
Quantification of Farnesylated Progerin in Hutchinson-Gilford Progeria Patient Cells by Mass Spectrometry
 , , , and
, , , and
Abstract
1. Introduction
2. Results and Discussion
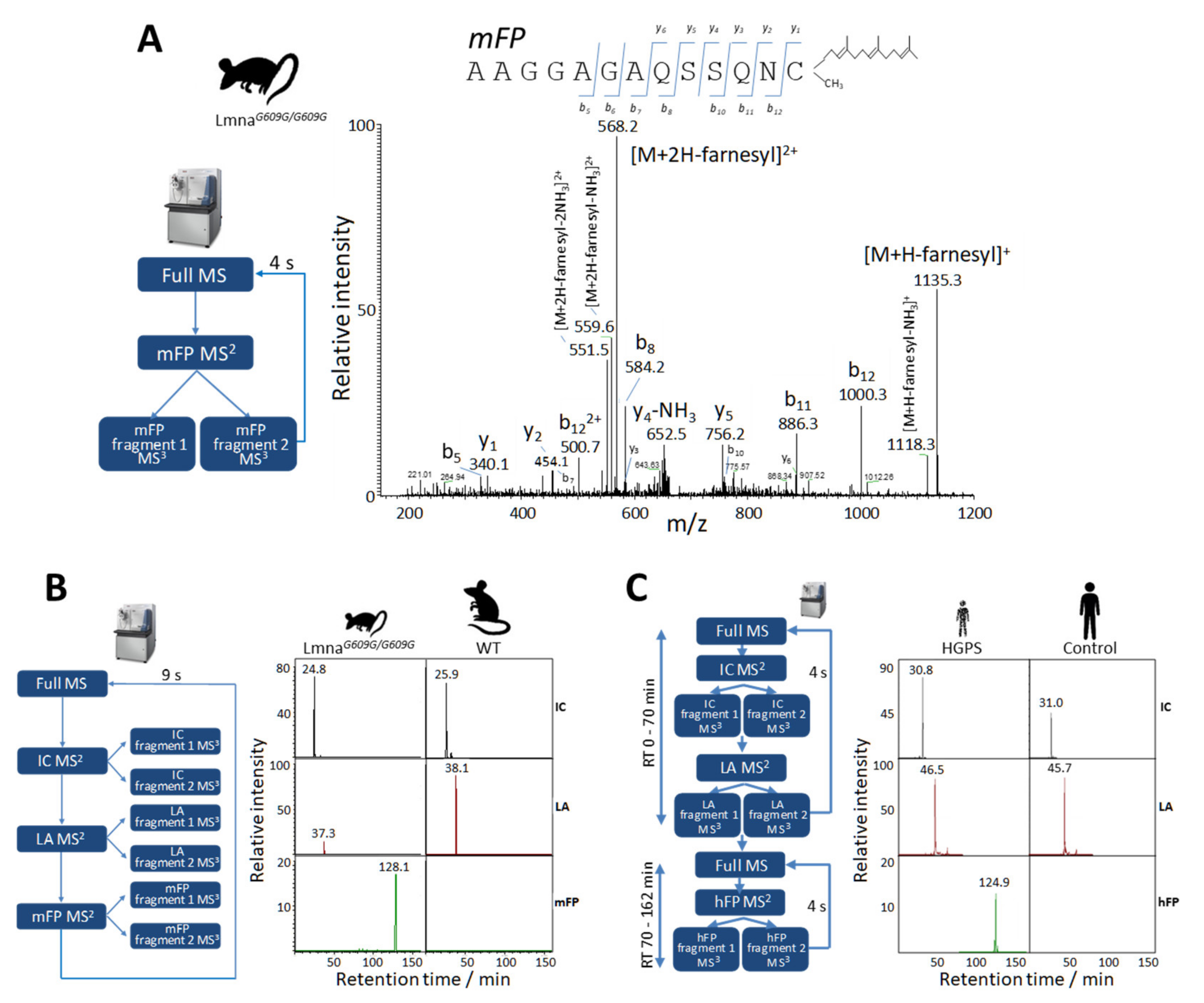
2.1. Identification of Lamin A peptides by Shotgun LC-MS/MS
2.2. Relative Quantification of Lamin A and Progerin in Mice Mouse Fibroblasts by Parallel Reaction Monitoring
2.3. Relative Quantification of Lamin A and Progerin in Human Fibroblasts by PRM
2.4. Absolute Quantification of Lamin A and Progerin in Human Samples by PRM
2.5. PRM Analysis of of Lamin A and Progerin in Blood from HGPS Patients
3. Materials and Methods
3.1. Biological Materials
3.1.1. Mouse Cells and Tissues
3.1.2. Human Cells
3.2. Synthetic Peptides
- Heavy internal control peptide, IC* (TVLCGTCGQPADK);
- Heavy lamin A peptide, LA* (SVGGSGGGSFGDNLVTR);
- Heavy human progerin farnesylated peptide, hFP* (ASASGSGAQSPQNC, with O-methylated, farnesylated Cys).
3.3. Preparation of Protein Extracts
3.3.1. Protein Extracts for LC-MS/MS Analysis
3.3.2. Protein Extracts for Western Blot Analysis
3.4. Protein Digestion
3.5. LC-MS/MS Analysis
3.5.1. Shotgun LC-MS/MS Analysis
3.5.2. Targeted PRM LC-MS/MS Assays
Targeted PRM LC-MS/MS Assay for the Relative Quantification of Lamin A and Progerin in Mouse Samples
Targeted PRM LC-MS/MS Assays for Relative or Absolute Quantification of Lamin A and Progerin in Human Samples
3.6. Protein Identification and LC-MS/MS Data Analysis
3.7. Western Blot Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dorado, B.; Andres, V. A-type lamins and cardiovascular disease in premature aging syndromes. Curr. Opin. Cell Biol. 2017, 46, 17–25. [Google Scholar] [CrossRef]
- Gordon, L.B.; Shappell, H.; Massaro, J.; D’Agostino, R.B., Sr.; Brazier, J.; Campbell, S.E.; Kleinman, M.E.; Kieran, M.W. Association of Lonafarnib Treatment vs No Treatment With Mortality Rate in Patients With Hutchinson-Gilford Progeria Syndrome. JAMA 2018, 319, 1687–1695. [Google Scholar] [CrossRef]
- De Sandre-Giovannoli, A.; Bernard, R.; Cau, P.; Navarro, C.; Amiel, J.; Boccaccio, I.; Lyonnet, S.; Stewart, C.L.; Munnich, A.; Le Merrer, M.; et al. Lamin a truncation in Hutchinson-Gilford progeria. Science 2003, 300, 2055. [Google Scholar] [CrossRef]
- Eriksson, M.; Brown, W.T.; Gordon, L.B.; Glynn, M.W.; Singer, J.; Scott, L.; Erdos, M.R.; Robbins, C.M.; Moses, T.Y.; Berglund, P.; et al. Recurrent de novo point mutations in lamin A cause Hutchinson-Gilford progeria syndrome. Nature 2003, 423, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Hamczyk, M.R.; del Campo, L.; Andres, V. Aging in the Cardiovascular System: Lessons from Hutchinson-Gilford Progeria Syndrome. Annu. Rev. Physiol. 2018, 80, 27–48. [Google Scholar] [CrossRef]
- Reunert, J.; Wentzell, R.; Walter, M.; Jakubiczka, S.; Zenker, M.; Brune, T.; Rust, S.; Marquardt, T. Neonatal progeria: Increased ratio of progerin to lamin A leads to progeria of the newborn. Eur. J. Hum. Genet. 2012, 20, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Lopez, A.; Espinos-Estevez, C.; Gonzalez-Gomez, C.; Gonzalo, P.; Andres-Manzano, M.J.; Fanjul, V.; Riquelme-Borja, R.; Hamczyk, M.R.; Macias, A.; Del Campo, L.; et al. Cardiovascular Progerin Suppression and Lamin A Restoration Rescue Hutchinson-Gilford Progeria Syndrome. Circulation 2021, 144, 1777–1794. [Google Scholar] [CrossRef] [PubMed]
- Gordon, L.B.; Kleinman, M.E.; Massaro, J.; D’Agostino, R.B., Sr.; Shappell, H.; Gerhard-Herman, M.; Smoot, L.B.; Gordon, C.M.; Cleveland, R.H.; Nazarian, A.; et al. Clinical Trial of the Protein Farnesylation Inhibitors Lonafarnib, Pravastatin, and Zoledronic Acid in Children With Hutchinson-Gilford Progeria Syndrome. Circulation 2016, 134, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Ramiro, B.; Gil-Ordonez, A.; Marin-Ramos, N.I.; Ortega-Nogales, F.J.; Balabasquer, M.; Gonzalo, P.; Khiar-Fernandez, N.; Rolas, L.; Barkaway, A.; Nourshargh, S.; et al. Isoprenylcysteine Carboxylmethyltransferase-Based Therapy for Hutchinson-Gilford Progeria Syndrome. ACS Cent. Sci. 2021, 7, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Hisama, F.M.; Lessel, D.; Leistritz, D.; Friedrich, K.; McBride, K.L.; Pastore, M.T.; Gottesman, G.S.; Saha, B.; Martin, G.M.; Kubisch, C.; et al. Coronary artery disease in a Werner syndrome-like form of progeria characterized by low levels of progerin, a splice variant of lamin A. Am. J. Med. Genet. A 2011, 155A, 3002–3006. [Google Scholar] [CrossRef]
- Lopez-Mejia, I.C.; Vautrot, V.; De Toledo, M.; Behm-Ansmant, I.; Bourgeois, C.F.; Navarro, C.L.; Osorio, F.G.; Freije, J.M.; Stevenin, J.; De Sandre-Giovannoli, A.; et al. A conserved splicing mechanism of the LMNA gene controls premature aging. Hum. Mol. Genet. 2011, 20, 4540–4555. [Google Scholar] [CrossRef] [PubMed]
- Berndt, N.; Hamilton, A.D.; Sebti, S.M. Targeting protein prenylation for cancer therapy. Nat. Rev. Cancer 2011, 11, 775–791. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.H.; Del Toro, R.; Rivera-Torres, J.; Rak, J.; Korn, C.; Garcia-Garcia, A.; Macias, D.; Gonzalez-Gomez, C.; Del Monte, A.; Wittner, M.; et al. Remodeling of Bone Marrow Hematopoietic Stem Cell Niches Promotes Myeloid Cell Expansion during Premature or Physiological Aging. Cell Stem Cell 2019, 25, 407–418.e6. [Google Scholar] [CrossRef]
- Giannakouros, T.; Armstrong, J.; Magee, A.I. Protein prenylation in Schizosaccharomyces pombe. FEBS Lett. 1992, 297, 103–106. [Google Scholar] [CrossRef]
- Chan, L.N.; Hart, C.; Guo, L.; Nyberg, T.; Davies, B.S.; Fong, L.G.; Young, S.G.; Agnew, B.J.; Tamanoi, F. A novel approach to tag and identify geranylgeranylated proteins. Electrophoresis 2009, 30, 3598–3606. [Google Scholar] [CrossRef] [PubMed]
- Dursina, B.; Reents, R.; Delon, C.; Wu, Y.; Kulharia, M.; Thutewohl, M.; Veligodsky, A.; Kalinin, A.; Evstifeev, V.; Ciobanu, D.; et al. Identification and specificity profiling of protein prenyltransferase inhibitors using new fluorescent phosphoisoprenoids. J. Am. Chem. Soc. 2006, 128, 2822–2835. [Google Scholar] [CrossRef]
- Kho, Y.; Kim, S.C.; Jiang, C.; Barma, D.; Kwon, S.W.; Cheng, J.; Jaunbergs, J.; Weinbaum, C.; Tamanoi, F.; Falck, J.; et al. A tagging-via-substrate technology for detection and proteomics of farnesylated proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 12479–12484. [Google Scholar] [CrossRef]
- Nguyen, U.T.; Guo, Z.; Delon, C.; Wu, Y.; Deraeve, C.; Franzel, B.; Bon, R.S.; Blankenfeldt, W.; Goody, R.S.; Waldmann, H.; et al. Analysis of the eukaryotic prenylome by isoprenoid affinity tagging. Nat. Chem. Biol. 2009, 5, 227–235. [Google Scholar] [CrossRef]
- Doll, S.; Burlingame, A.L. Mass spectrometry-based detection and assignment of protein posttranslational modifications. ACS Chem. Biol. 2015, 10, 63–71. [Google Scholar] [CrossRef]
- Bronsema, K.J.; Bischoff, R.; van de Merbel, N.C. Internal standards in the quantitative determination of protein biopharmaceuticals using liquid chromatography coupled to mass spectrometry. J. Chromatogr. B 2012, 893–894, 1–14. [Google Scholar] [CrossRef]
- McClure, M.; DeLucas, L.J.; Wilson, L.; Ray, M.; Rowe, S.M.; Wu, X.; Dai, Q.; Hong, J.S.; Sorscher, E.J.; Kappes, J.C.; et al. Purification of CFTR for mass spectrometry analysis: Identification of palmitoylation and other post-translational modifications. Protein Eng. Des. Sel. 2012, 25, 7–14. [Google Scholar] [CrossRef]
- Sorek, N.; Akerman, A.; Yalovsky, S. Analysis of protein prenylation and S-acylation using gas chromatography-coupled mass spectrometry. Methods Mol. Biol. 2013, 1043, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Appels, N.M.; Rosing, H.; Stephens, T.C.; Hughes, A.; Schellens, J.H.; Beijnen, J.H. Absolute quantification of farnesylated Ras levels in complex samples using liquid chromatography fractionation combined with tryptic digestion and electrospray tandem mass spectrometry. Anal. Biochem. 2006, 352, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wotske, M.; Wu, Y.; Wolters, D.A. Liquid chromatographic analysis and mass spectrometric identification of farnesylated peptides. Anal. Chem. 2012, 84, 6848–6855. [Google Scholar] [CrossRef] [PubMed]
- Varela, I.; Pereira, S.; Ugalde, A.P.; Navarro, C.L.; Suarez, M.F.; Cau, P.; Cadinanos, J.; Osorio, F.G.; Foray, N.; Cobo, J.; et al. Combined treatment with statins and aminobisphosphonates extends longevity in a mouse model of human premature aging. Nat. Med. 2008, 14, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Palsuledesai, C.C.; Ochocki, J.D.; Markowski, T.W.; Distefano, M.D. A combination of metabolic labeling and 2D-DIGE analysis in response to a farnesyltransferase inhibitor facilitates the discovery of new prenylated proteins. Mol. Biosyst. 2014, 10, 1094–1103. [Google Scholar] [CrossRef]
- Varela, I.; Cadinanos, J.; Pendas, A.M.; Gutierrez-Fernandez, A.; Folgueras, A.R.; Sanchez, L.M.; Zhou, Z.; Rodriguez, F.J.; Stewart, C.L.; Vega, J.A.; et al. Accelerated ageing in mice deficient in Zmpste24 protease is linked to p53 signalling activation. Nature 2005, 437, 564–568. [Google Scholar] [CrossRef]
- Tevethia, M.J.; Ozer, H.L. SV40-mediated immortalization. Methods Mol. Biol. 2001, 165, 185–199. [Google Scholar] [CrossRef]
- Dignam, J.D.; Lebovitz, R.M.; Roeder, R.G. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983, 11, 1475–1489. [Google Scholar] [CrossRef]
- Bonzon-Kulichenko, E.; Perez-Hernandez, D.; Nunez, E.; Martinez-Acedo, P.; Navarro, P.; Trevisan-Herraz, M.; Ramos Mdel, C.; Sierra, S.; Martinez-Martinez, S.; Ruiz-Meana, M.; et al. A robust method for quantitative high-throughput analysis of proteomes by 18O labeling. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef]
- Martinez-Bartolome, S.; Navarro, P.; Martin-Maroto, F.; Lopez-Ferrer, D.; Ramos-Fernandez, A.; Villar, M.; Garcia-Ruiz, J.P.; Vazquez, J. Properties of average score distributions of SEQUEST: The probability ratio method. Mol. Cell. Proteom. 2008, 7, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Navarro, P.; Vazquez, J. A refined method to calculate false discovery rates for peptide identification using decoy databases. J. Proteom. Res. 2009, 8, 1792–1796. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Amino Acid Sequence | MS2 m/z (Charge) | MS3 m/z (Ion) | MS3 XIC Trace |
|---|---|---|---|---|---|
| IC | Internal control peptide | TVLC#GTC#GQPADK | 703.8238 (+2) | 430.2296 (y4) | 430.2296 > 333.1769 (y3) + 603.7657 > 933.4095 (y9) |
| 603.7657 (y112+) | |||||
| LA | Mature lamin A peptide | SVGGSGGGSFGDNLVTR | 783.8791 (+2) | 690.8288 (y152+) | 690.8288 > 774.4104 (y7) + 774.4104 > 602.3620 (y5) |
| 774.4104 (y7) | |||||
| mFP | LmnaG609G/G609G mouse farnesylated peptide | AAGGAGAQSSQNCΔ,O | 670.3374 (+2) | 568.2435 (NL2+) | 568.2435 > 886.4013 (b11) + 1135.4797 > 993.4054 (y11) |
| 1135.4797 (NL) |
| RT Range | Symbol | Description | Amino Acid Sequence | MS2 m/z (Charge) | MS3 m/z (Ion) | MS3 XIC Trace |
|---|---|---|---|---|---|---|
| 0–70 min | IC | Internal control peptide | TVLC#GTC#GQPADK | 703.8238 (+2) | 430.2296 (y4) | 430.2296 > 333.1769 (y3) + 603.7657 > 933.4095 (y9) |
| 603.7657 (y112+) | ||||||
| LA | Mature lamin A peptide | SVGGSGGGSFGDNLVTR | 783.8791 (+2) | 690.8288 (y152+) | 690.8288 > 774.4104 (y7) + 774.4104 > 602.3620 (y5) | |
| 774.4104 (y7) | ||||||
| 70–162 min | hFP | Human farnesylated peptide | ASASGSGAQSPQNCΔ,O | 741.8665 (+2) | 639.7726 (NL2+) 1278.5379 (NL) | 639.7726 > 717.3162 (b9) + 1278.5379 > 1260.5273 (-H2O) |
| RT Range | Symbol | Description | Amino Acid Sequence | MS2 m/z (Charge) | MS2 XIC Trace |
|---|---|---|---|---|---|
| 0–35 min | IC | Internal control peptide | TVLC#GTC#GQPADK | 703.8238 (+2) | 703.8238 > 933.4095 (y9) + 703.8238 > 603.7657 (y112+) |
| IC* | Isotopically labeled IC | TVLC#GTC#GQPADK* | 707.8309 (+2) | 707.8309 > 941.4237 (y9) + 707.8309 > 607.7728 (y112+) | |
| 35–70 min | LA | Mature lamin A peptide | SVGGSGGGSFGDNLVTR | 783.8791 (+2) | 783.8791 > 690.8288 (y152+) + 783.8791 > 921.4789 (y8) |
| LA* | Isotopically labeled LA | SVGGSGGGSFGDNLVTR* | 788.8831 (+2) | 788.8831 > 695.8329 (y152+) + 788.8831 > 931.4871 (y8) | |
| 70–162 min | hFP | Human farnesylated peptide | ASASGSGAQSPQNCΔ,O | 741.8665 (+2) | 741.8665 > 1278.5379 (NL) + 741.8665 > 1029.4596 (b12) |
| hFP* | Isotopically labeled hFP | A*SASGSGAQSPQNCΔ,O | 743.8700 (+2) | 743.8701 > 1286.5521 (NL) + 743.8701 > 1033.4667 (b12) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camafeita, E.; Jorge, I.; Rivera-Torres, J.; Andrés, V.; Vázquez, J. Quantification of Farnesylated Progerin in Hutchinson-Gilford Progeria Patient Cells by Mass Spectrometry. Int. J. Mol. Sci. 2022, 23, 11733. https://doi.org/10.3390/ijms231911733
Camafeita E, Jorge I, Rivera-Torres J, Andrés V, Vázquez J. Quantification of Farnesylated Progerin in Hutchinson-Gilford Progeria Patient Cells by Mass Spectrometry. International Journal of Molecular Sciences. 2022; 23(19):11733. https://doi.org/10.3390/ijms231911733
Chicago/Turabian StyleCamafeita, Emilio, Inmaculada Jorge, José Rivera-Torres, Vicente Andrés, and Jesús Vázquez. 2022. "Quantification of Farnesylated Progerin in Hutchinson-Gilford Progeria Patient Cells by Mass Spectrometry" International Journal of Molecular Sciences 23, no. 19: 11733. https://doi.org/10.3390/ijms231911733
APA StyleCamafeita, E., Jorge, I., Rivera-Torres, J., Andrés, V., & Vázquez, J. (2022). Quantification of Farnesylated Progerin in Hutchinson-Gilford Progeria Patient Cells by Mass Spectrometry. International Journal of Molecular Sciences, 23(19), 11733. https://doi.org/10.3390/ijms231911733