
Investigating the Short Peptidome Profile of Italian Dry-Cured Ham at Different Processing Times by High-Resolution Mass Spectrometry and Chemometrics

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
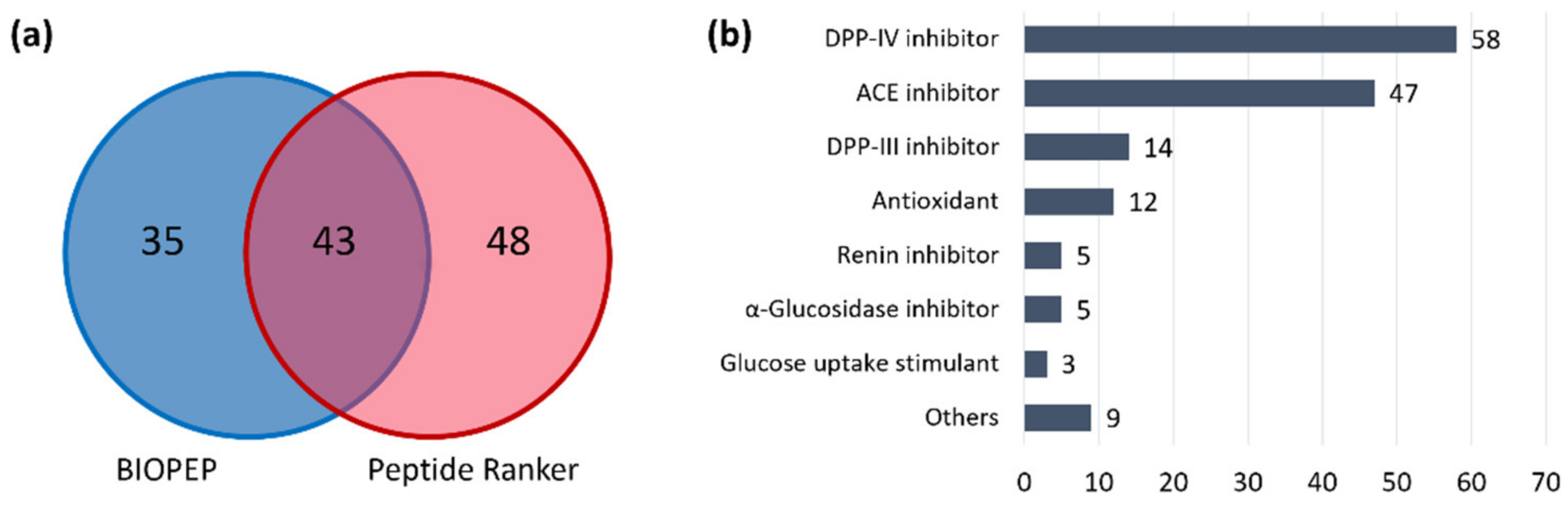
2.1. Natural Short Peptide Profile of Dry-Cured Ham
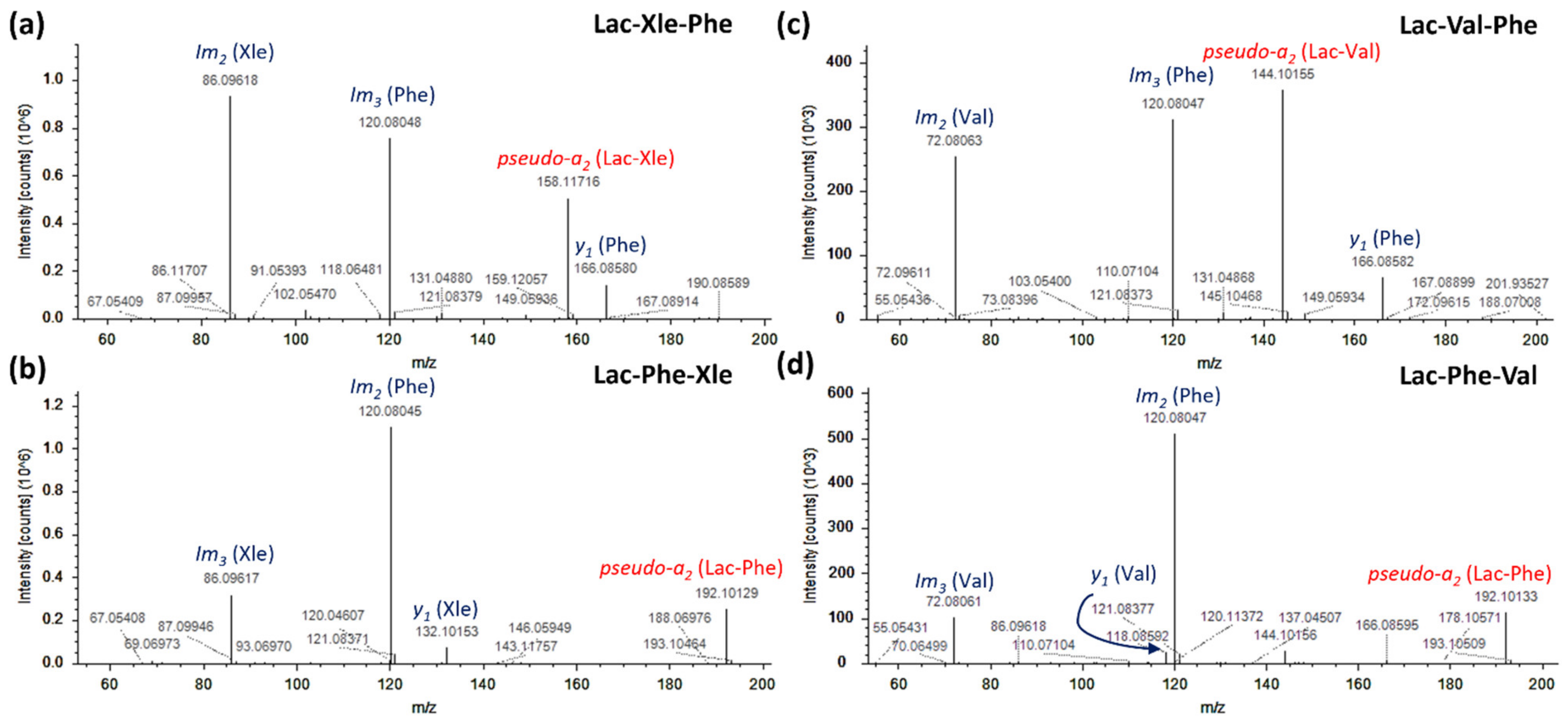
2.2. Modified Short Peptide Profile of Dry-Cured Ham
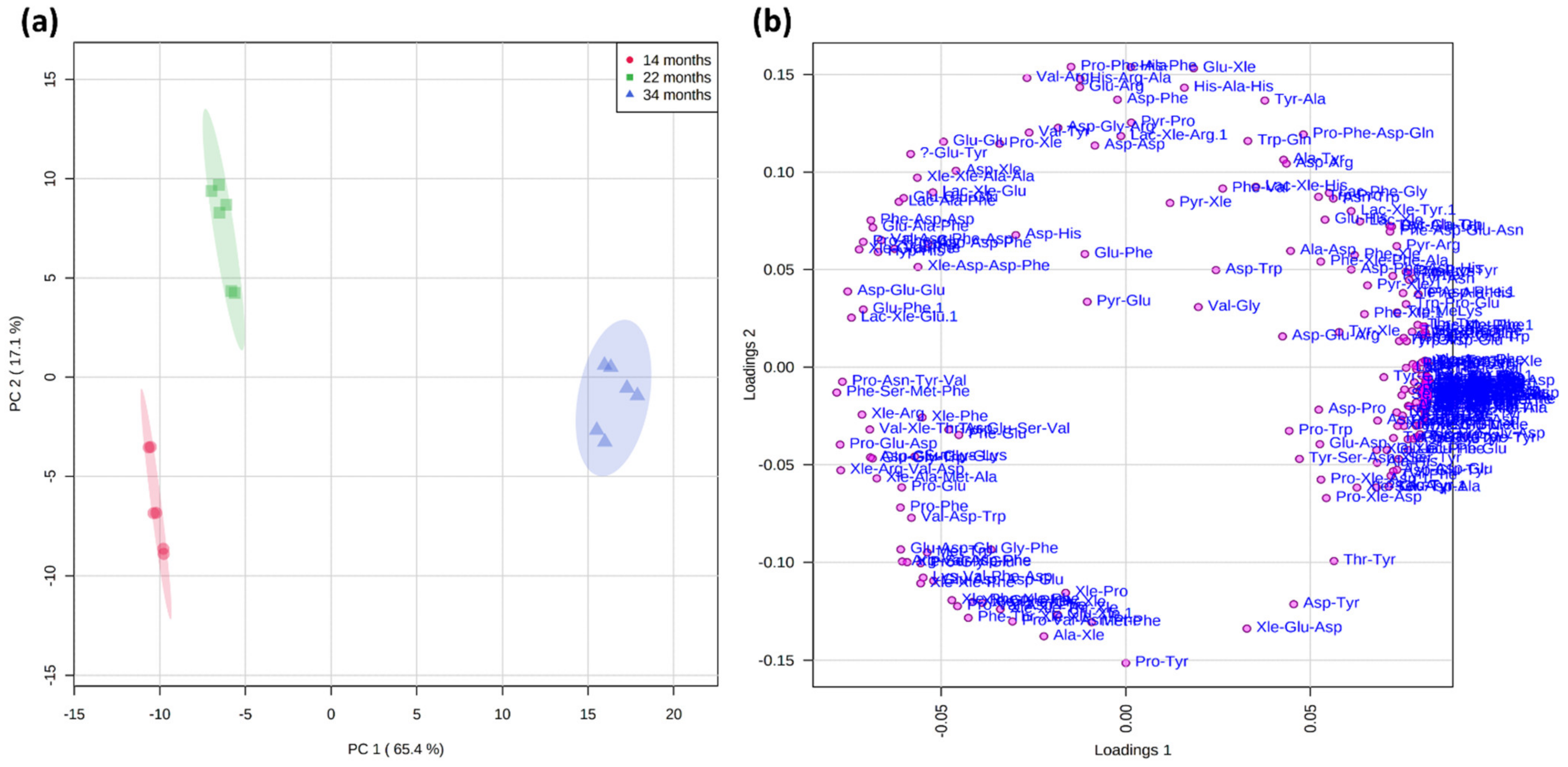
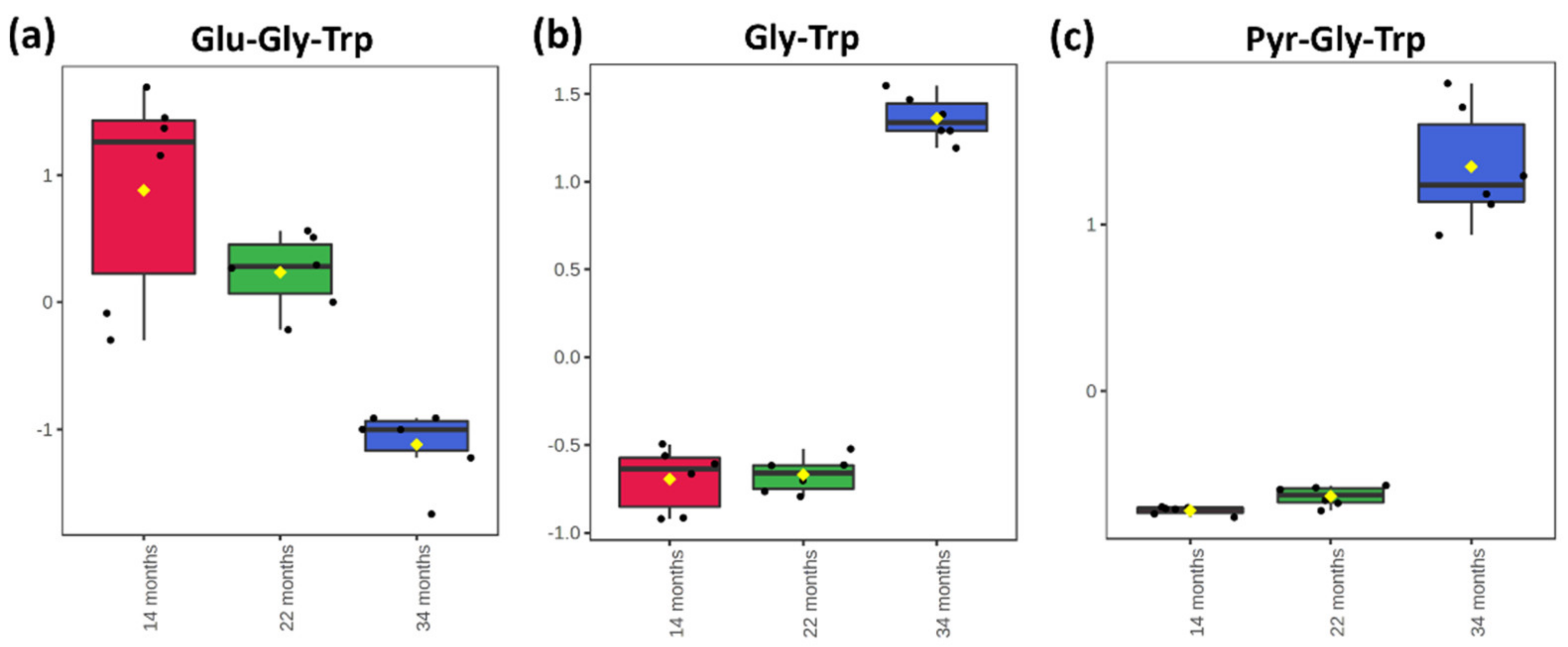
2.3. Principal Component Analysis of the Short Peptidome Profile of Dry-Cured Ham
3. Materials and Methods
3.1. Chemicals and Sample Collection
3.2. Short Peptide Extraction
3.3. Short Peptide Purification on GCB
3.4. Liquid Chromatography-Mass Spectrometry and Short Peptide Identification
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.-C.; Mora, L. Bioactive peptides generated in the processing of dry-cured ham. Food Chem. 2020, 321, 126689. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Flores, M. The Role of Muscle Proteases and Lipases in Flavor Development During the Processing of Dry-Cured Ham. Crit. Rev. Food Sci. Nutr. 1998, 38, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Rico, E.; Flores, J. Cathepsin B, D, H and L activities in the processing of dry-cured ham. J. Sci. Food Agric. 1993, 62, 157–161. [Google Scholar] [CrossRef]
- Jurado, Á.; García, C.; Timón, M.L.; Carrapiso, A.I. Effect of ripening time and rearing system on amino acid-related flavour compounds of Iberian ham. Meat Sci. 2007, 75, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Sentandreu, M.A.; Stoeva, S.; Aristoy, M.C.; Laib, K.; Voelter, W.; Toldra, E. Identification of Small Peptides Generated in Spanish Dry-cured Ham. J. Food Sci. 2003, 68, 64–69. [Google Scholar] [CrossRef]
- Sforza, S.; Pigazzani, A.; Motti, M.; Porta, C.; Virgili, R.; Galaverna, G.; Dossena, A.; Marchelli, R. Oligopeptides and free amino acids in Parma hams of known cathepsin B activity. Food Chem. 2001, 75, 267–273. [Google Scholar] [CrossRef]
- Stadnik, J.; Dolatowski, Z.J. Free Amino Acids and Biogenic Amines Content during Ageing of Dry-cured Pork Loins Inoculated with Lactobacillus casei ŁOCK 0900 Probiotic Strain. Food Sci. Technol. Res. 2015, 21, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Kęska, P.; Stadnik, J. Taste-active peptides and amino acids of pork meat as components of dry-cured meat products: An in-silico study. J. Sens. Stud. 2017, 32, e12301. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M.; Sieniawski, K.; Starowicz, P. BIOPEP database of sensory peptides and amino acids. Food Res. Int. 2016, 85, 155–161. [Google Scholar] [CrossRef]
- Van den Oord, A.H.A.; van Wassenaar, P.D. Umami peptides: Assessment of their alleged taste properties. Z. Leb. Forsch. A 1997, 205, 125–130. [Google Scholar] [CrossRef]
- Webb, K.E.; Matthews, J.C.; DiRienzo, D.B. Peptide absorption: A review of current concepts and future perspectives. J. Anim. Sci. 1992, 70, 3248–3257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, M.; Aristoy, M.-C.; Toldrá, F. Dipeptidyl peptidase IV inhibitory peptides generated in Spanish dry-cured ham. Meat Sci. 2014, 96, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, M.; Mora, L.; Hayes, M.; Reig, M.; Toldrá, F. Peptides with Potential Cardioprotective Effects Derived from Dry-Cured Ham Byproducts. J. Agric. Food Chem. 2019, 67, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian dry-cured ham as a potential source of α-glucosidase-inhibitory peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Paolella, S.; Prandi, B.; Falavigna, C.; Buhler, S.; Dossena, A.; Sforza, S.; Galaverna, G. Occurrence of non-proteolytic amino acyl derivatives in dry-cured ham. Food Res. Int. 2018, 114, 38–46. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, H.; Niu, H.; Wu, R. Peptidomic analyses: The progress in enrichment and identification of endogenous peptides. TrAC Trends Anal. Chem. 2020, 125, 115835. [Google Scholar] [CrossRef]
- Heres, A.; Yokoyama, I.; Gallego, M.; Toldrá, F.; Arihara, K.; Mora, L. Antihypertensive potential of sweet Ala-Ala dipeptide and its quantitation in dry-cured ham at different processing conditions. J. Funct. Foods 2021, 87, 104818. [Google Scholar] [CrossRef]
- Gallego, M.; Toldrá, F.; Mora, L. Quantification and in silico analysis of taste dipeptides generated during dry-cured ham processing. Food Chem. 2022, 370, 130977. [Google Scholar] [CrossRef]
- Cerrato, A.; Aita, S.E.; Capriotti, A.L.; Cavaliere, C.; Montone, C.M.; Laganà, A.; Piovesana, S. A new opening for the tricky untargeted investigation of natural and modified short peptides. Talanta 2020, 219, 121262. [Google Scholar] [CrossRef]
- Fricker, L.D. Limitations of Mass Spectrometry-Based Peptidomic Approaches. J. Am. Soc. Mass Spectrom. 2015, 26, 1981–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montone, C.M.; Capriotti, A.L.; Cerrato, A.; Antonelli, M.; La Barbera, G.; Piovesana, S.; Laganà, A.; Cavaliere, C. Identification of bioactive short peptides in cow milk by high-performance liquid chromatography on C18 and porous graphitic carbon coupled to high-resolution mass spectrometry. Anal. Bioanal. Chem. 2019, 411, 3395–3404. [Google Scholar] [CrossRef] [PubMed]
- Piovesana, S.; Capriotti, A.L.; Cerrato, A.; Crescenzi, C.; La Barbera, G.; Laganà, A.; Montone, C.M.; Cavaliere, C. Graphitized Carbon Black Enrichment and UHPLC-MS/MS Allow to Meet the Challenge of Small Chain Peptidomics in Urine. Anal. Chem. 2019, 91, 11474–11481. [Google Scholar] [CrossRef]
- Xiao, Y.; Vecchi, M.M.; Wen, D. Distinguishing between Leucine and Isoleucine by Integrated LC–MS Analysis Using an Orbitrap Fusion Mass Spectrometer. Anal. Chem. 2016, 88, 10757–10766. [Google Scholar] [CrossRef]
- Virgili, R.; Parolari, G.; Bordini, C.S.; Schivazappa, C.; Cornet, M.; Monin, G. Free Amino Acids and Dipeptides in Dry-Cured Ham. J. Muscle Foods 1999, 10, 119–130. [Google Scholar] [CrossRef]
- Toldra, F. Proteolysis and lipolysis in flavour development of dry-cured meat products. Meat Sci. 1998, 49, S101–S110. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Toldrá, F. The relevance of dipeptides and tripeptides in the bioactivity and taste of dry-cured ham. Food Prod. Process. Nutr. 2019, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the Improved Discovery and Design of Functional Peptides: Common Features of Diverse Classes Permit Generalized Prediction of Bioactivity. PLoS ONE 2012, 7, e45012. [Google Scholar] [CrossRef] [Green Version]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The Future of Peptide-based Drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef]
- Sgarbi, E.; Lazzi, C.; Iacopino, L.; Bottesini, C.; Lambertini, F.; Sforza, S.; Gatti, M. Microbial origin of non proteolytic aminoacyl derivatives in long ripened cheeses. Food Microbiol. 2013, 35, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Sforza, S.; Cavatorta, V.; Galaverna, G.; Dossena, A.; Marchelli, R. Accumulation of non-proteolytic aminoacyl derivatives in Parmigiano-Reggiano cheese during ripening. Int. Dairy J. 2009, 19, 582–587. [Google Scholar] [CrossRef]
- Schilling, S.; Stenzel, I.; von Bohlen, A.; Wermann, M.; Schulz, K.; Demuth, H.-U.; Wasternack, C. Isolation and characterization of the glutaminyl cyclases from Solanum tuberosum and Arabidopsis thaliana: Implications for physiological functions. Biol. Chem. 2007, 388, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Toelstede, S.; Hofmann, T. Kokumi-Active Glutamyl Peptides in Cheeses and Their Biogeneration by Penicillium roquefortii. J. Agric. Food Chem. 2009, 57, 3738–3748. [Google Scholar] [CrossRef]
- Szókán, G.; Kelemen, G.; Török, A. High-performance liquid chromatography of isopeptides. J. Chromatogr. A 1986, 366, 283–292. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Metabolomic Data Processing, Analysis, and Interpretation Using MetaboAnalyst. In Current Protocols in Bioinformatics; Wiley: New York, NY, USA, 2011; Volume 34. [Google Scholar] [CrossRef]
- Bro, R.; Smilde, A.K. Principal component analysis. Anal. Methods 2014, 6, 2812–2831. [Google Scholar] [CrossRef] [Green Version]
- Piasentier, E.; Pizzutti, N.; Lippe, G. The Influence of the Type of Dry-Cured Italian PDO Ham on Cathepsin B Activity Trend during Processing. Foods 2021, 10, 3123. [Google Scholar] [CrossRef]
- Gazme, B.; Boachie, R.T.; Tsopmo, A.; Udenigwe, C.C. Occurrence, properties and biological significance of pyroglutamyl peptides derived from different food sources. Food Sci. Hum. Wellness 2019, 8, 268–274. [Google Scholar] [CrossRef]
- Strohalm, M.; Kavan, D.; Novák, P.; Volný, M.; Havlíček, V. mMass 3: A Cross-Platform Software Environment for Precise Analysis of Mass Spectrometric Data. Anal. Chem. 2010, 82, 4648–4651. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerrato, A.; Aita, S.E.; Capriotti, A.L.; Cavaliere, C.; Montone, A.M.I.; Montone, C.M.; Laganà, A. Investigating the Short Peptidome Profile of Italian Dry-Cured Ham at Different Processing Times by High-Resolution Mass Spectrometry and Chemometrics. Int. J. Mol. Sci. 2022, 23, 3193. https://doi.org/10.3390/ijms23063193
Cerrato A, Aita SE, Capriotti AL, Cavaliere C, Montone AMI, Montone CM, Laganà A. Investigating the Short Peptidome Profile of Italian Dry-Cured Ham at Different Processing Times by High-Resolution Mass Spectrometry and Chemometrics. International Journal of Molecular Sciences. 2022; 23(6):3193. https://doi.org/10.3390/ijms23063193
Chicago/Turabian StyleCerrato, Andrea, Sara Elsa Aita, Anna Laura Capriotti, Chiara Cavaliere, Angela Michela Immacolata Montone, Carmela Maria Montone, and Aldo Laganà. 2022. "Investigating the Short Peptidome Profile of Italian Dry-Cured Ham at Different Processing Times by High-Resolution Mass Spectrometry and Chemometrics" International Journal of Molecular Sciences 23, no. 6: 3193. https://doi.org/10.3390/ijms23063193
APA StyleCerrato, A., Aita, S. E., Capriotti, A. L., Cavaliere, C., Montone, A. M. I., Montone, C. M., & Laganà, A. (2022). Investigating the Short Peptidome Profile of Italian Dry-Cured Ham at Different Processing Times by High-Resolution Mass Spectrometry and Chemometrics. International Journal of Molecular Sciences, 23(6), 3193. https://doi.org/10.3390/ijms23063193