
BBX24 Interacts with DELLA to Regulate UV-B-Induced Photomorphogenesis in Arabidopsis thaliana

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. UV-B Regulates the Abundance of DELLA Protein
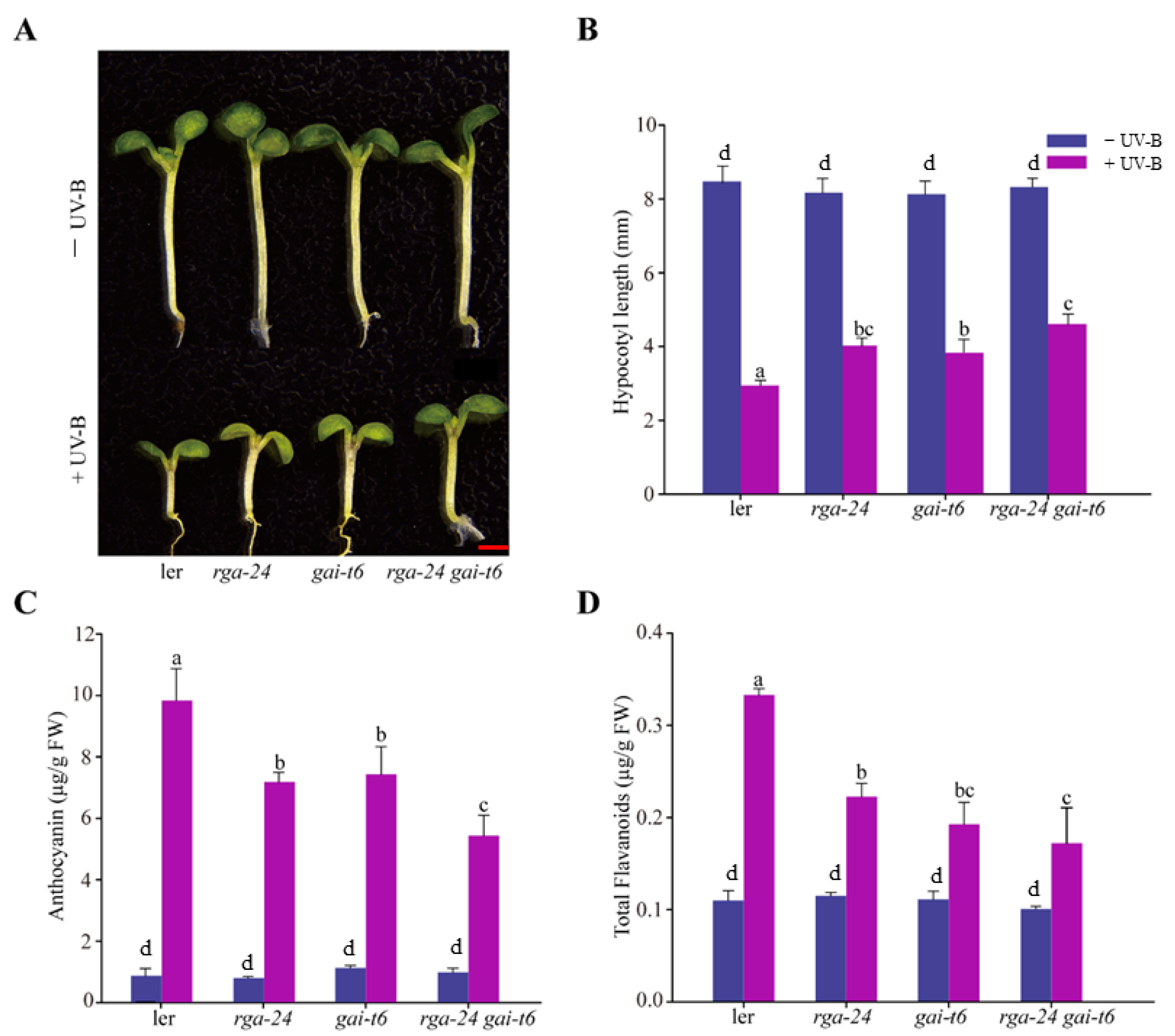
2.2. DELLA Positively Regulates UV-B-Induced Photomorphogenesis
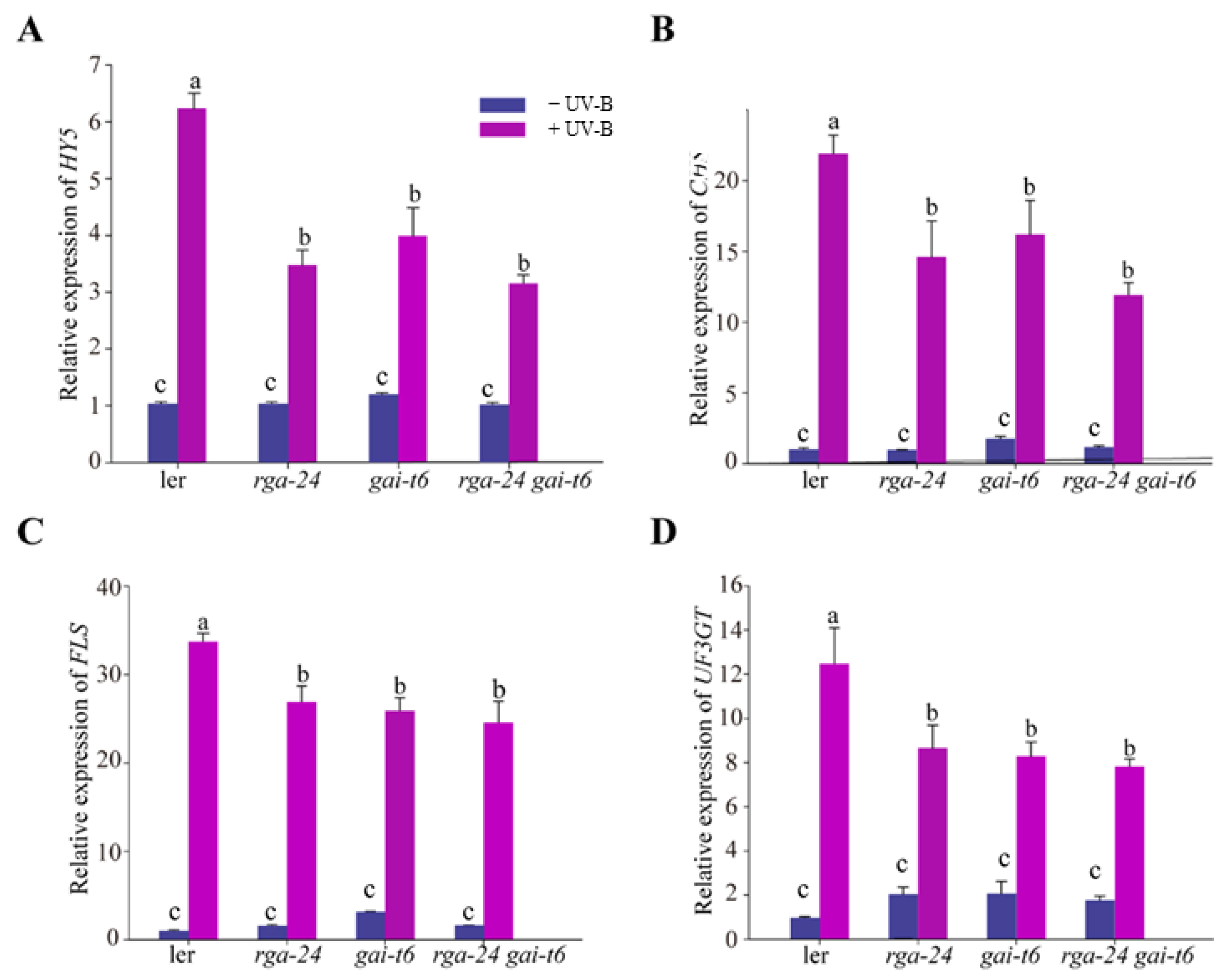
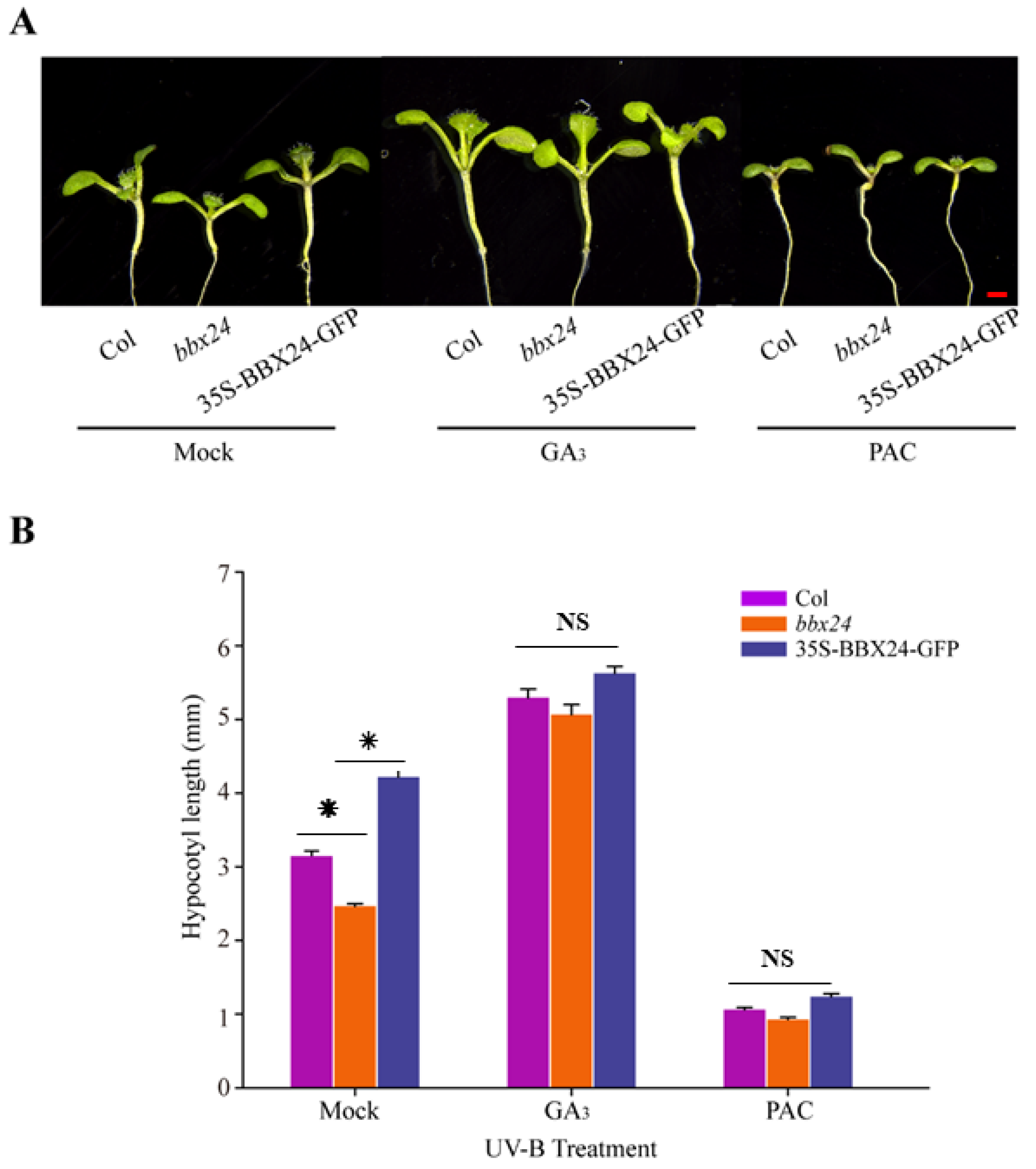
2.3. Antagonistic Regulation of DELLA and BBX24 in UV-B Photomorphogenesis
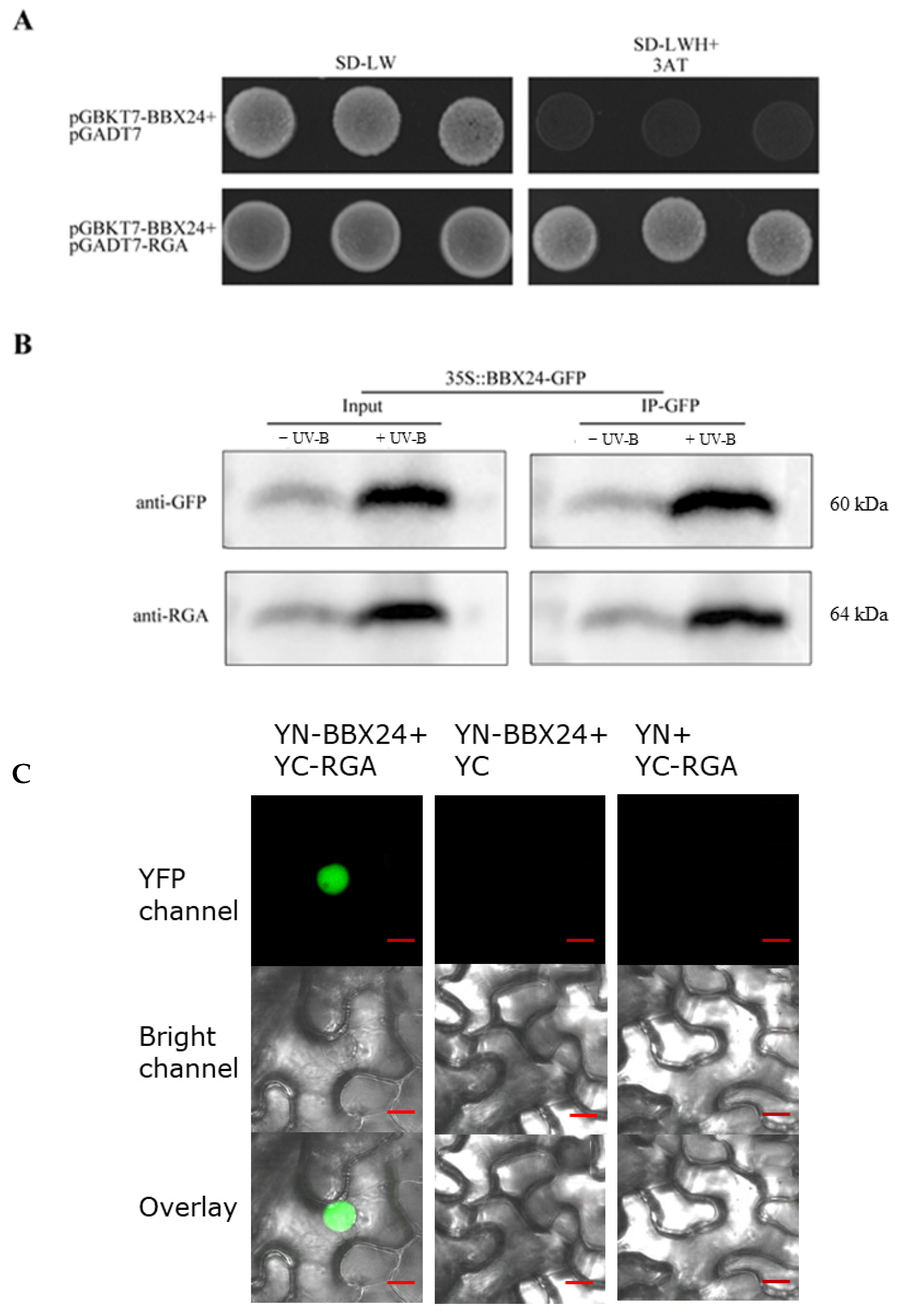
2.4. RGA Physically Interacts with BBX24 Protein to Regulate UV-B Signaling
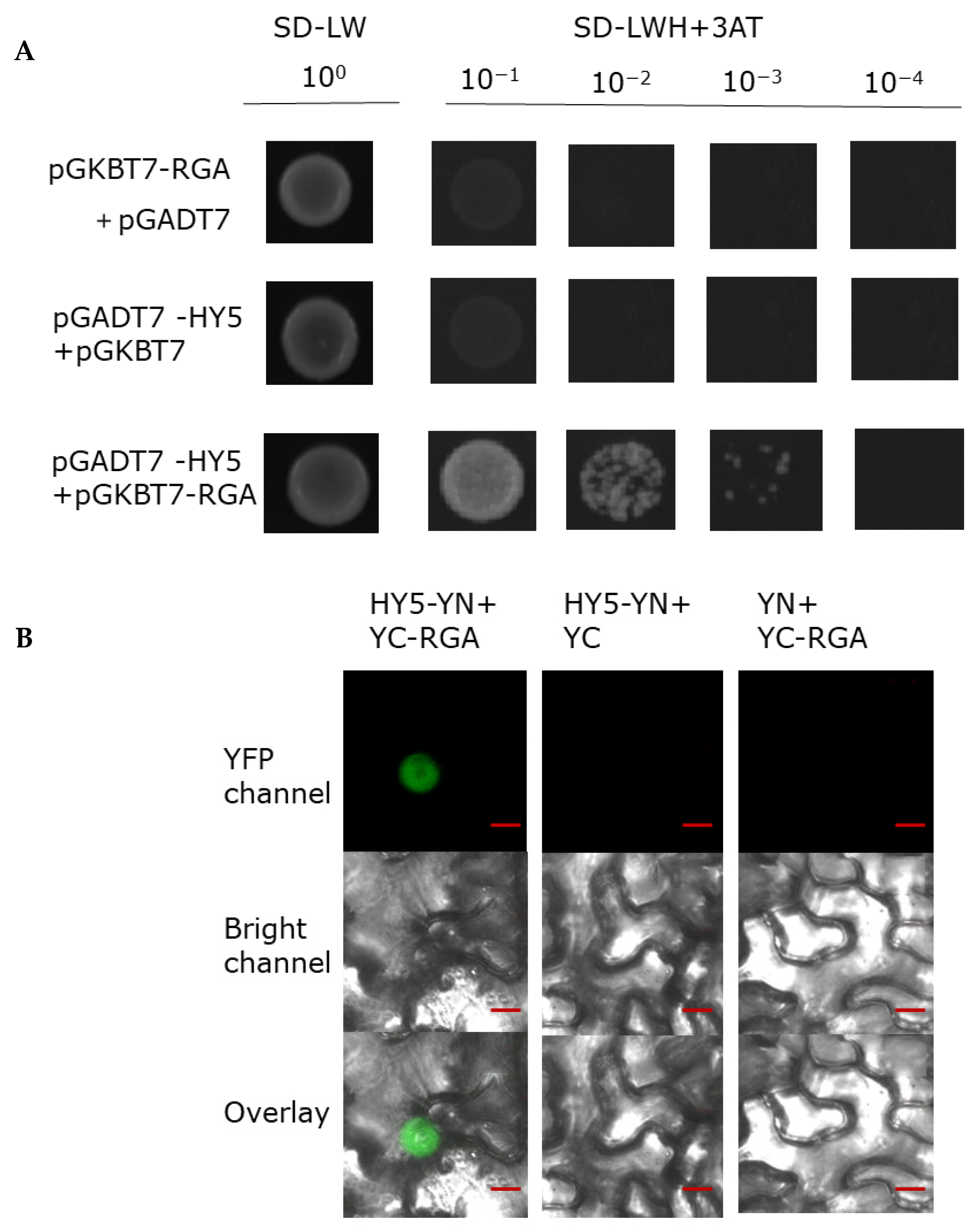
2.5. HY5 Physically Interacts with RGA Protein
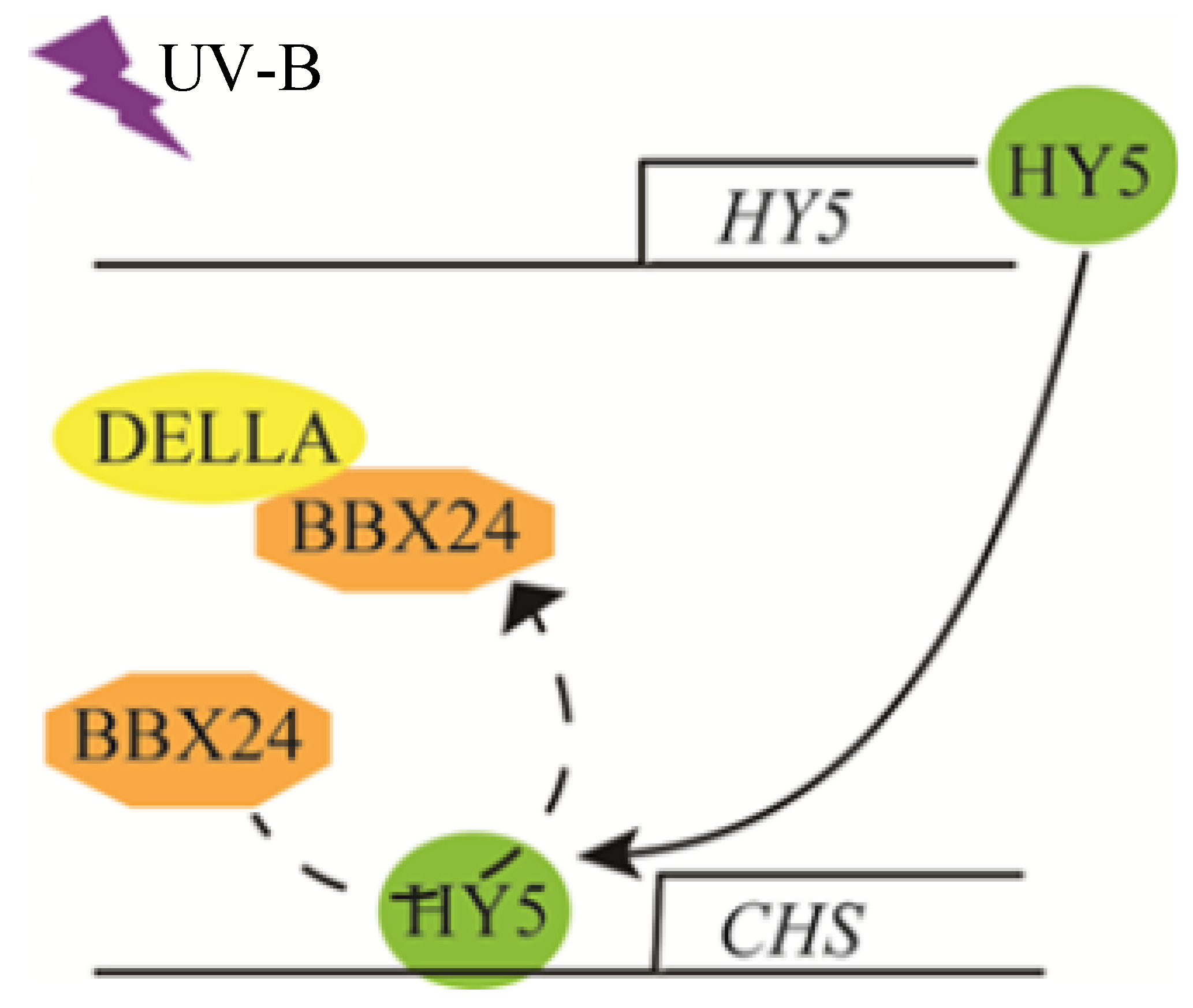
3. Discussion
4. Materials and Methods
4.1. Plant Materials and UV-B Treatment
4.2. Plant Hypocotyl Measurements
4.3. Anthocyanin and Flavonoid Measurement
4.4. RNA Extraction and Real-Time PCR
4.5. GA and PAC Treatments
4.6. Confocal Microscopy
4.7. Yeast Two-Hybrid Assay
4.8. Co-IP Assays and Immunoblot Analyses
4.9. Bi-Molecular Fluorescence Complementation
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fina, J.; Casadevall, R.; AbdElgawad, H.; Prinsen, E.; Markakis, M.N.; Beemster, G.T.S.; Casati, P. UV-B inhibits leaf growth through cchanges in growth regulating factors and gibberellin levels. Plant Physiol. 2017, 174, 1110–1126. [Google Scholar] [CrossRef] [Green Version]
- Henry-Kirk, R.A.; Plunkett, B.; Hall, M.; McGhie, T.; Allan, A.C.; Wargent, J.J.; Espley, R.V. Solar UV light regulates flavonoid metabolism in apple (Malus x domestica). Plant Cell Environ. 2018, 41, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, M.; Zhang, G.; Li, P.; Ma, F. Differential Regulation of Anthocyanin Synthesis in Apple Peel under Different Sunlight Intensities. Int. J. Mol. Sci. 2019, 20, 6060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijde, M.; Ulm, R. Reversion of the Arabidopsis UV-B photoreceptor UVR8 to the homodimeric ground state. Proc. Natl. Acad. Sci. USA 2012, 110, 1113–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriconi, V.; Binkert, M.; Costigliolo, C.; Sellaro, R.; Ulm, R.; Casal, J.J. Perception of Sunflecks by the UV-B Photoreceptor UV RESISTANCE LOCUS8. Plant Physiol. 2018, 177, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Ulm, R. How plants cope with UV-B: From perception to response. Curr. Opin. Plant Biol. 2017, 37, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, R.; Arongaus, A.B.; Binkert, M.; Ulm, R. Two distinct domains of the UVR8 photoreceptor interact with COP1 to initiate UV-B signaling in Arabidopsis. Plant Cell 2015, 27, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, X.; Jang, Z.; Chen, Z.; Ruo, X.; Jin, W.; Wu, Y.; Shi, X.; Xu, M. UV RESISTANCE LOCUS 8 from Chrysanthemum morifolium Ramat (CmUVR8) plays important roles in UV-B signal transduction and UV-B-induced accumulation of flavonoids. Front. Plant Sci. 2018, 9, 955. [Google Scholar] [CrossRef]
- Ren, H.; Han, J.; Yang, P.; Mao, W.; Liu, X.; Qiu, L.; Qian, C.; Liu, Y.; Chen, Z.; Ouyang, X.; et al. Two E3 ligases antagonistically regulate the UV-B response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 4722–4731. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Skvortsova, M.Y.; Loubéry, S.; Ulm, R. COP1 is required for UV-B–induced nuclear accumulation of the UVR8 photoreceptor. Proc. Natl. Acad. Sci. USA 2016, 113, E4415–E4422. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Mei, S.; Shi, C.; Yang, Y.; Peng, Y.; Ma, L.; Wang, F.; Li, X.; Huang, X.; Yin, Y.; et al. UVR8 Interacts with BES1 and BIM1 to Regulate Transcription and Photomorphogenesis in Arabidopsis. Dev. Cell 2018, 44, 512–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhaelewyn, L.; Viczián, A.; Prinsen, E.; Bernula, P.; Serrano, A.M.; Arana, M.V.; Ballaré, C.L.; Nagy, F.; Van Der Straeten, D.; Vandenbussche, F. Differential UVR8 signal across the stem controls UV-B–induced inflorescence phototropism. Plant Cell 2019, 31, 2070–2088. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yu, R.; Fan, L.-M.; Wei, N.; Chen, H.; Deng, X.W. DELLA-mediated PIF degradation contributes to coordination of light and gibberellin signalling in Arabidopsis. Nat. Commun. 2016, 7, 11868. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Gao, Z.; Zhu, Z. DELLA–PIF modules: Old dogs learn new tricks. Trends Plant Sci. 2016, 21, 813–815. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.; Zhong, S. Interplay between light and plant hormones in the control of Arabidopsis seedling chlorophyll biosynthesis. Front. Plant Sci. 2017, 8, 1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolgikh, A.V.; Kirienko, A.N.; Tikhonovich, I.A.; Foo, E.; Dolgikh, E.A. The DELLA proteins influence the expression of cytokinin biosynthesis and response genes during nodulation. Front. Plant Sci. 2019, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Tourinan, N.; Legris, M.; Minguet, E.G.; Costigliolo-Rojas, C.; Nohales, M.A.; Iniesto, E.; Garcia-Leon, M.; Pacin, M.; Heucken, N.; Blomeier, T.; et al. COP1 destabilizes DELLA proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 13792–13799. [Google Scholar] [CrossRef]
- Davies, J.-M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar]
- Wang, Y.; Deng, D. Molecular basis and evolutionary pattern of GA–GID1–DELLA regulatory module. Mol. Genet. Genom. 2013, 289, 1–9. [Google Scholar] [CrossRef]
- Zhao, B.; Li, H.; Li, J.; Wang, B.; Dai, C.; Wang, J.; Liu, K. Brassica napus DS-3, encoding a DELLA protein, negatively regulates stem elongation through gibberellin signaling pathway. Theor. Appl. Genet. 2017, 130, 727–741. [Google Scholar] [CrossRef]
- Li, W.; Zhang, J.; Sun, H.; Wang, S.; Chen, K.; Liu, Y.; Li, H.; Ma, Y.; Zhang, Z. FveRGA1, encoding a DELLA protein, negatively regulates runner production in Fragaria vesca. Planta 2017, 247, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Alabadi, D.; Gil, J.; Blazquez, M.A.; Garcia-Martinez, J.L. Gibberellins repress photomorphogenesis in darkness. Plant Physiol. 2004, 134, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, S.; Velanis, C.N.; Jenkins, G.I.; Franklin, K.A. UV-B detected by the UVR8 photoreceptor antagonizes auxin signaling and plant shade avoidance. Proc. Natl. Acad. Sci. USA 2014, 111, 11894–11899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crocco, C.D.; Locascio, A.; Escudero, C.M.; Alabadí, D.; Blázquez, M.A.; Botto, J.F. The transcriptional regulator BBX24 impairs DELLA activity to promote shade avoidance in Arabidopsis thaliana. Nat. Commun. 2015, 6, 6202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Wang, Y.; Li, Q.-F.; Björn, L.O.; He, J.-X.; Li, S.-S. Arabidopsis STO/BBX24 negatively regulates UV-B signaling by interacting with COP1 and repressing HY5 transcriptional activity. Cell Res. 2012, 22, 1046–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef]
- Xu, F.; Li, T.; Xu, P.-B.; Li, L.; Du, S.-S.; Lian, H.-L.; Yang, H.-Q. DELLA proteins physically interact with CONSTANS to regulate flowering under long days in Arabidopsis (U-I Flügge, Ed.). FEBS Lett. 2016, 590, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Stawska, M.; Oracz, K. phyB and HY5 are involved in the blue light-mediated alleviation of dormancy of Arabidopsis seeds possibly via the modulation of expression of genes related to light, GA, and ABA. Int. J. Mol. Sci. 2019, 20, 5882. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zheng, S.; Liu, Z.; Wang, L.; Bi, Y. Both HY5 and HYH are necessary regulators for low temperature-induced anthocyanin accumulation in Arabidopsis seedlings. J. Plant Physiol. 2011, 168, 367–374. [Google Scholar] [CrossRef]
- Miao, T.; Li, D.; Huang, Z.; Huang, Y.; Li, S.; Wang, Y. Gibberellin regulates UV-B-induced hypocotyl growth inhibition in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, e1966587. [Google Scholar] [CrossRef]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorrai, R.; Boccaccini, A.; Ruta, V.; Possenti, M.; Costantino, P.; Paola, V. ABA inhibits hypocotyl elongation acting on gibberellins, DELLA proteins and auxin. AoB Plants 2018, 10, ply061. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Gong, Y.; Liu, B.; Wu, D.; Zhang, M.; Xie, D.; Song, S. The DELLA proteins interact with MYB21 and MYB24 to regulate filament elongation in Arabidopsis. BMC Plant Biol. 2020, 20, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Z.; Liu, J.; Lin, S.; Wang, J.; Lin, W.; Xu, W. GA-DELLA pathway is involved in regulation of nitrogen deficiency-induced anthocyanin accumulation. Plant Cell Rep. 2017, 36, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Clayton, W.A.; Albert, N.W.; Thrimawithana, A.H.; McGhie, T.K.; Deroles, S.C.; Schwinn, K.E.; Warren, B.A.; McLachlan, A.R.; Bowman, J.L.; Jordan, B.R.; et al. UVR8-mediated induction of flavonoid biosynthesis for UVB tolerance is conserved between the liverwort Marchantia polymorph and flowering plants. Plant J. 2018, 96, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Man, C.; Xie, Y.; Yan, J.; Chu, J.; Huang, J. A crucial role of GA-regulated flavonol biosynthesis in root growth of Arabidopsis. Mol. Plant 2019, 12, 521–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, G.; Li, D.; Xiong, H.; Xiao, L.; Tong, J.; Ning, C.; Wang, P.; Li, S. Quantitative Proteomic Analyses Identify STO/BBX24 -Related Proteins Induced by UV-B. Int. J. Mol. Sci. 2020, 21, 2496. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Ramos, L.A.; O’Hara, A.; Kanagarajan, S.; Farkas, D.; Strid, Å.; Jenkins, G.I. Difference in the action spectra for UVR8 monomerization and HY5 transcript accumulation in Arabidopsis. Photochem. Photobiol. Sci. 2018, 17, 1108–1117. [Google Scholar] [CrossRef] [Green Version]
- Verdaguer, D.; Jansen, M.A.K.; Llorens, L.; Morales, L.O.; Neugart, S. UV-A radiation effects on higher plants: Exploring the known unknown. Plant Sci. 2017, 255, 72–81. [Google Scholar] [CrossRef]
- Kucera, B.; Leubner-Metzger, G.; Wellmann, E. Distinct Ultraviolet-Signaling Pathways in Bean Leaves. DNA damage is associated with β-1,3-glucanase gene induction, but not with flavonoid formation. Plant Physiol. 2003, 133, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Gangappa, S.N.; Prasad, V.B.R.; Chattopadhyay, S. Functional interconnection of MYC2 and SPA1 in the photomorphogenic seedling development of Arabidopsis. Plant Physiol. 2010, 154, 1210–1219. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Xiong, H.; Xie, Y.; Lyu, S.; Miao, T.; Li, T.; Lyu, G.; Li, S. BBX24 Interacts with DELLA to Regulate UV-B-Induced Photomorphogenesis in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 7386. https://doi.org/10.3390/ijms23137386
Huang Y, Xiong H, Xie Y, Lyu S, Miao T, Li T, Lyu G, Li S. BBX24 Interacts with DELLA to Regulate UV-B-Induced Photomorphogenesis in Arabidopsis thaliana. International Journal of Molecular Sciences. 2022; 23(13):7386. https://doi.org/10.3390/ijms23137386
Chicago/Turabian StyleHuang, Yuewei, Hui Xiong, Yuxin Xie, Suihua Lyu, Tingting Miao, Tingting Li, Guizhen Lyu, and Shaoshan Li. 2022. "BBX24 Interacts with DELLA to Regulate UV-B-Induced Photomorphogenesis in Arabidopsis thaliana" International Journal of Molecular Sciences 23, no. 13: 7386. https://doi.org/10.3390/ijms23137386
APA StyleHuang, Y., Xiong, H., Xie, Y., Lyu, S., Miao, T., Li, T., Lyu, G., & Li, S. (2022). BBX24 Interacts with DELLA to Regulate UV-B-Induced Photomorphogenesis in Arabidopsis thaliana. International Journal of Molecular Sciences, 23(13), 7386. https://doi.org/10.3390/ijms23137386