
Genetic Variation in ZmPAT7 Contributes to Tassel Branch Number in Maize

 and
and
Abstract
:1. Introduction
2. Results
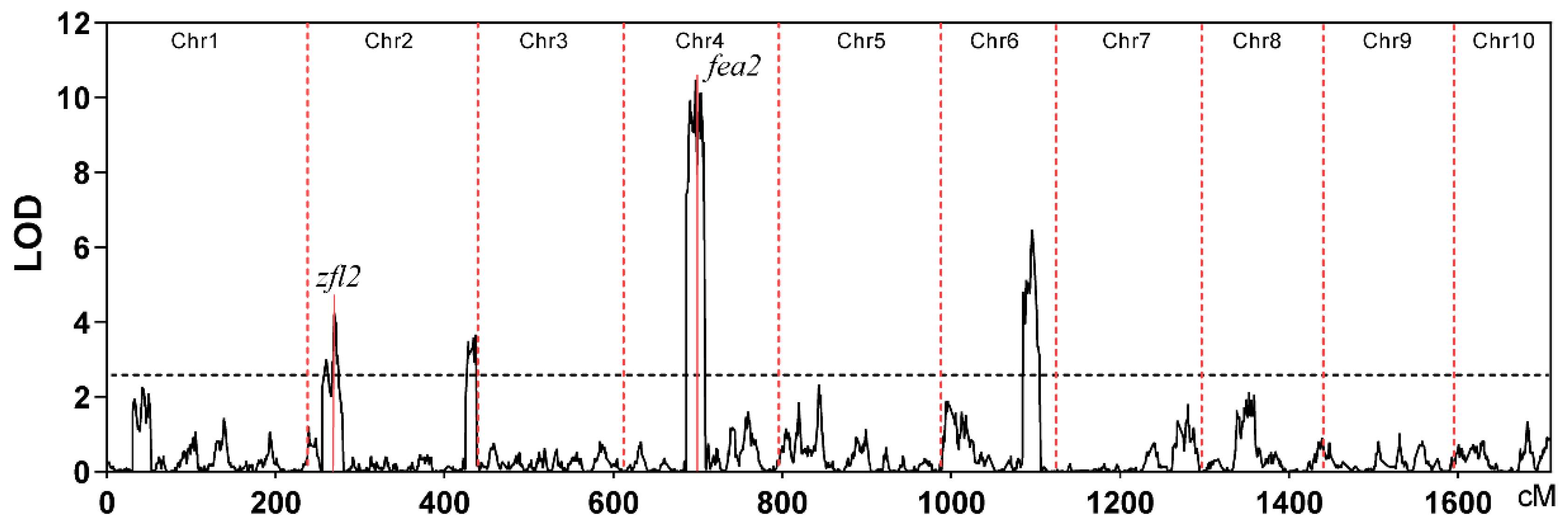
2.1. QTL Mapping and Identification of Candidate Genes for TBN
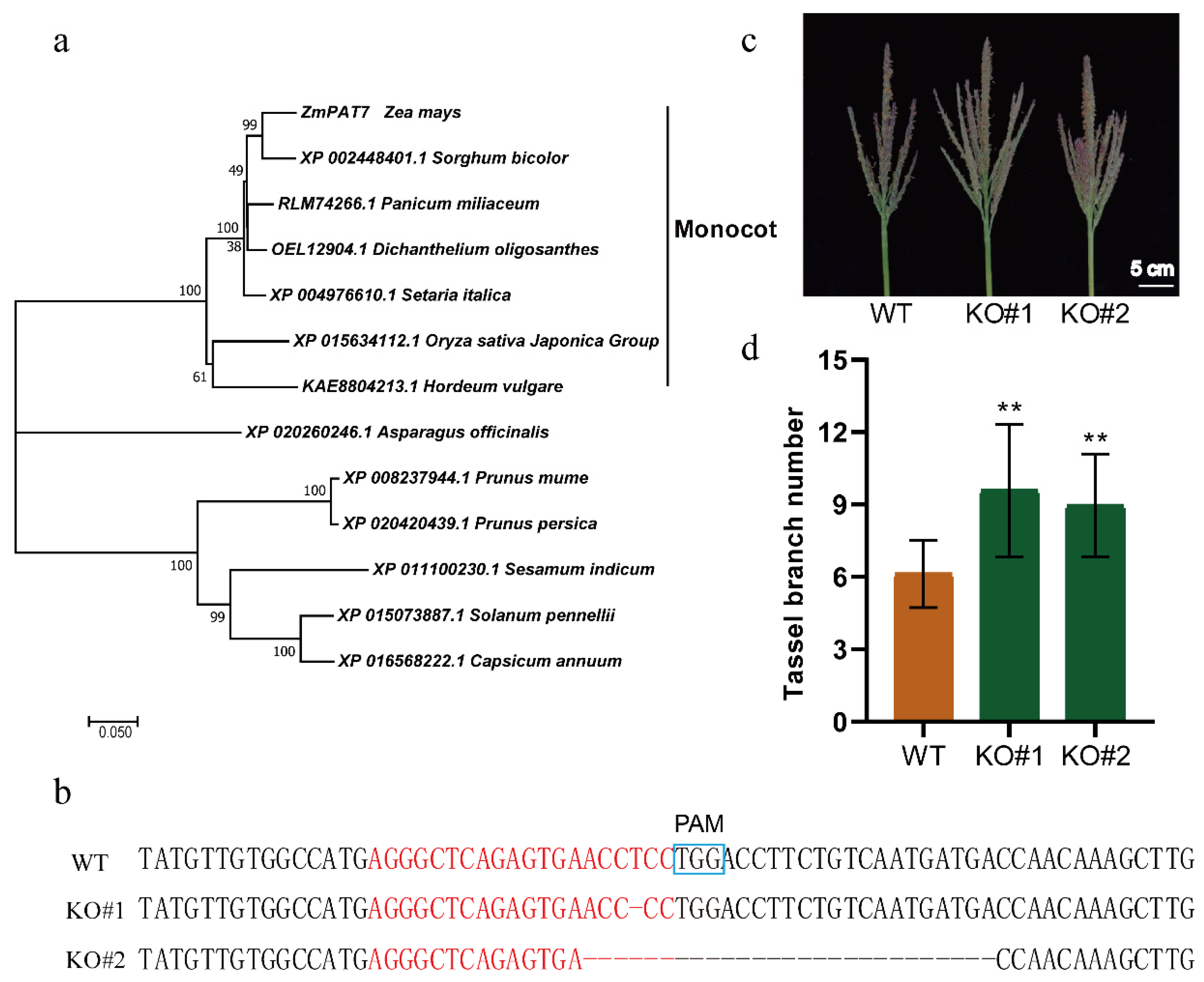
2.2. ZmPAT7 Negatively Controls Tassel Branch Number in Maize
2.3. Natural Alleles of ZmPAT7
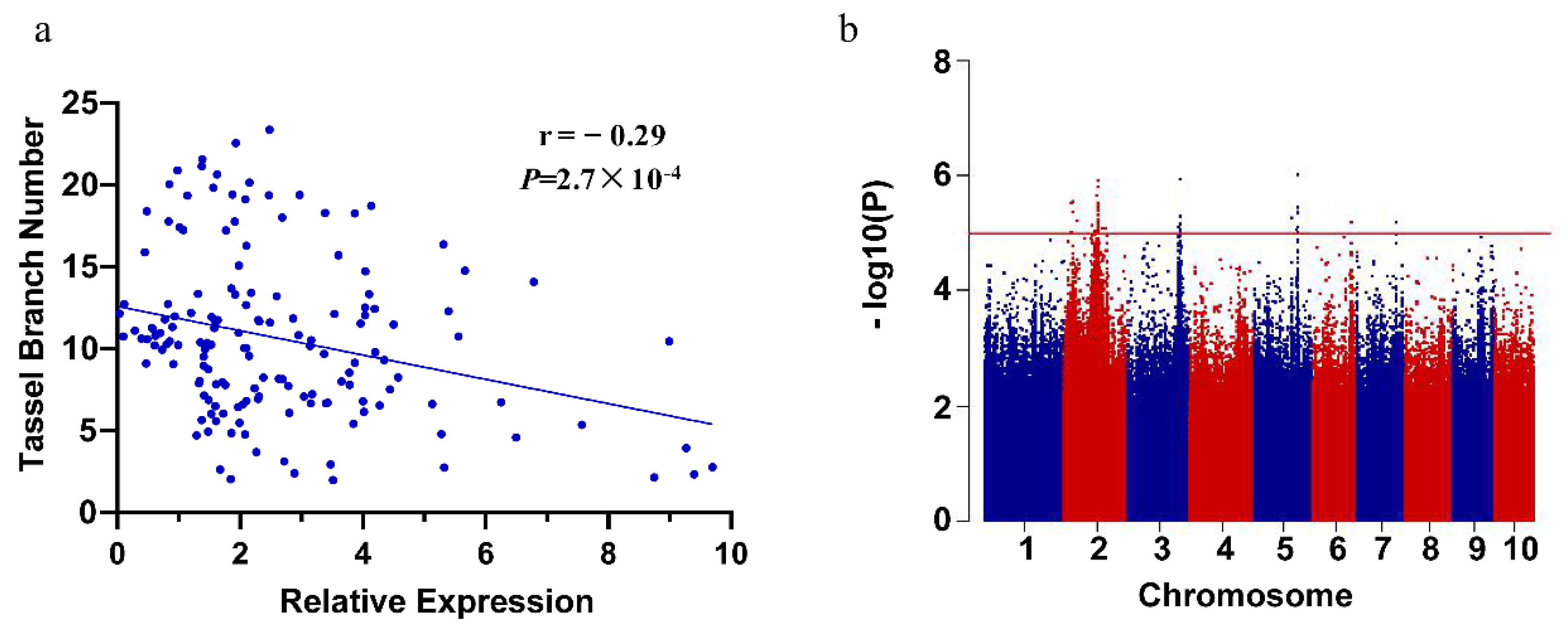
2.4. Expression Quantitative Trait Loci Analysis of ZmPAT7
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. QTL Mapping and Identification of Candidate Genes
4.3. Identification of the CRISPR/Cas9 Mutants of ZmPAT7
4.4. Candidate Gene-Based Association Analysis
4.5. Expression Quantitative Trait Loci Analysis
4.6. RNA Isolation and Quantitative Reverse Transcription PCR (qRT–PCR)
4.7. Sequence Blast and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duvick, D.N.; Cassman, K.G. Post-green revolution trends in yield potential of temperate maize in the north-central United States. Crop Sci. 1999, 39, 1622–1630. [Google Scholar] [CrossRef]
- Gage, J.L.; White, M.R.; Edwards, J.W.; Kaeppler, S.; de Leon, N. Selection Signatures Underlying Dramatic Male Inflorescence Transformation during Modern Hybrid Maize Breeding. Genetics 2018, 210, 1125–1138. [Google Scholar] [CrossRef] [Green Version]
- Duvick, D.N.; Smith, J.S.C.; Cooper, M.; Longper, M.; Smith, J.S.C.C. ng Modern Hybrid Maize Breeding. In Plant Breeding Reviews, Part 2: Long-Term Selection: Crops, Animals, and Bacteria; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004; Volume 24, pp. 109–151. [Google Scholar]
- Upadyayula, N.; da Silva, H.S.; Bohn, M.O.; Rocheford, T.R. Genetic and QTL analysis of maize tassel and ear inflorescence architecture. Theor. Appl. Genet. 2006, 112, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.B.; Lin, Z.C.; Li, X.; Zhao, Y.P.; Zhao, B.B.; Wu, G.X.; Ma, X.J.; Wang, H.; Xie, Y.R.; Li, Q.Q.; et al. Genome-wide selection and genetic improvement during modern maize breeding. Nat. Genet. 2020, 52, 565–571. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Peng, Y.L.; Zhang, J.W.; Fang, P.; Wu, B.Y. Mapping QTLs and meta-QTLs for two inflorescence architecture traits in multiple maize populations under different watering environments. Mol. Breed. 2017, 37, 91. [Google Scholar] [CrossRef]
- Li, M.F.; Zhong, W.S.; Yang, F.; Zhang, Z.X. Genetic and Molecular Mechanisms of Quantitative Trait Loci Controlling Maize Inflorescence Architecture. Plant Cell Physiol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.B.; Liu, H.; Liu, Z.P.; Dong, X.M.; Guo, J.J.; Li, W.; Chen, J.; Gao, C.; Zhu, Y.B.; Zheng, X.M.; et al. Identification of minor effect QTLs for plant architecture related traits using super high density genotyping and large recombinant inbred population in maize (Zea mays). BMC Plant Biol. 2018, 18, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.Y.; Hao, L.Y.; Kou, S.R.; Su, E.H.; Zhou, Y.Q.; Wang, R.L.; Mohamed, A.; Gao, C.X.; Zhang, D.F.; Li, Y.X.; et al. High-density quantitative trait locus mapping revealed genetic architecture of leaf angle and tassel size in maize. Mol. Breed. 2019, 39, 7. [Google Scholar] [CrossRef]
- Wang, Y.L.; Chen, J.; Guan, Z.R.; Zhang, X.X.; Zhang, Y.C.; Ma, L.L.; Yao, Y.M.; Peng, H.W.; Zhang, Q.; Zhang, B.; et al. Combination of multi-locus genome-wide association study and QTL mapping reveals genetic basis of tassel architecture in maize. Mol. Genet. Genom. 2019, 294, 1421–1440. [Google Scholar] [CrossRef]
- Brown, P.J.; Upadyayula, N.; Mahone, G.S.; Tian, F.; Bradbury, P.J.; Myles, S.; Holland, J.B.; Flint-Garcia, S.; McMullen, M.D.; Buckler, E.S.; et al. Distinct Genetic Architectures for Male and Female Inflorescence Traits of Maize. PLoS Genet. 2011, 7, e1002383. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Li, Y.X.; Shi, Y.S.; Song, Y.C.; Zhang, D.F.; Li, C.H.; Buckler, E.S.; Li, Y.; Zhang, Z.W.; Wang, T.Y. Joint-linkage mapping and GWAS reveal extensive genetic loci that regulate male inflorescence size in maize. Plant Biotechnol. J. 2016, 14, 1551–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortiri, E.; Chuck, G.; Vollbrecht, E.; Rocheford, T.; Martienssen, R.; Hake, S. ramosa2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell 2006, 18, 574–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh-Nagasawa, N.; Nagasawa, N.; Malcomber, S.; Sakai, H.; Jackson, D. A trehalose metabolic enzyme controls inflorescence architecture in maize. Nature 2006, 441, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Taguchi-Shiobara, F.; Yuan, Z.; Hake, S.; Jackson, D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Gene Dev. 2001, 15, 2755–2766. [Google Scholar] [CrossRef] [Green Version]
- Je, B.I.; Gruel, J.; Lee, Y.K.; Bommert, P.; Arevalo, E.D.; Eveland, A.L.; Wu, Q.Y.; Goldshmidt, A.; Meeley, R.; Bartlett, M.; et al. Signaling from maize organ primordia via FASCIATED EAR3 regulates stem cell proliferation and yield traits. Nat Genet. 2016, 48, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Whipple, C.; Jackson, D.; Hake, S. The maize SBP-box transcription factor encoded by tasselsheath4 regulates bract development and the establishment of meristem boundaries. Development 2010, 137, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.S.; Brown, P.J.; Meeley, R.; Hake, S. Maize SBP-box transcription factors unbranched2 and unbranched3 affect yield traits by regulating the rate of lateral primordia initiation. Proc. Natl. Acad. Sci. USA 2014, 111, 18775–18780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSteen, P.; Hake, S. barren inflorescence2 regulates axillary meristem development in the maize inflorescence. Development 2001, 128, 2881–2891. [Google Scholar] [CrossRef]
- Skirpan, A.; Culler, A.H.; Gallavotti, A.; Jackson, D.; Cohen, J.D.; McSteen, P. BARREN INFLORESCENCE2 Interaction with ZmPIN1a Suggests a Role in Auxin Transport during Maize Inflorescence Development. Plant Cell Physiol. 2009, 50, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.E.; Tian, S.K.; Zhang, W.L.; Dong, X.; Ma, C.X.; Wang, Y.; Yan, J.B.; Yue, B. Q(Dtbn1), an F-box gene affecting maize tassel branch number by a dominant model. Plant Biotechnol. J. 2021, 19, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Bomblies, K.; Wang, R.L.; Ambrose, B.A.; Schmidt, R.J.; Meeley, R.B.; Doebley, J. Duplicate FLORICAULA/LEAFY homologs zfl1 and zfl2 control inflorescence architecture and flower patterning in maize. Development 2003, 130, 2385–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemsley, P.A.; Kemp, A.C.; Grierson, C.S. The TIP GROWTH DEFECTIVE1 S-acyl transferase regulates plant cell growth in Arabidopsis. Plant Cell 2005, 17, 2554–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Lin, J.Z.; Peng, D.; Yang, Y.Z.; Guo, M.; Tang, D.Y.; Tan, X.F.; Liu, X.M. Plant architecture and grain yield are regulated by the novel DHHC-type zinc finger protein genes in rice (Oryza sativa L.). Plant Sci. 2017, 254, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.G.; Yan, Q.; Zhao, S.L.; He, F.; Xu, J.F.; Qi, B.X.; Zhang, Y.X. Knockout of the S-acyltransferase Gene, PbPAT14, Confers the Dwarf Yellowing Phenotype in First Generation Pear by ABA Accumulation. Int. J. Mol. Sci. 2019, 20, 6347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.M.; Mi, Y.; Mao, H.D.; Liu, S.X.; Chen, L.M.; Qin, F. Genetic variation in ZmTIP1 contributes to root hair elongation and drought tolerance in maize. Plant Biotechnol. J. 2020, 18, 1271–1283. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.W.; Zhang, S.Z.; Sun, M.H.; Liu, S.Y.; Qi, B.X.; Li, X.Z. Putative DHHC-Cysteine-Rich Domain S-Acyltransferase in Plants. PLoS ONE 2013, 8, e75985. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.L.; Fu, J.J.; Zong, N.; Wang, J.; Song, D.D.; Zhang, X.; He, C.; Fang, T.; Zhang, H.W.; Fan, Y.L.; et al. Kernel size-related genes revealed by an integrated eQTL analysis during early maize kernel development. Plant J. 2019, 98, 19–32. [Google Scholar] [CrossRef]
- Fu, J.J.; Cheng, Y.B.; Linghu, J.J.; Yang, X.H.; Kang, L.; Zhang, Z.X.; Zhang, J.; He, C.; Du, X.M.; Peng, Z.Y.; et al. RNA sequencing reveals the complex regulatory network in the maize kernel. Nat. Commun. 2013, 4, 3832. [Google Scholar] [CrossRef] [Green Version]
- Berke, T.G.; Rocheford, T.R. Quantitative trait loci for tassel traits in maize. Crop Sci. 1999, 39, 1439–1443. [Google Scholar] [CrossRef]
- Mickelson, S.M.; Stuber, C.S.; Senior, L.; Kaeppler, S.M. Quantitative trait loci controlling leaf and tassel traits in a B73 x MO17 population of maize. Crop Sci. 2002, 42, 1902–1909. [Google Scholar] [CrossRef]
- Chen, Z.L.; Wang, B.B.; Dong, X.M.; Liu, H.; Ren, L.H.; Chen, J.; Hauck, A.; Song, W.B.; Lai, J.S. An ultra-high density bin-map for rapid QTL mapping for tassel and ear architecture in a large F2 maize population. BMC Genom. 2014, 15, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.J.; Yang, C.; Tang, D.G.; Zhang, L.; Zhang, L.; Qu, J.T.; Liu, J. Dissection of the genetic architecture for tassel branch number by QTL analysis in two related populations in maize. J. Integr. Agric. 2017, 16, 1432–1442. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Lu, Y.L.; Yang, X.H.; Huang, J.; Zhou, Y.; Ali, F.; Wen, W.W.; Liu, J.; Li, J.S.; Yan, J.B. Genome Wide Association Studies Using a New Nonparametric Model Reveal the Genetic Architecture of 17 Agronomic Traits in an Enlarged Maize Association Panel. PLoS Genet. 2014, 10, e1004573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.Y.; Liu, Q.; Li, X.; Yang, J.L.; Liu, S.Z.; Huang, Y.L.; Scanlon, M.J.; Nettleton, D.; Schnable, P.S. Substantial contribution of genetic variation in the expression of transcription factors to phenotypic variation revealed by eRD-GWAS. Genome Biol. 2017, 18, 192. [Google Scholar] [CrossRef]
- Hemsley, P.A.; Weimar, T.; Lilley, K.S.; Dupree, P.; Grierson, C.S. A proteomic approach identifies many novel palmitoylated proteins in Arabidopsis. New Phytol. 2013, 197, 805–814. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Tan, X.F.; Zhang, L.; Yuan, D.Y.; Lin, J.Z.; Liu, X.M.; Jiang, Y.Q.; Zhou, B. Increasing branch and seed yield through heterologous expression of the novel rice S-acyl transferase gene OsPAT15 in Brassica napus L. Breed. Sci. 2018, 68, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.B.; Yu, B.Y.; Cao, Z.D.; Chen, Y.M.; Wu, Q.; Huang, J.Y.; Yang, C.W. Two homologous protein S-acyltransferases, PAT13 and PAT14, cooperatively regulate leaf senescence in Arabidopsis. J. Exp. Bot. 2015, 66, 6345–6353. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Scott, R.; Doughty, J.; Grant, M.; Qi, B.X. Protein S-Acyltransferase 14: A Specific Role for Palmitoylation in Leaf Senescence in Arabidopsis. Plant Physiol. 2016, 170, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, B.X.; Doughty, J.; Hooley, R. A Golgi and tonoplast localized S-acyl transferase is involved in cell expansion, cell division, vascular patterning and fertility in Arabidopsis. New Phytol. 2013, 200, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.Z.; Li, S.; Feng, Q.N.; Zhang, Y.L.; Zhao, X.Y.; Zeng, Y.L.; Wang, H.; Jiang, L.W.; Zhang, Y. PROTEIN S-ACYL TRANSFERASE10 is Critical for Development and Salt Tolerance in Arabidopsis. Plant Cell 2013, 25, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.Y.; Chai, S.; Ge, F.R.; Feng, Q.N.; Zhang, Y.; Li, S. Arabidopsis PROTEIN S-ACYL TRANSFERASE4 mediates root hair growth. Plant J. 2017, 90, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.X.; Li, H.J.; Morgan, C.; Bomblies, K.; Yang, W.C.; Qi, B.X. Both male and female gametogenesis require a fully functional protein S-acyl transferase 21 in Arabidopsis thaliana. Plant J. 2019, 100, 754–767. [Google Scholar] [CrossRef]
- Li, C.H.; Guan, H.H.; Jing, X.; Li, Y.Y.; Wang, B.B.; Li, Y.X.; Liu, X.Y.; Zhang, D.F.; Liu, C.; Xie, X.Q.; et al. Genomic Insights into Historical Improvement of Heterotic Groups during Modern Hybrid Maize Breeding; to be submitted; Institute of Crop Science, Chinese Academy of Agricultural Sciences: Beijing, China, 2022. [Google Scholar]
- Li, C.H.; Li, Y.X.; Bradbury, P.J.; Wu, X.; Shi, Y.S.; Song, Y.C.; Zhang, D.F.; Rodgers-Melnick, E.; Buckler, E.S.; Zhang, Z.W.; et al. Construction of high-quality recombination maps with low-coverage genomic sequencing for joint linkage analysis in maize. BMC Biol. 2015, 13, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5 Department of Statistics; North Carolina State University: Raleigh, NC, USA, 2012; Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 15 May 2020).
- Liu, H.J.; Jian, L.M.; Xu, J.T.; Zhang, Q.H.; Zhang, M.L.; Jin, M.L.; Peng, Y.; Yan, J.L.; Han, B.Z.; Liu, J.; et al. High-Throughput CRISPR/Cas9 Mutagenesis Streamlines Trait Gene Identification in Maize. Plant Cell 2020, 32, 1397–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chow, C.N.; Lee, T.Y.; Hung, Y.C.; Li, G.Z.; Tseng, K.C.; Liu, Y.H.; Kuo, P.L.; Zheng, H.Q.; Chang, W.C. PlantPAN3.0: A new and updated resource for reconstructing transcriptional regulatory networks from ChIP-seq experiments in plants. Nucleic Acids Res. 2019, 47, D1155–D1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL | Chr | Peaks (cM) | Interval (cM) | LOD | Interval (Mb) | Add | R2 (%) | Number of Genes in the Interval |
|---|---|---|---|---|---|---|---|---|
| qTBN2a | 2 | 30.12 | 17.25–37.37 | 4.24 | 7.85–14.72 | −0.56 | 6.08 | 571 |
| qTBN2b | 2 | 192.7 | 187.05–199.07 | 3.69 | 233.63–236.44 | 0.56 | 5.21 | 217 |
| qTBN4 | 4 | 68.09 | 61.14–77.39 | 10.47 | 58.51–134.62 | 0.93 | 16.22 | 2510 |
| qTBN6 | 6 | 104.6 | 99.09–109.45 | 6.47 | 152.15–156.88 | −0.7 | 9.53 | 413 |
| QTL | Tag SNPs | Chr | Pos (bp) | p Value | Candidate Genes That near the Tag SNPs |
|---|---|---|---|---|---|
| qTBN2a | S2_8809018 | 2 | 8,809,018 | 3.77 × 10−10 | GRMZM2G002559; GRMZM2G302701; GRMZM2G302712 |
| S2_8889278 | 2 | 8,889,278 | 3.31 × 10−9 | GRMZM5G897776 | |
| S2_9443046 | 2 | 9,443,046 | 1.09 × 10−10 | GRMZM2G080041; GRMZM2G080054 | |
| S2_11829845 | 2 | 11,829,845 | 5.70 × 10−10 | GRMZM2G392125; GRMZM2G091118; GRMZM2G090872 | |
| S2_12687587 | 2 | 12,687,587 | 5.61 × 10−11 | GRMZM2G149556 | |
| S2_12691259 | 2 | 12,691,259 | 7.73 × 10−10 | GRMZM2G149556 | |
| S2_12798388 | 2 | 12,798,388 | 9.40 × 10−11 | GRMZM2G024898 | |
| S2_12898426 | 2 | 12,898,426 | 2.16 × 10−11 | GRMZM2G463280; GRMZM2G463267; GRMZM2G162266 | |
| S2_13299879 | 2 | 13,299,879 | 4.32 × 10−10 | GRMZM2G038722; GRMZM2G038714; GRMZM2G342039 | |
| S2_13636982 | 2 | 13,636,982 | 4.49 × 10−10 | ||
| S2_13638186 | 2 | 13,638,186 | 8.50 × 10−9 | GRMZM2G090332 | |
| S2_14264763 | 2 | 14,264,763 | 3.38 × 10−9 | GRMZM2G163067 | |
| S2_14267757 | 2 | 14,267,757 | 3.8 × 10−10 | GRMZM2G163067 | |
| S2_14467324 | 2 | 14,467,324 | 1.38 × 10−10 | GRMZM2G146866; GRMZM2G146847; GRMZM2G010011 | |
| S2_14541295 | 2 | 14,541,295 | 2.43 × 10−10 | GRMZM2G314396; GRMZM2G017197; GRMZM2G314386; GRMZM2G314412 | |
| qTBN2b | S2_233632388 | 2 | 233,632,388 | 5.21 × 10−10 | GRMZM2G176347; GRMZM2G176375 |
| S2_234531325 | 2 | 234,531,325 | 1.94 × 10−10 | GRMZM2G423640; GRMZM2G423636 | |
| S2_234707306 | 2 | 234,707,306 | 1.46 × 10−9 | GRMZM2G324507 | |
| S2_235802795 | 2 | 235,802,795 | 1.03 × 10−10 | GRMZM2G347995 | |
| qTBN4 | S4_66857201 | 4 | 66,857,201 | 3.35 × 10−10 | GRMZM2G399421 |
| S4_66908736 | 4 | 66,908,736 | 1.39 × 10−10 | GRMZM2G030628 | |
| S4_71986141 | 4 | 71,986,141 | 7.14 × 10−12 | GRMZM2G448456 | |
| PZE-104061279 | 4 | 120,847,621 | 3.38 × 10−6 | ||
| S4_122747978 | 4 | 122,747,978 | 1.53 × 10−12 | ||
| qTBN6 | S6_153206873 | 6 | 153,206,873 | 2.97 × 10−12 | GRMZM2G359892 |
| Code of eQTL | Peak_SNP | p_Value | V4_Gene_Id | Annotation |
|---|---|---|---|---|
| eQTL-1 | 2:26,517,652 | 3.05 × 10−6 | Zm00001d002917 | RNA-binding region RNP-1 |
| eQTL-2 | 2:33,010,637 | 4.51 × 10−6 | Zm00001d003123 | NA |
| Zm00001d003124 | DNA glycosylase superfamily protein | |||
| Zm00001d003125 | Uncharacterized protein | |||
| eQTL-3 | 2:52,125,742 | 6.25 × 10−6 | ||
| eQTL-4 | 2:104,153,316 | 7.62 × 10−6 | Zm00001d004336 | NA |
| eQTL-5 | 2:115,870,588 | 9.34 × 10−6 | Zm00001d004509 | Terpene synthase 7 |
| Zm00001d004512 | Transcription factor E2FC | |||
| Zm00001d004513 | Bifunctional 3-phosphoadenosine 5-phosphosulfate synthetase 2 | |||
| eQTL-6 | 2:122,415,924 | 2.26 × 10−6 | Zm00001d004619 | NA |
| eQTL-7 | 2:131,987,662 | 1.27 × 10−6 | Zm00001d004697 | IAA27-auxin-responsive Aux/IAA family member |
| Zm00001d004698 | Serine–threonine-protein kinase-like protein CCR4 | |||
| eQTL-8 | 2:147,699,071 | 8.28 × 10−6 | Zm00001d004897 | Basic-leucine zipper (bZIP) transcription factor family protein |
| eQTL-9 | 2:158,878,809 | 8.32 × 10−6 | Zm00001d005112 | SAC3 family protein B |
| eQTL-10 | 3:194,080,838 | 8.12 × 10−6 | Zm00001d043273 | SR protein related family member |
| Zm00001d043274 | Insulin-degrading enzyme-like 1 peroxisomal | |||
| Zm00001d043275 | Vacuole membrane protein KMS1 | |||
| eQTL-11 | 3:200,375,447 | 9.92 × 10−6 | Zm00001d043444 | plant-specific domain TIGR01615 family protein expressed |
| Zm00001d043445 | Dihydroorotase | |||
| Zm00001d043446 | Uncharacterized protein | |||
| Zm00001d043447 | PP2A | |||
| Zm00001d043449 | RNA-binding S4 domain-containing protein | |||
| Zm00001d043450 | DNA-binding WRKY | |||
| Zm00001d043451 | Dynamin-related protein 5A | |||
| Zm00001d043452 | Hox2a protein | |||
| Zm00001d043453 | Early nodulin-like protein 3 | |||
| Zm00001d043454 | Uncharacterized protein | |||
| eQTL-12 | 3:200,789,018 | 8.80 × 10−6 | Zm00001d043461 | ZCN12 |
| Zm00001d043462 | Uncharacterized protein | |||
| Zm00001d043463 | Protein NRT1/PTR FAMILY 4.2 | |||
| Zm00001d043464 | Oil body-associated protein 2A | |||
| Zm00001d043465 | 50S ribosomal protein L13 chloroplastic | |||
| eQTL-13 | 5:143,694,885 | 5.49 × 10−6 | Zm00001d016083 | NA |
| Zm00001d016084 | Pre-mRNA-splicing factor 3 | |||
| Zm00001d016085 | Putative uncharacterized protein | |||
| eQTL-14 | 5:163,991,516 | 8.74 × 10−6 | Zm00001d016477 | Eukaryotic aspartyl protease family protein |
| eQTL-15 | 5:170,985,132 | 1.02 × 10−6 | Zm00001d016636 | Putative uncharacterized protein |
| Zm00001d016637 | NA | |||
| eQTL-16 | 6:151,600,488 | 6.51 × 10−6 | Zm00001d038205 | MLO-like protein |
| Zm00001d038206 | Pentatricopeptide repeat-containing protein mitochondrial | |||
| Zm00001d038207 | ZmNAC3, NAC domain-containing protein 35 | |||
| eQTL-17 | 7:145,022,165 | 6.56 × 10−6 | Zm00001d021172 | NA |
| Zm00001d021173 | Formin-like protein 1 | |||
| Zm00001d021176 | Disease resistance protein RPM1 | |||
| Zm00001d021177 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, H.; Chen, X.; Wang, K.; Liu, X.; Zhang, D.; Li, Y.; Song, Y.; Shi, Y.; Wang, T.; Li, C.; et al. Genetic Variation in ZmPAT7 Contributes to Tassel Branch Number in Maize. Int. J. Mol. Sci. 2022, 23, 2586. https://doi.org/10.3390/ijms23052586
Guan H, Chen X, Wang K, Liu X, Zhang D, Li Y, Song Y, Shi Y, Wang T, Li C, et al. Genetic Variation in ZmPAT7 Contributes to Tassel Branch Number in Maize. International Journal of Molecular Sciences. 2022; 23(5):2586. https://doi.org/10.3390/ijms23052586
Chicago/Turabian StyleGuan, Honghui, Xiaojing Chen, Kailiang Wang, Xuyang Liu, Dengfeng Zhang, Yongxiang Li, Yanchun Song, Yunsu Shi, Tianyu Wang, Chunhui Li, and et al. 2022. "Genetic Variation in ZmPAT7 Contributes to Tassel Branch Number in Maize" International Journal of Molecular Sciences 23, no. 5: 2586. https://doi.org/10.3390/ijms23052586
APA StyleGuan, H., Chen, X., Wang, K., Liu, X., Zhang, D., Li, Y., Song, Y., Shi, Y., Wang, T., Li, C., & Li, Y. (2022). Genetic Variation in ZmPAT7 Contributes to Tassel Branch Number in Maize. International Journal of Molecular Sciences, 23(5), 2586. https://doi.org/10.3390/ijms23052586