
Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Characteristic of CDPKs Family in Sweet Potato and Its Two Diploid Relatives
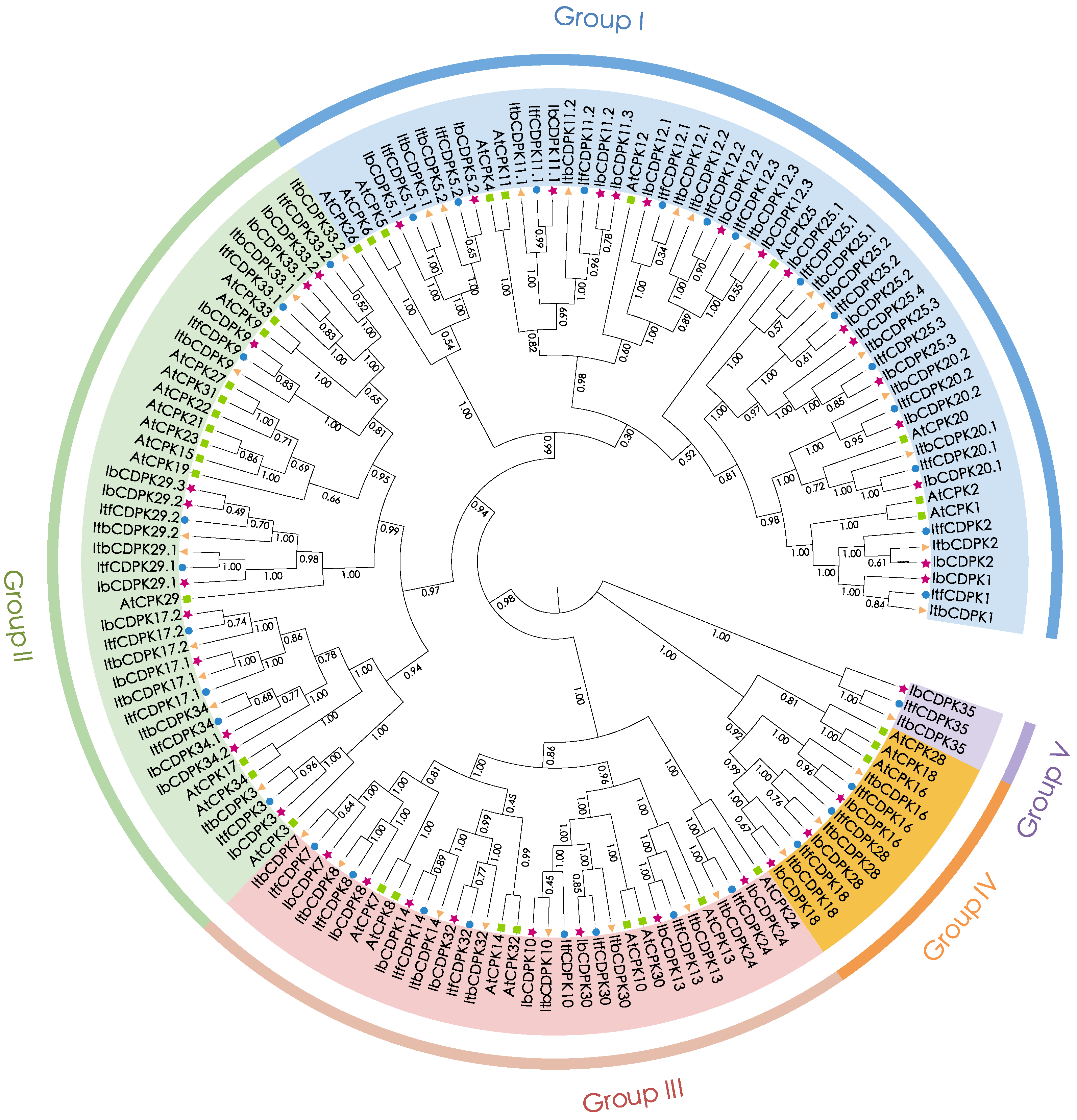
2.2. Phylogenetic Relationship of CDPKs in Sweet Potato and Its Two Diploid Relatives
2.3. Conserved Motif and Exon-Intron Structure Analysis of CDPKs in Sweet Potato and Its Two Diploid Relatives
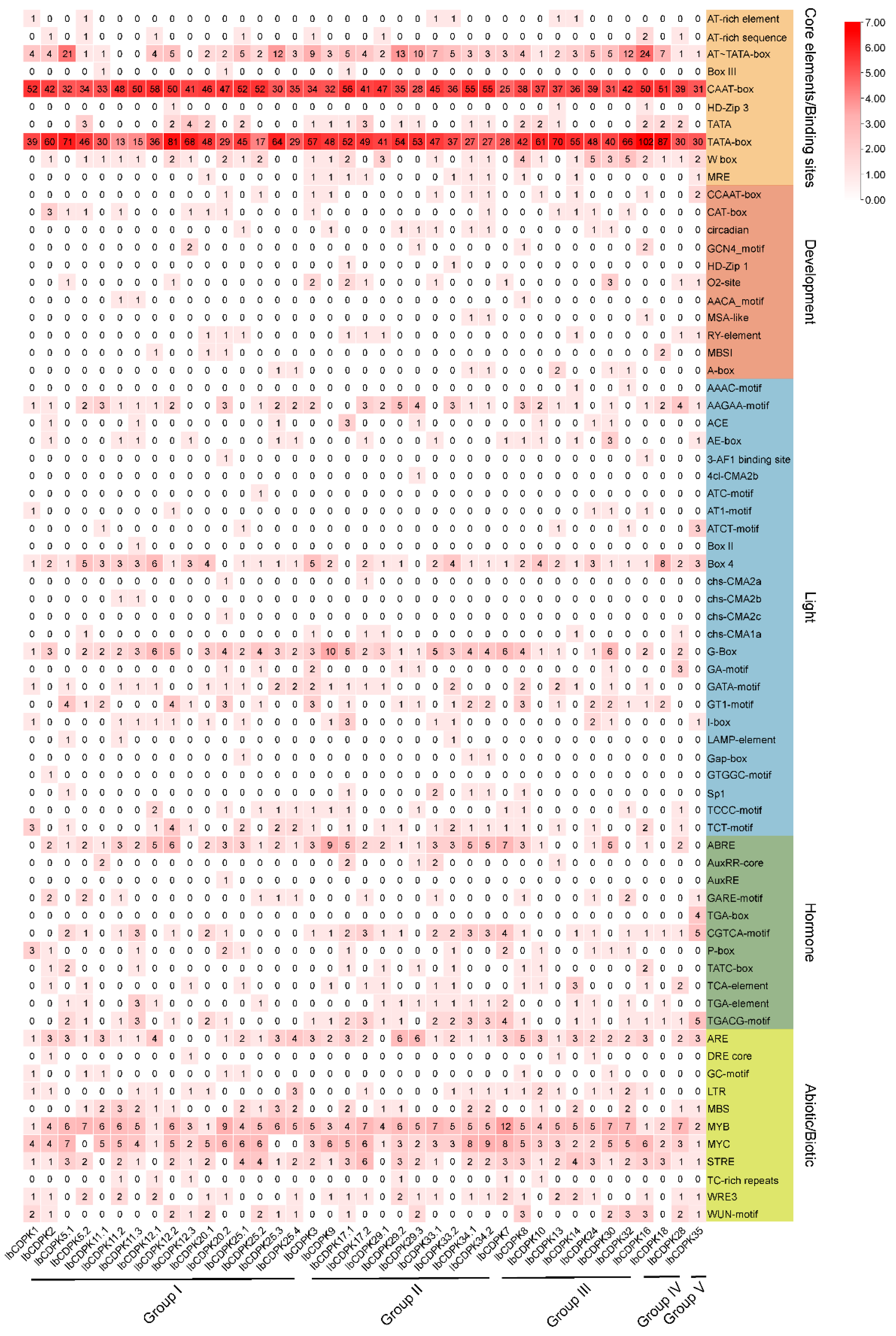
2.4. Cis-Element Analysis in the Promoter of IbCDPKs in Sweet Potato
2.5. Protein Interaction Network of IbCDPKs
2.6. Expression Analysis of CDPKs in Sweet Potato and Its Two Diploid Relatives
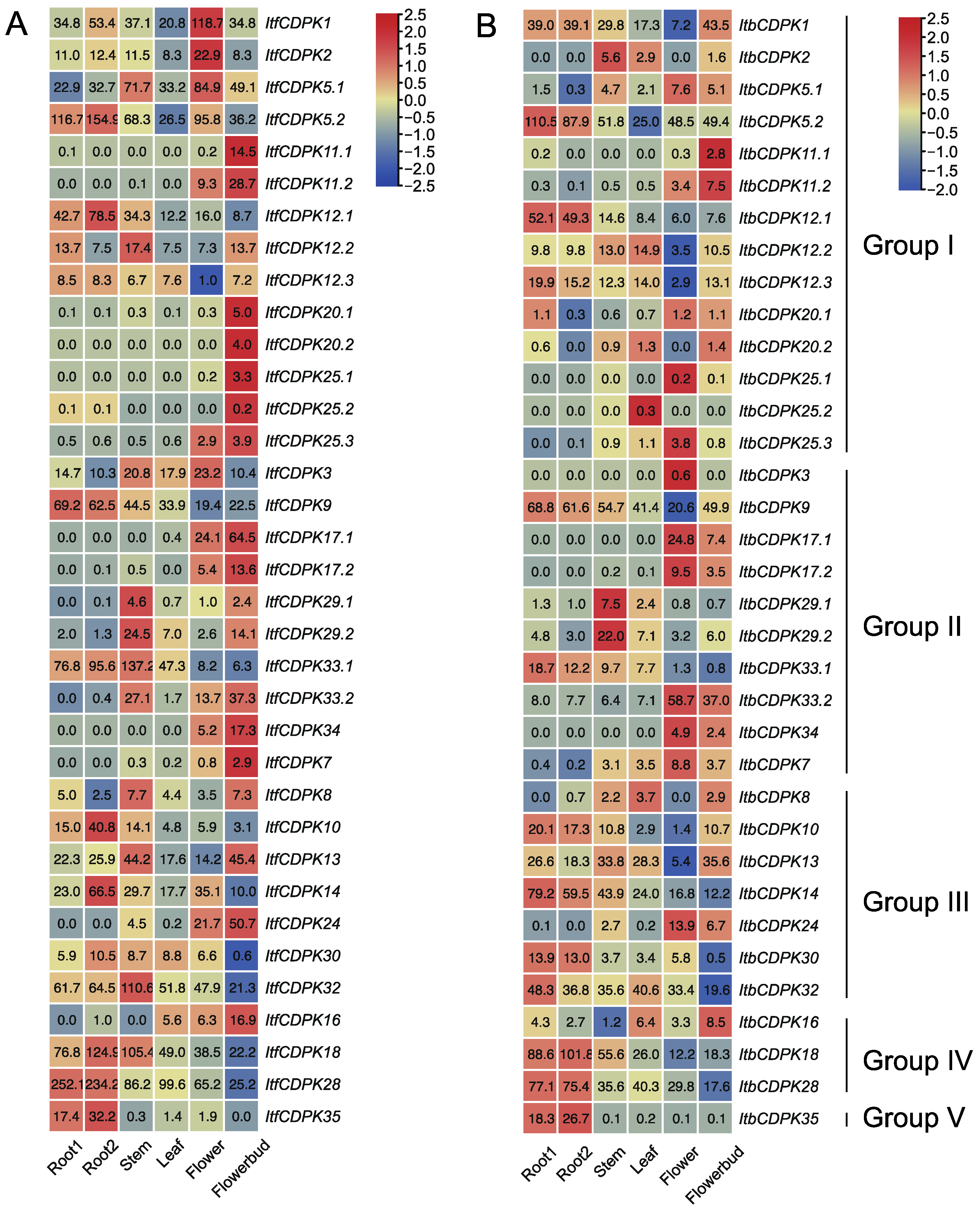
2.6.1. Expression Analysis in Various Tissues
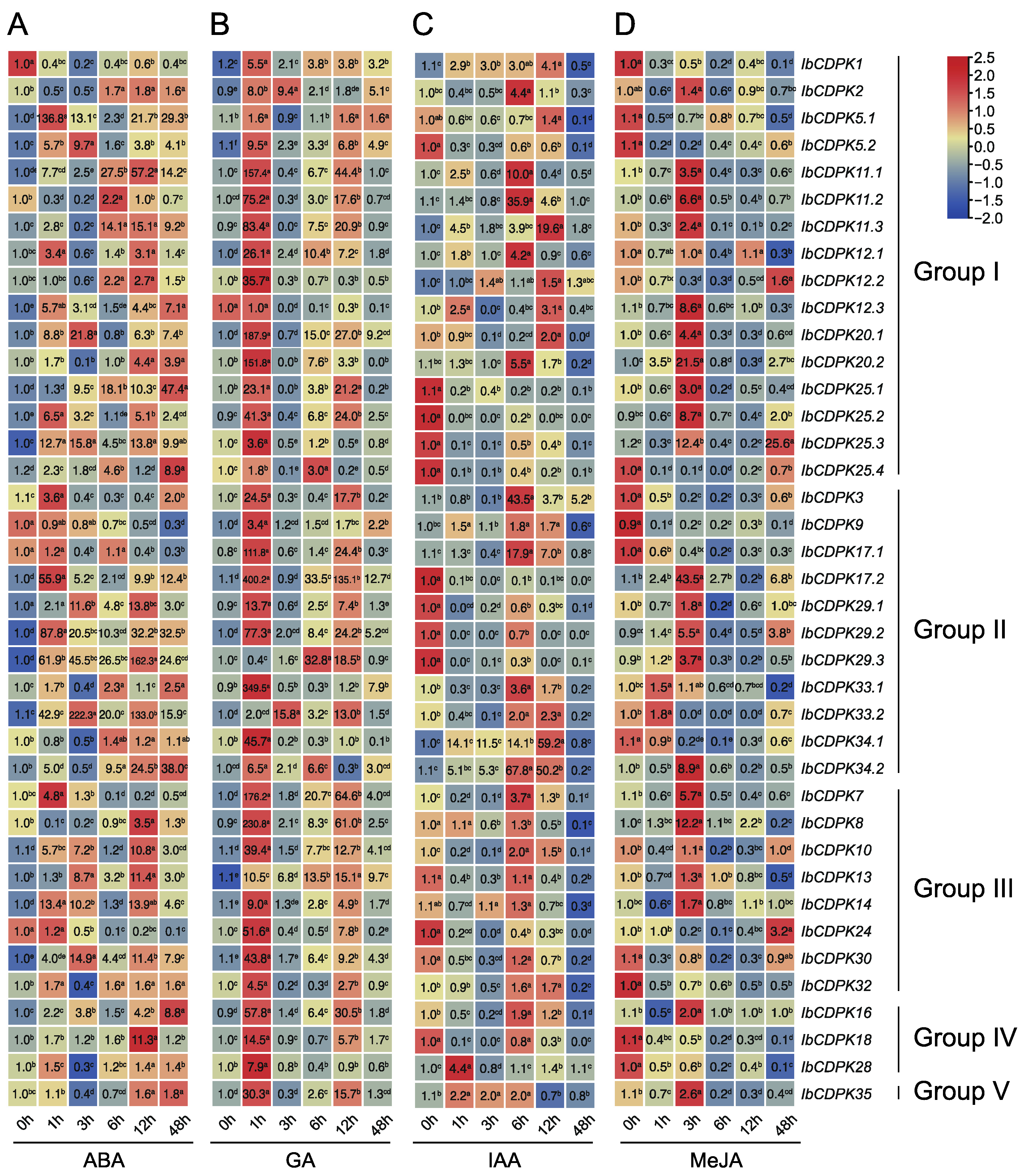
2.6.2. Expression Analysis of Hormone Response
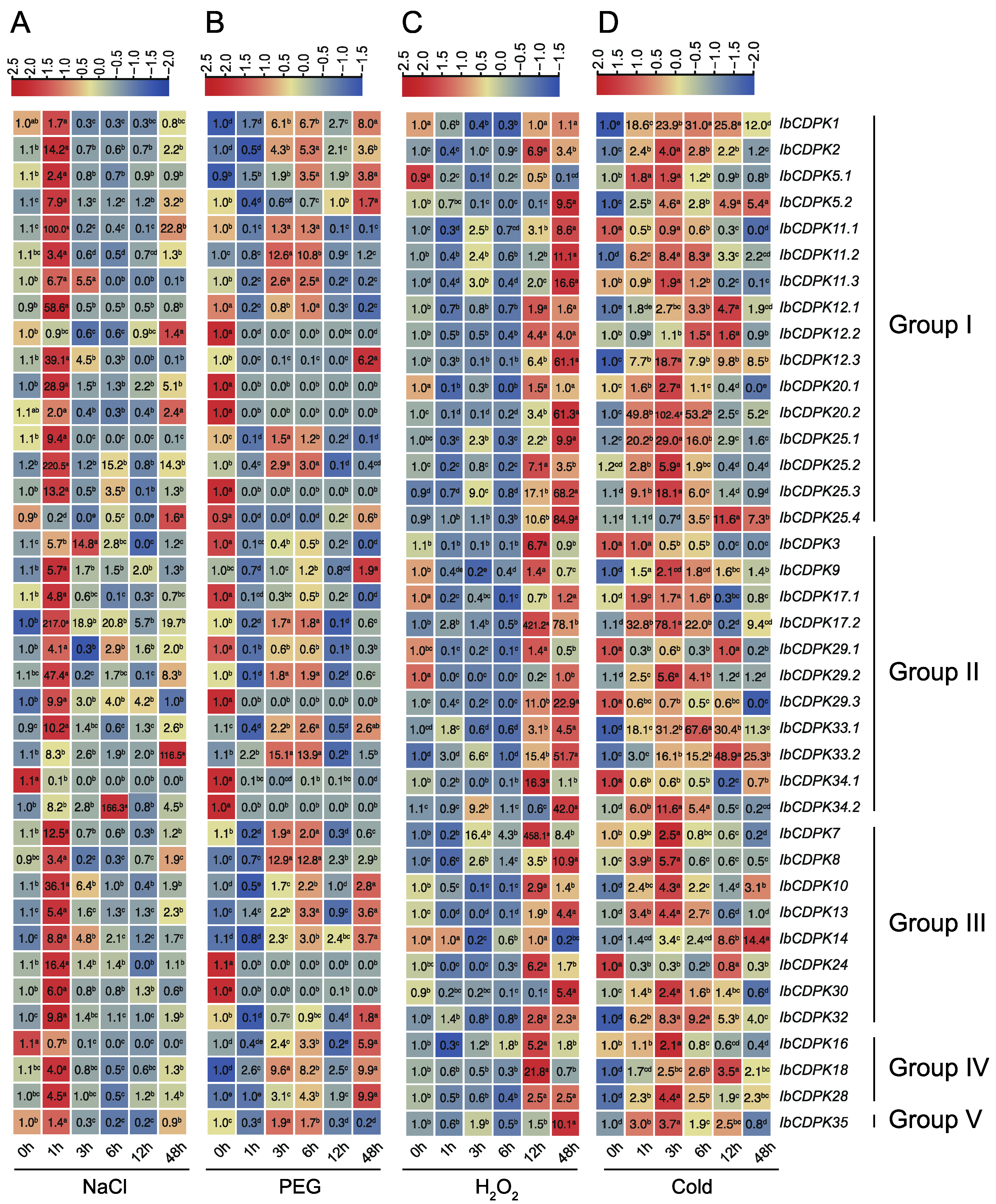
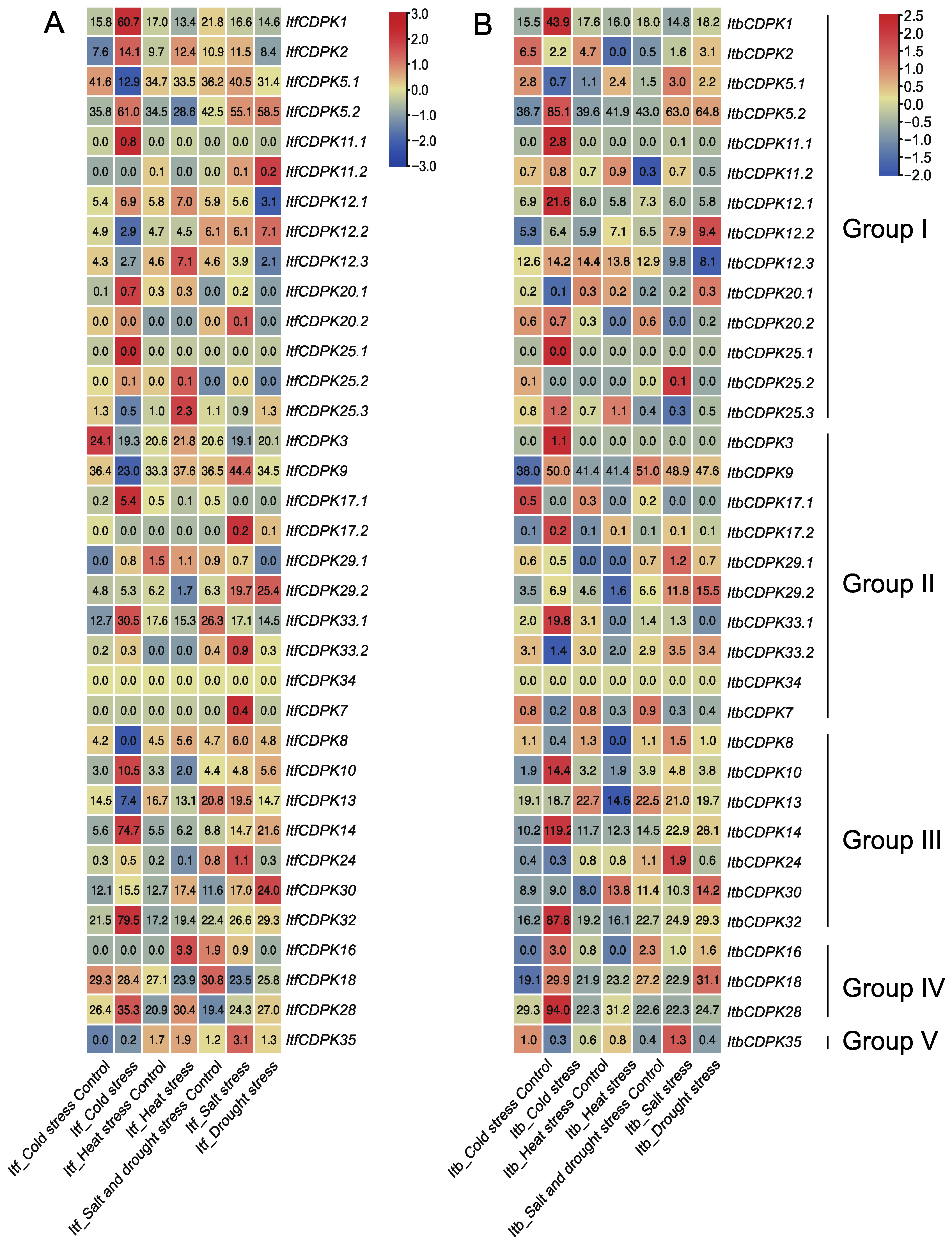
2.6.3. Expression Analysis under Abiotic Stresses
3. Discussion
3.1. Evolution of the CDPK Gene Family in Sweet Potato and Its Two Diploid Relatives
3.2. Different Functions of CDPKs on Growth and Development between Sweet Potato and Its Two Diploid Relatives
3.3. Different Functions of CDPKs on Hormone Crosstalk between Sweet Potato and Its Two Diploid Relatives
3.4. Different Functions of CDPKs on Multiple Abiotic Stress Response between Sweet Potato and Its Two Diploid Relatives
4. Materials and Methods
4.1. Identification of CDPKs
4.2. Chromosomal Distribution of CDPKs
4.3. Protein Properties Prediction of CDPKs
4.4. Phylogenetic Analysis of CDPKs
4.5. Domain Identification and Conserved Motifs Analysis of CDPKs
4.6. Exon–Intron Structures and Promoter Analysis of CDPKs
4.7. Protein Interaction Network of CDPKs
4.8. qRT-PCR Analysis of CDPKs
4.9. Transcriptome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudsocq, M.; Willmann, M.R.; McCormack, M.; Lee, H.; Shan, L.B.; He, P.; Bush, J.; Cheng, S.H.; Sheen, J. Differential innate immune signalling via Ca2+ sensor protein kinases. Nature 2010, 464, 418–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.H.; Willmann, M.R.; Chen, H.C.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.B.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Luan, S. The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.W.; Snedden, W.A. Calmodulin-Related Proteins Step Out from the Shadow of Their Namesake. Plant Physiol. 2013, 163, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.P.; Munyampundu, J.P.; Xu, Y.P.; Cai, X.Z. Phylogeny of Plant Calcium and Calmodulin-Dependent Protein Kinases (CCaMKs) and Functional Analyses of Tomato CCaMK in Disease Resistance. Front. Plant Sci. 2015, 6, 1075. [Google Scholar] [CrossRef] [Green Version]
- Poovaiah, B.W.; Du, L.; Wang, H.; Yang, T. Recent advances in calcium/calmodulin-mediated signaling with an emphasis on plant-microbe interactions. Plant Physiol. 2013, 163, 531–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Agarwal, P.; Arora, R.; Kapoor, S.; Tyagi, A.K. Expression analysis of calcium-dependent protein kinase gene family during reproductive development and abiotic stress conditions in rice (Oryza sativa L. ssp. indica). Mol. Genet. Genom. 2007, 278, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Li, A.L.; Zhu, Y.F.; Tan, X.M.; Wang, X.; Wei, B.; Guo, H.Z.; Zhang, Z.L.; Chen, X.B.; Zhao, G.Y.; Kong, X.Y.; et al. Evolutionary and functional study of the CDPK gene family in wheat (Triticum aestivum L.). Plant Mol. Biol. 2008, 66, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Liu, J.; Yang, X.; Ma, R. Genome-wide identification of the maize calcium-dependent protein kinase gene family. Appl. Biochem. Biotechnol. 2013, 169, 2111–2125. [Google Scholar] [CrossRef] [PubMed]
- Valmonte, G.R.; Arthur, K.; Higgins, C.M.; MacDiarmid, R.M. Calcium-dependent protein kinases in plants: Evolution, expression and function. Plant Cell Physiol. 2014, 55, 551–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Han, Y.T.; Zhao, F.L.; Hu, Y.; Gao, Y.R.; Ma, Y.F.; Zheng, Y.; Wang, Y.J.; Wen, Y.Q. Genome-wide Identification and Expression Analysis of the CDPK Gene Family in Grape, Vitis spp. BMC Plant Biol. 2015, 15, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.E.; Breton, G.; Harmon, A. Decoding Ca2+ signals through plant protein kinases. Annu. Rev. Plant Biol. 2004, 55, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.L.; Busconi, L. Membrane localization of a rice calcium-dependent protein kinase (CDPK) is mediated by myristoylation and palmitoylation. Plant J. 2000, 24, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Hrabak, E.M.; Chan, C.W.M.; Gribskov, M.; Harper, J.F.; Choi, J.H.; Halford, N.; Kudla, J.; Luan, S.; Nimmo, H.G.; Sussman, M.R.; et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003, 132, 666–680. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J. Ca2+-Dependent Protein Kinases and Stress Signal Transduction in Plants. Science 1997, 274, 1900–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.F.; Huang, J.F.; Lloyd, S.J. Genetic Identification of an Autoinhibitor in CDPK, a Protein-Kinase with a Calmodulin-Like Domain. Biochemistry 1994, 33, 7267–7277. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Wang, L.; Xie, W.; Wan, B.; Li, X.; Lin, Y. Expression profile of calcium-dependent protein kinase (CDPKs) genes during the whole lifespan and under phytohormone treatment conditions in rice (Oryza sativa L. ssp. indica). Plant Mol. Biol. 2009, 70, 311–325. [Google Scholar] [CrossRef]
- Mori, I.C.; Murata, Y.; Yang, Y.Z.; Munemasa, S.; Wang, Y.F.; Andreoli, S.; Tiriac, H.; Alonso, J.M.; Harper, J.F.; Ecker, J.R.; et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- and Ca2+-permeable channels and stomatal closure. PLoS Biol. 2006, 4, 1749–1762. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.Y.; Yu, X.C.; Wang, X.J.; Zhao, R.; Li, Y.; Fan, R.C.; Shang, Y.; Du, S.Y.; Wang, X.F.; Wu, F.Q.; et al. Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis. Plant Cell 2007, 19, 3019–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.Y.; Wu, W.H. AtCPK23 functions in Arabidopsis responses to drought and salt stresses. Plant Mol. Biol. 2007, 65, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, P.; Ehlert, B.; Romeis, T. ZmCPK1, a calcium-independent kinase member of the Zea mays CDPK gene family, functions as a negative regulator in cold stress signalling. Plant Cell Environ. 2015, 38, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cheng, Y.; Yu, X.; Liu, J.; Yang, L.; Gao, Y.; Ke, G.; Zhou, M.; Mu, B.; Xiao, S.; et al. Overexpression of two CDPKs from wild Chinese grapevine enhances powdery mildew resistance in Vitis vinifera and Arabidopsis. New Phytol. 2021, 230, 2029–2046. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Zhu, L.; Zhang, X.; Guan, Q.; Xiao, S.; Min, L.; Zhang, X. GhCPK33 Negatively Regulates Defense against Verticillium dahliae by Phosphorylating GhOPR3. Plant Physiol. 2018, 178, 876–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munemasa, S.; Hossain, M.A.; Nakamura, Y.; Mori, I.C.; Murata, Y. The Arabidopsis Calcium-Dependent Protein Kinase, CPK6, Functions as a Positive Regulator of Methyl Jasmonate Signaling in Guard Cells. Plant Physiol. 2011, 155, 553–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivashuta, S.; Liu, J.Y.; Liu, J.Q.; Lohar, D.P.; Haridas, S.; Bucciarelli, B.; VandenBosch, K.A.; Vance, C.P.; Harrison, M.J.; Gantt, J.S. RNA interference identifies a calcium-dependent protein kinase involved in Medicago truncatula root development. Plant Cell 2005, 17, 2911–2921. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.H.; Zhang, H.F.; Wei, X.Y.; Yang, L.; Yang, B.; Zhang, L.; Li, J.; Jiang, Y.Q. Functional characterization of calcium-dependent protein kinase (CPK) 2 gene from oilseed rape (Brassica napus L.) in regulating reactive oxygen species signaling and cell death control. Gene 2018, 651, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.; Romanowsky, S.M.; Barron, Y.D.; Garg, S.; Azuse, C.L.; Curran, A.; Davis, R.M.; Hatton, J.; Harmon, A.C.; Harper, J.F. Calcium-dependent protein kinases regulate polarized tip growth in pollen tubes. Plant J. 2009, 59, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, K.; Pawelek, A.; Kopcewicz, J.; Szmidt-Jaworska, A. The calcium-dependent protein kinase (PnCDPK1) is involved in Pharbitis nil flowering. J. Plant Physiol. 2012, 169, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Matschi, S.; Werner, S.; Schulze, W.X.; Legen, J.; Hilger, H.H.; Romeis, T. Function of calcium-dependent protein kinase CPK28 of Arabidopsis thaliana in plant stem elongation and vascular development. Plant J. 2013, 73, 883–896. [Google Scholar] [CrossRef]
- Saijo, Y.; Kinoshita, N.; Ishiyama, K.; Hata, S.; Kyozuka, J.; Hayakawa, T.; Nakamura, T.; Shimamoto, K.; Yamaya, T.; Izui, K. A Ca2+-dependent protein kinase that endows rice plants with cold- and salt-stress tolerance functions in vascular bundles. Plant Cell Physiol. 2001, 42, 1228–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.I.; Park, H.J.; Park, J.H.; Kim, S.; Im, M.Y.; Seo, H.H.; Kim, Y.W.; Hwang, I.; Kim, S.Y. Arabidopsis calcium-dependent protein kinase AtCPK32 interacts with ABF4, a transcriptional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol. 2005, 139, 1750–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.X.; Shen, S.H.; Yang, S.H.; Komatsu, S. OsCDPK13, a calcium-dependent protein kinase gene from rice, is induced in response to cold and gibberellin. Plant Physiol. Bioch. 2003, 41, 369–374. [Google Scholar] [CrossRef]
- Liu, Q.C. Sweet potato Omics and Biotechnology in China. Plant Omics 2011, 4, 295–301. [Google Scholar] [CrossRef]
- Bach, T.J. Some New Aspects of Isoprenoid Biosynthesis in Plants-a Review. Lipids 1995, 30, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.L.; Zheng, J.L.; Sun, Z.; Fan, W.J.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef]
- Wu, S.; Lau, K.H.; Cao, Q.H.; Hamilton, J.P.; Sun, H.H.; Zhou, C.X.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.Y.; et al. Genome sequences of two diploid wild relatives of cultivated sweetpotato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 2580. [Google Scholar] [CrossRef] [Green Version]
- Chalker, D.L.; Sandmeyer, S.B. Sites of Rna Polymerase-Iii Transcription Initiation and Ty3 Integration at the U6 Gene Are Positioned by the Tata Box. Proc. Natl. Acad. Sci. USA 1993, 90, 4927–4931. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Zhao, X.Y.; Liu, Y.B.; Zhang, C.L.; O’Neill, S.D.; Zhang, X.S. Auxin-induced WUS expression is essential for embryonic stem cell renewal during somatic embryogenesis in Arabidopsis. Plant J. 2009, 59, 448–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Schumaker, K.S.; Guo, Y. Sumoylation of transcription factor MYB30 by the small ubiquitin-like modifier E3 ligase SIZ1 mediates abscisic acid response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2012, 109, 12822–12827. [Google Scholar] [CrossRef] [Green Version]
- Ju, L.; Jing, Y.X.; Shi, P.T.; Liu, J.; Chen, J.S.; Yan, J.J.; Chu, J.F.; Chen, K.M.; Sun, J.Q. JAZ proteins modulate seed germination through interaction with ABI5 in bread wheat and Arabidopsis. New Phytol. 2019, 223, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.A.; Wang, L.J.; Xu, Z.H.; Xia, Z.A. Foliar modifications induced by inhibition of polar transport of auxin. Cell Res. 1999, 9, 27–35. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, J.Q.; Zhai, Q.Z.; Zhou, W.K.; Qi, L.L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H.L.; Qi, J.; et al. The Basic Helix-Loop-Helix Transcription Factor MYC2 Directly Represses PLETHORA Expression during Jasmonate-Mediated Modulation of the Root Stem Cell Niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.Y.; Wabnik, K.; Niu, T.T.; Li, H.B.; Yu, Q.Q.; Pollmann, S.; Vanneste, S.; Govaerts, W.; Rolcik, J.; Geisler, M.; et al. WOX5-IAA17 Feedback Circuit-Mediated Cellular Auxin Response Is Crucial for the Patterning of Root Stem Cell Niches in Arabidopsis. Mol. Plant 2014, 7, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, C.B.; Xiao, L.H.; Huaa, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. USA 2018, 115, 6058–6063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.L.; Zhou, L.M.; Huang, M.K.; He, X.M.; Yang, Y.H.; Liu, X.; Li, Y.G.; Hou, X.L. Gibberellins play an essential role in late embryogenesis of Arabidopsis. Nat. Plants 2018, 4, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Y.; Chen, M.X.; Chen, Q.F.; Xiao, S.; Chye, M.L. Arabidopsis acyl-CoA-binding protein ACBP1 participates in the regulation of seed germination and seedling development. Plant J. 2013, 74, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Feng, C.Z.; Ye, Q.; Wu, W.H.; Chen, Y.F. Arabidopsis WRKY6 Transcription Factor Acts as a Positive Regulator of Abscisic Acid Signaling during Seed Germination and Early Seedling Development. PLoS Genet. 2016, 12, e1005833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.P.; Kuo, C.H.; Lu, H.H.; Lo, H.S.; Yeh, K.W. The Sweet Potato NAC-Domain Transcription Factor IbNAC1 Is Dynamically Coordinated by the Activator IbbHLH3 and the Repressor IbbHLH4 to Reprogram the Defense Mechanism against Wounding. PLoS Genet. 2016, 12, e1006397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Q.; Zhai, H.; Gao, S.P.; Yang, L.; Wang, Z.; Xu, Y.T.; Huo, J.X.; Ren, Z.T.; Zhao, N.; et al. IbBBX24 Promotes the Jasmonic Acid Pathway and Enhances Fusarium Wilt Resistance in Sweet Potato. Plant Cell 2020, 32, 1102–1123. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.M.; Dowd, P.E.; Gilroy, S.; McCubbin, A.G. Calcium-dependent protein kinase isoforms in Petunia have distinct functions in pollen tube growth, including regulating polarity. Plant Cell 2006, 18, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yu, C.; Xu, S.; Zhu, Y.; Huang, W. OsDi19-4 acts downstream of OsCDPK14 to positively regulate ABA response in rice. Plant Cell Environ. 2016, 39, 2740–2753. [Google Scholar] [CrossRef] [PubMed]
- Geiger, D.; Scherzer, S.; Mumm, P.; Marten, I.; Ache, P.; Matschi, S.; Liese, A.; Wellmann, C.; Al-Rasheid, K.A.S.; Grill, E.; et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proc. Natl. Acad. Sci. USA 2010, 107, 8023–8028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.S.; Zhang, D.; Wang, L.; Pan, J.W.; Liu, Y.; Kong, X.P.; Zhou, Y.; Li, D.Q. A maize calcium-dependent protein kinase gene, ZmCPK4, positively regulated abscisic acid signaling and enhanced drought stress tolerance in transgenic Arabidopsis. Plant Physiol. Bioch. 2013, 71, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, G.; Wang, C.; He, Z.Z.; Lan, X.X.; Zhang, S.Y.; Lan, H.Y. The bHLH transcription factor CgbHLH001 is a potential interaction partner of CDPK in halophyte Chenopodium glaucum. Sci. Rep. 2017, 7, 8441. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Jia, W.S.; Zhang, J.H. AtMKK1 mediates ABA-induced CAT1 expression and H2O2 production via AtMPK6-coupled signaling in Arabidopsis. Plant J. 2008, 54, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, W.Y.; Li, Z.H.; Deng, X.W.; Wu, W.H.; Xue, Y.B. F-Box Protein DOR Functions As a Novel Inhibitory Factor for Abscisic Acid-Induced Stomatal Closure under Drought Stress in Arabidopsis. Plant Physiol. 2008, 148, 2121–2133. [Google Scholar] [CrossRef] [Green Version]
- Schafer, B.W.; Heizmann, C.W. The S100 family of EF-hand calcium-binding proteins: Functions and pathology. Trends Biochem. Sci. 1996, 21, 134–140. [Google Scholar] [CrossRef]
- Frattini, M.; Morello, L.; Breviario, D. Rice calcium-dependent protein kinase isoforms OsCDPK2 and OsCDPK11 show different responses to light and different expression patterns during seed development. Plant Mol. Biol. 1999, 41, 753–764. [Google Scholar] [CrossRef]
- Yu, T.F.; Zhao, W.Y.; Fu, J.D.; Liu, Y.W.; Chen, M.; Zhou, Y.B.; Ma, Y.Z.; Xu, Z.S.; Xi, Y.J. Genome-Wide Analysis of CDPK Family in Foxtail Millet and Determination of SiCDPK24 Functions in Drought Stress. Front. Plant Sci. 2018, 9, 651. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Xie, Y.B.; Zheng, Y.Y.; Li, H.Y.; Luo, X.T.; He, Z.H.; Cao, S.; Shi, Y.; Zhao, Q.; Xue, Y.; Zuo, Z.X.; et al. GPS-Lipid: A robust tool for the prediction of multiple lipid modification sites. Sci. Rep. 2016, 6, 28249. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | CDS Length/bp | Protein Size/aa | MW/kDa | pI | No. of EF-Hands | N-Myristoylation | Palmitoylation |
|---|---|---|---|---|---|---|---|---|
| IbCDPK1 | Ib01g3472 | 1755 | 584 | 65.253 | 5.72 | 4 | No | No |
| IbCDPK2 | Ib07g25704 | 1962 | 653 | 72.939 | 6 | 4 | Yes | Yes |
| IbCDPK3 | Ib05g19617 | 2058 | 685 | 77.012 | 5.55 | 4 | Yes | Yes |
| IbCDPK5.1 | Ib10g41295 | 1701 | 566 | 62.970 | 5.2 | 4 | Yes | Yes |
| IbCDPK5.2 | Ib14g59849 | 1689 | 562 | 63.084 | 5.49 | 4 | Yes | Yes |
| IbCDPK7 | Ib12g50922 | 1677 | 558 | 62.852 | 8.69 | 4 | Yes | Yes |
| IbCDPK8 | Ib03g11409 | 1602 | 533 | 59.869 | 6.15 | 4 | Yes | Yes |
| IbCDPK9 | Ib09g34428 | 1563 | 520 | 58.469 | 5.98 | 4 | Yes | Yes |
| IbCDPK10 | Ib13g53626 | 1770 | 589 | 66.286 | 7.16 | 4 | Yes | Yes |
| IbCDPK11.1 | Ib01g3989 | 1416 | 471 | 52.521 | 6.05 | 4 | Yes | No |
| IbCDPK11.2 | Ib02g8624 | 1632 | 543 | 60.666 | 5.31 | 4 | No | Yes |
| IbCDPK11.3 | Ib02g6000 | 1509 | 502 | 55.721 | 5.67 | 4 | No | Yes |
| IbCDPK12.1 | Ib07g27783 | 1530 | 509 | 57.151 | 5.01 | 4 | No | No |
| IbCDPK12.2 | Ib05g20844 | 1518 | 505 | 56.550 | 4.97 | 4 | No | Yes |
| IbCDPK12.3 | Ib09g35646 | 1668 | 555 | 62.731 | 5.71 | 3 | No | No |
| IbCDPK13 | Ib14g58861 | 1461 | 486 | 54.436 | 5.63 | 4 | Yes | Yes |
| IbCDPK14 | Ib14g58509 | 1614 | 537 | 60.600 | 6.25 | 4 | Yes | Yes |
| IbCDPK16 | Ib07g29399 | 3513 | 1170 | 130.824 | 8.09 | 4 | Yes | Yes |
| IbCDPK17.1 | Ib02g4428 | 1716 | 571 | 63.410 | 5.53 | 4 | Yes | Yes |
| IbCDPK17.2 | Ib12g50948 | 1575 | 524 | 58.850 | 5.31 | 4 | Yes | Yes |
| IbCDPK18 | Ib01g414 | 1866 | 621 | 70.113 | 8.65 | 3 | Yes | Yes |
| IbCDPK20.1 | Ib14g58329 | 1635 | 544 | 59.975 | 5.53 | 4 | Yes | Yes |
| IbCDPK20.2 | Ib06g25035 | 1548 | 515 | 56.855 | 5.02 | 4 | Yes | Yes |
| IbCDPK24 | Ib09g36010 | 1353 | 450 | 51.744 | 6.05 | 3 | No | No |
| IbCDPK25.1 | Ib06g25043 | 1911 | 636 | 71.191 | 5.89 | 4 | No | No |
| IbCDPK25.2 | Ib11g45599 | 1866 | 621 | 70.036 | 5.13 | 4 | Yes | Yes |
| IbCDPK25.3 | Ib14g58318 | 1767 | 588 | 65.341 | 5.56 | 4 | Yes | Yes |
| IbCDPK25.4 | Ib14g58307 | 1056 | 351 | 38.732 | 4.75 | 3 | Yes | Yes |
| IbCDPK28 | Ib05g16972 | 1800 | 599 | 67.199 | 9.28 | 4 | Yes | Yes |
| IbCDPK29.1 | Ib10g38551 | 1497 | 498 | 56.102 | 5.74 | 4 | No | No |
| IbCDPK29.2 | Ib15g60594 | 1443 | 480 | 53.887 | 4.87 | 4 | Yes | Yes |
| IbCDPK29.3 | Ib15g60591 | 1518 | 505 | 56.806 | 5.14 | 3 | Yes | Yes |
| IbCDPK30 | Ib08g33870 | 1692 | 563 | 63.786 | 6.08 | 4 | Yes | Yes |
| IbCDPK32 | Ib06g24869 | 1560 | 519 | 58.264 | 7.62 | 3 | Yes | Yes |
| IbCDPK33.1 | Ib01g217 | 1509 | 502 | 71.191 | 6.54 | 4 | Yes | No |
| IbCDPK33.2 | Ib15g61179 | 1554 | 517 | 57.706 | 5.88 | 4 | Yes | Yes |
| IbCDPK34.1 | Ib03g11293 | 1683 | 560 | 62.193 | 5.84 | 4 | Yes | Yes |
| IbCDPK34.2 | Ib03g11291 | 1578 | 525 | 58.304 | 5.49 | 4 | Yes | Yes |
| IbCDPK35 | Ib06g24207 | 1488 | 495 | 54.747 | 5.23 | 3 | No | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhao, L.; Zhang, H.; Liu, Q.; Zhai, H.; Zhao, N.; Gao, S.; He, S. Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2022, 23, 3088. https://doi.org/10.3390/ijms23063088
Li X, Zhao L, Zhang H, Liu Q, Zhai H, Zhao N, Gao S, He S. Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences. 2022; 23(6):3088. https://doi.org/10.3390/ijms23063088
Chicago/Turabian StyleLi, Xu, Limeng Zhao, Huan Zhang, Qingchang Liu, Hong Zhai, Ning Zhao, Shaopei Gao, and Shaozhen He. 2022. "Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives" International Journal of Molecular Sciences 23, no. 6: 3088. https://doi.org/10.3390/ijms23063088
APA StyleLi, X., Zhao, L., Zhang, H., Liu, Q., Zhai, H., Zhao, N., Gao, S., & He, S. (2022). Genome-Wide Identification and Characterization of CDPK Family Reveal Their Involvements in Growth and Development and Abiotic Stress in Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences, 23(6), 3088. https://doi.org/10.3390/ijms23063088