hydrogenated vegetable oils, margarine | Increases production of inflammatory cytokines associated with atherosclerosis; increases the concentration of plasma biomarkers of inflammation and endothelial dysfunction | Trans FA increase plasma concentrations of CRP, IL-6, soluble TNF receptor 2, E-selectin, and sVCAM-1 | [65] |
| Refined carbohydrates | White bread, pasta, candy, pastries, cookies, cakes, sugary soft drinks, and all processed foods with added sugar or flour | Enhance the growth of inflammatory gut bacteria that can increase the risk of obesity and inflammatory bowel disease | [66] | |
| Advanced glycation end products | Red and processed meat such as grilled meat, sausage, bacon, ham, smoked meat, and beef jerky | Increased oxidative load and higher inflammatory response | Increased N-nitrosation and oxidative load leading to DNA adducts and lipid peroxidation in the intestinal epithelium, proliferative stimulation of the epithelium, higher inflammatory response, which may trigger pro-malignant processes | [67] |
| Ethanol | Alcoholic beverages such as wine, beer, spirits, liquors | Increased levels of C-reactive protein (CRP) may cause a “leaky gut” condition | N/A | [68] |
| Meat | Meat, both processed and unprocessed | Higher meat consumption, particularly of processed meat, was positively associated with the inflammatory marker—serum CRP (mg/L) | CRP, white blood cell count, interleukin 6 (IL-6), and TNF-α | [69,70] |
| Nutrients | Foods | Effects | Molecular Targets | Restrictions | Ref. |
|---|---|---|---|---|---|
| Magnesium/Mg | Yogurt, kefir, pumpkin seeds, cocoa, nuts, bitter chocolate, whole-grain cereal products, avocado, some fatty fish, green vegetables | Participates in many enzyme reactions, in melatonin synthesis, anti-inflammatory, deficit leads to immune disorders | Mg decreases C-reactive protein and antibodies against thyroglobulin (TG), part of Mg-SOD | N/A | [56,60] |
| Dietary fiber | Whole grains, beans, lentils, broccoli, apple, berries, dried fruits, avocado, popcorn | Support a proper gut microbiome, absorb toxins, anti-inflammatory; slows down and impedes nutrient uptake | N/A | N/A | [4] |
| Fatty acid | Fish, oils (flaxseed, extra-virgin olive oil, canola oil, sunflower oil), chia seeds, walnuts, avocados | Excess of saturated and deficiency of unsaturated fatty acids; provoke leaky intestinal syndrome and worse intestinal microbiota; induce cellular stress | Balance of dietary fatty acids improves gut microbiota, metabolism, and has an anti-inflammatory effect; saturated lipids induce inflammation via Toll-like receptor activation; n-3 PUFA decrease endotoxin permeability | N/A | [4,56,80] |
| Resveratrol | Vegetables and fruits, such as grapes, berries, and peanuts | Antioxidant, anti-inflammatory, and antiproliferative agent; short-term activation and long-term inhibition of expression and function of Na/I symporter | Na/I symporter | High doses of resveratrol will induce hypothyroidism | [81] |
| Gingerol, shogaol and paradols | Ginger (Zingiber officinale) roots | Phenolic substances with antioxidant and pleiotropic anti-inflammatory mechanisms regulating NO, PI3K/Akt, TNF-alpha signaling; antimicrobial and anti-cancer activity | Nrf2 enhances the expression of antioxidative enzymes, anti-inflammatory effects, glutathione, and stress-induced enzymes such as heme-oxygenase 1 | N/A | [82] |
| Omega-9 mono- unsaturated oleic FA | Olive and fish oil | Balancing gut microbiota and inflammatory response; omega-9 mono-unsaturated oleic FA, which can be converted to anti-inflammatory eicosatrienoic acid | Inhibits leukotriene B4 synthesis | N/A | [83] |
| Omega-3 | fish (salmon, mackerel, tuna, herring, and sardines), seafood, nuts and seeds (flaxseed, chia seeds, and walnuts), plant oils (flaxseed oil, soybean oil, and canola oil) | Regulates food intake, substrates for pro-resolving lipid mediators, anti-inflammatory effect | N/A | N/A | [56] |
| Sugars | candies, cakes, cookies, pies, sweet rolls, pastries, doughnuts, ice cream, sweetened yogurt, sweetened drinks, juice | Dysbiosis of gut microbiota increases gut inflammation, weight gain, and alters vagal gut–brain communication | N/A | Restrictions are needed to prevent from cardiovascular, metabolic, and other chronic diseases | [49,84] |
| Flavonoid and non-flavonoid dietary polyphenols | Plant sources—vegetables, fruits, dry legumes, cereals, olives, cocoa, coffee, tea, wine | Influence dendritic cells, have an immunomodulatory effect on macrophages, increase proliferation of B cells and T cells, and suppress Th1, Th2, Th17, and Th9 cells. Polyphenols reduce inflammation by suppressing the proinflammatory cytokines in inflammatory bowel disease by inducing Treg cells in the intestine, inhibiting TNF-α, inducing apoptosis, decreasing DNA damage. Role in prevention/treatment of autoimmune diseases | Downregulation of proinflammatory cytokines IL-1, IL-6, and INF-γ | N/A | [85] |
| Flavonoids | Plant sources—fruits, tea, vegetables, legumes, dark chocolate | Anti-inflammatory and antioxidant effect by free-radical scavengers and inhibition of their formation | Inhibition of protein kinases, phosphodiesterases, PLA2, COX, LOX, Modulation of NF-κB, GATA-3, and STAT-6 signaling | N/A | [85,86] |
| MCP | Dietary pectin is modified by pH and heat-controlled enzymatic treatment | MCP is absorbed in circulation; has anti-cancer effects; increases urinary excretion of lead, arsenic, and cadmium; reduces fibrosis in the liver, kidney, and adipose tissue; improves immune function | Antagonism of galectin-3 | N/A | [87] |
| Diet | Active Ingredients | Effects | Restrictions | Ref. |
|---|---|---|---|---|
| Beetroot (Beta vulgaris) | High levels of antioxidants betalain pigments and betaine | Amplify specific enzymes that support the liver and its detoxification, choleretic effect | Beet can make urine or stools appear pink or red. But this is not harmful. There is concern that beets might cause low calcium levels and kidney damage, but this has not been shown in people. | [92] |
| Broccoli (Brassica oleracea) | Diindolylmethane, glucoraphanin | Antiviral, anti-cancer | Gas or bowel irritation caused by broccoli’s high amounts of fiber. | [93] |
| Lemon (Citrus limon) | Naringin, citric acid | Decreasing liver damage, prevents oxidative (stress-related) damage. Regulate blood pressure. | Skin irritation is the most common side effect of using fruit acids. Lemon is extremely acidic, which can irritate the skin. | [94] |
| Pomegranate (Punica granatum) | Punicalagins (pomegranate ellagitannins) | Anti-cancer | Some people have experienced sensitivity to pomegranate extract; they include itching, swelling, runny nose, difficulty breathing | [95] |
| Watermelon (Citrullus lanatus) | Citrulline, lycopene | Antitoxic, hypoglycemic | Excessive use can lead to diarrhea and other digestive troubles. | [93] |
| Avocado (Persea americana) | Rich in fatty acids; vitamins; minerals, fiber, phytochemicals | Protect against damage caused by liver toxin d-galactosamine, contains antioxidants, anti-cancer, and anti-inflammatory components | Excess use may lead to excess calories intake. | [96,97] |
| Apple (Malus domestica) | Soluble fiber pectin | Helps purge toxins from the bloodstream and lower LDL cholesterol; aid the excretion of mercury and lead | No side effects of apple fruit or apple juice; apple seeds contain cyanide and are poisonous | [98] |
| Turmeric (Curcuma longa) | Curcumin | Antibacterial, antiviral, anti-inflammatory, antitumor, antioxidant, antiseptic, cardioprotective, hepatoprotective, nephroprotective, radioprotective, and digestive activities | Turmeric usually does not cause serious side effects. Some people can experience mild side effects such as stomach upset, nausea, dizziness, or diarrhea. These side effects are more common at higher doses. | [99] |
| Blueberries (Vaccinium angustifolium) | Antioxidants (anthocyanins) | Lower blood pressure, boost vascular health, protect lungs, anti-cancer effect, prevent Alzheimer’s disease. | Blueberry fruit is safe when consumed in normal amounts | [100] |
| Cilantro (Coriandrum sativum) | Metal-binding proteins | Enhance mercury excretion and decrease lead absorption. | Rarely people might experience allergies after eating cilantro, such as hives, facial or throat swelling | [101] |
| Ginger (Zingiber officinale) | Phytochemicals | Boost the body’s ability to process food and eliminate waste; protects against oxidative stress, anti-inflammatory, and anti-cancer effects | Higher doses of 5 g can cause mild side effects, including heartburn, diarrhea, burping, and general stomach discomfort | [102] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danailova, Y.; Velikova, T.; Nikolaev, G.; Mitova, Z.; Shinkov, A.; Gagov, H.; Konakchieva, R. Nutritional Management of Thyroiditis of Hashimoto. Int. J. Mol. Sci. 2022, 23, 5144. https://doi.org/10.3390/ijms23095144
Danailova Y, Velikova T, Nikolaev G, Mitova Z, Shinkov A, Gagov H, Konakchieva R. Nutritional Management of Thyroiditis of Hashimoto. International Journal of Molecular Sciences. 2022; 23(9):5144. https://doi.org/10.3390/ijms23095144
Chicago/Turabian StyleDanailova, Yana, Tsvetelina Velikova, Georgi Nikolaev, Zorka Mitova, Alexander Shinkov, Hristo Gagov, and Rossitza Konakchieva. 2022. "Nutritional Management of Thyroiditis of Hashimoto" International Journal of Molecular Sciences 23, no. 9: 5144. https://doi.org/10.3390/ijms23095144
APA StyleDanailova, Y., Velikova, T., Nikolaev, G., Mitova, Z., Shinkov, A., Gagov, H., & Konakchieva, R. (2022). Nutritional Management of Thyroiditis of Hashimoto. International Journal of Molecular Sciences, 23(9), 5144. https://doi.org/10.3390/ijms23095144