1. Introduction
Metal ions are inherent elements that enable the course of physiological processes in living organisms. Besides macronutrients such as Ca
2+, Mg
2+, or K
+ ions, plants also require micronutrients for proper development, including heavy metals. Some heavy metal ions play important roles in biochemical processes in plant cells, such as Cu
2+ ions, which are involved in electron transport reactions in photosynthesis and respiration [
1]. However, most heavy metals are nonessential for plants and may negatively impact their growth and development, even in small quantities. Among the various heavy metals, Cd is considered the most phytotoxic one. Cadmium salts are soluble in water, and Cd ions are easily absorbed by plants, thus entering the food chain [
2,
3]. Cadmium toxicity is represented by the replacement of biogenic metals in the active centers of enzymes and the high affinity to the thiol groups of proteins, which inhibits enzyme activity [
4,
5]. Cadmium toxicity in plants is manifested by stunted growth and chlorosis; at higher concentrations, it can lead to growth inhibition and necrosis [
6,
7] as well as reduced photosynthetic activity by decreasing the chlorophyll content [
8]. As soil is the main sink for heavy metals, plant roots are the most important organs in metal ion uptake and relocation. Roots are also the first organs directly affected by cadmium damage, disturbing the normal physiological metabolism of the entire plant [
7,
9].
A critical step in plant survival strategy during exposure to external stress is maintaining the fluidity and integrity of the cell membranes [
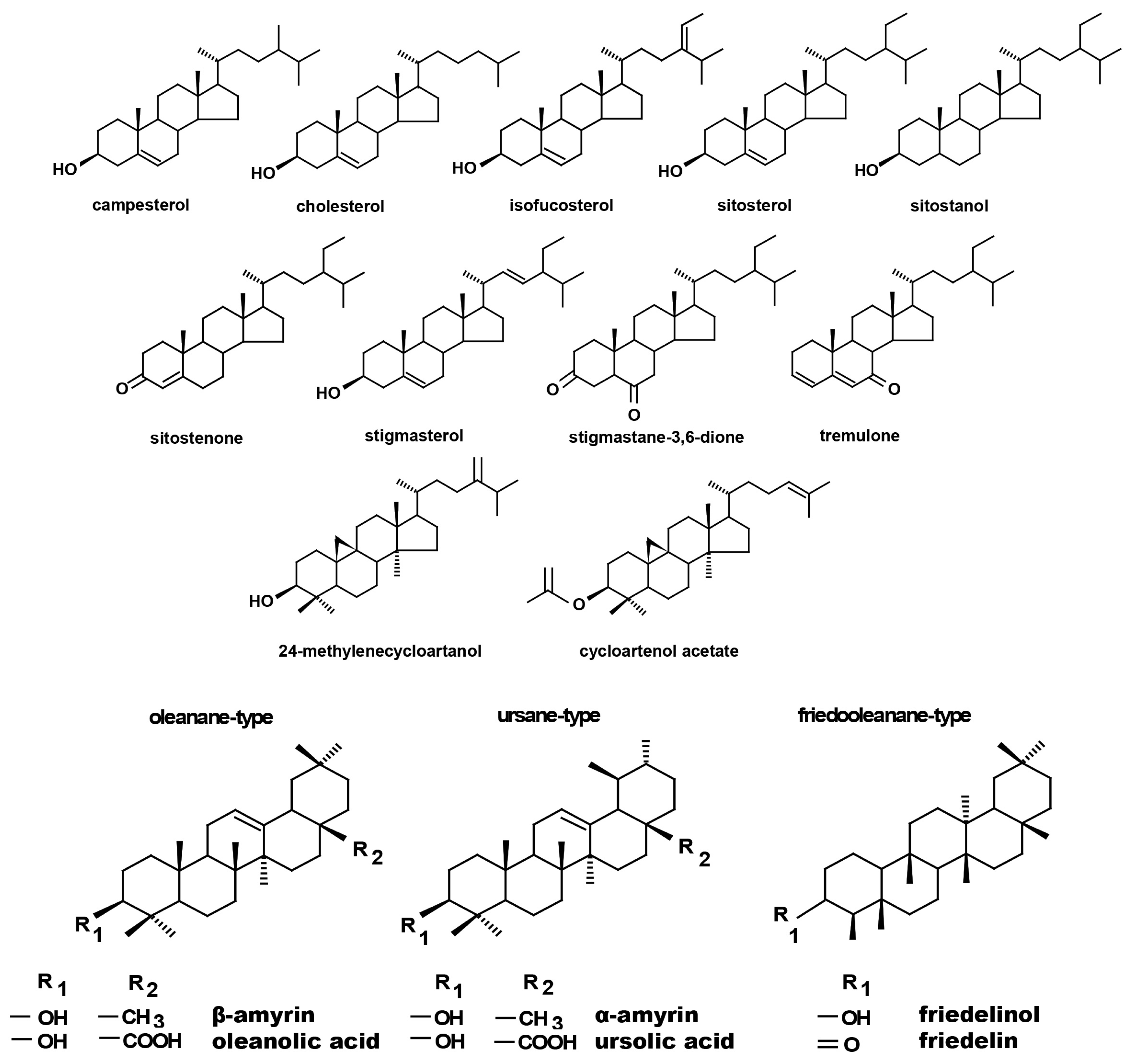
10]. As sterols are plant membrane components, changes in the profiles of these compounds seems to be important as a response to adverse environmental conditions. Sterols not only perform a structural function along with sphingolipids and glycerolipids, but also influence the activity of integral membrane proteins, including enzymes, ion channels, receptors, and components of signal transduction pathways (i.e., ATPases). These isoprenoids are regarded as primary or, more aptly, general metabolites due to their metabolic, regulatory, and membrane architectural functions [
11]. Cell membranes are important sites perceiving abiotic stress signals, and, therefore, the recomposition of endogenous sterols in these structures improves plant stress resistance. A specific role in this process is played by C24-alkyl sterols, which enhance plant resistance to stress factors by improving the cohesion of cell membranes and changing the permeability of membranes, thereby affecting proton efflux [
12,
13]. The proportions between sitosterol and stigmasterol (which contains 22(23)-double bond in the side chain) and the ratios of certain conjugated sterol forms (i.e., sterol esters and sterol glycosides) can change under abiotic stress [
11]. In sterol esters forms, fatty acids estrified to C3 hydroxyl group may vary in lengths from twelve carbon atoms to twenty-two. The most common fatty acids found in sterol esters are palmitic, stearic, oleic, linoleic, and linolenic. In the case of glycosylated sterols, sugar moiety is linked through β-glycosidic bond to C3 hydroxyl group of sterol core. The most common monosaccharide found in plant glycosylated sterols is glucose; however, other monosaccharides, such as galactose, xylose, and mannose, have also been identified [
14].
Another interesting aspect of plant response to stress is the relationship between general and secondary metabolism. Specialized metabolites deal with various external agents (including stress factors), protection against herbivores, or attraction of pollinators. Some stress conditions enhance the expression of genes involved in the biosynthesis of certain natural defense compounds [
15]. Sterols, considered general metabolites, and triterpenoids, which are specialized compounds, have a common precursor, namely, squalene. Therefore, the metabolic strategy of the plant facing stress factors can be regarded as a kind of compromise: either the boost of the biosynthesis of specialized defense metabolites at the expense of general metabolites, which may result in growth inhibition, or enhanced biosynthesis of general metabolites to better adapt to adverse conditions, along with the reduced production of specialized metabolites.
Cadmium uptake by roots has been extensively studied in various important crop species, including cereals (maize, rice, and wheat) and vegetables (lettuce, radish, and onion). These studies provide evidence that cadmium ions directly affect the physical properties and biological functions of cell membranes in roots, along with an indirect effect via reactive oxygen species formation [
16]. However, little is known about the effect of cadmium stress on cell membrane composition in the context of possible alterations in sterol content. To date, only limited data are available on the influence of cadmium ions on sterols and secondary metabolites in
Calendula officinalis L. (calendula, marigold), a plant with ornamental and medicinal value and native to the south of Europe. Due to its high adaptability, it does not require specific soil conditions and seems to be a promising candidate for cadmium phytoremediation [
17,
18,
19].
The present study investigated the changes in the contents of steroids and triterpenoids in C. officinalis hairy root cultures and plants exposed to cadmium stress over time. Apart from the evaluation of the influence of cadmium ions on steroid and triterpenoid biosynthesis (and, thus, the potential trade-off between general and specialized metabolism), the additional aim of this study was to compare the observed effects in two different experimental models. Investigations of in vitro cultures are convenient due to the controlled environmental conditions as well as the less complex culture maintenance and supplementation with stress agents or elicitors. However, the common critical reproach against such studies is that in vitro cultures might not reflect the stress response of the plant under natural conditions. In this context, this study complements the knowledge on the usability of plant in vitro cultures as experimental models for research on physiological stress reactions, and not only for typical studies on plant productivity of valuable bioactive phytochemicals.
3. Discussion
This study investigated the influence of Cd stress on the steroid and triterpenoid metabolism in
C. officinalis plants and hairy roots cultures and represents a targeted GC-MS metabolomic approach to study stress-induced metabolic modifications. As it has been frequently reported for toxic metals and metalloids, high Cd concentrations in plants can alter metabolic processes and exert various effects at the physiological, morphological, and molecular levels, e.g., growth inhibition, nutrition imbalance, photosynthesis suppression, chlorosis, and ROS (reactive oxygen species) increase [
21,
22]. Various reports indicate that the composition of steroids and triterpenoids might be modified under stress conditions; however, detailed studies on such effects exerted after exposure to heavy metals are scarce [
11,
23,
24]. The first reported studies on this phenomenon, performed either on algae or plants, demonstrated effects such as increased levels of cholesterol and a decrease in the ratio of 24-ethylcholest-5-en-3β-ol to 24-ethylcholesta-5,24(28)
Z-dien-3β-ol, two major sterols occurring in the marine diatom
Asterionella gracilis [
25], or the increase of stigmasterol and sitosterol contents parallel to the decrease of isofucosterol and campesterol, as observed for rice (
Oryza sativa cv. Bahía) seedlings [
26,
27] exposed to Cd stress.
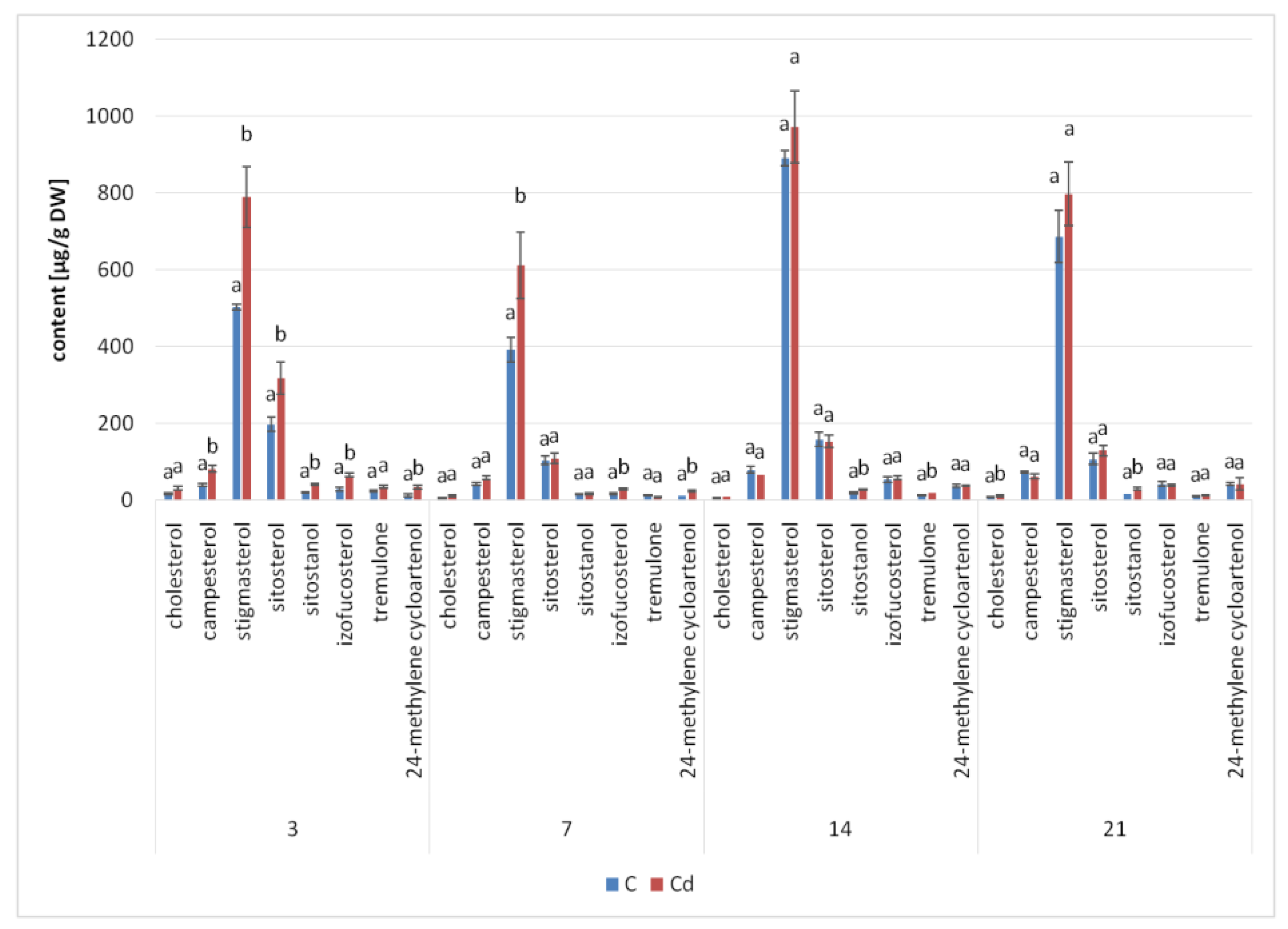
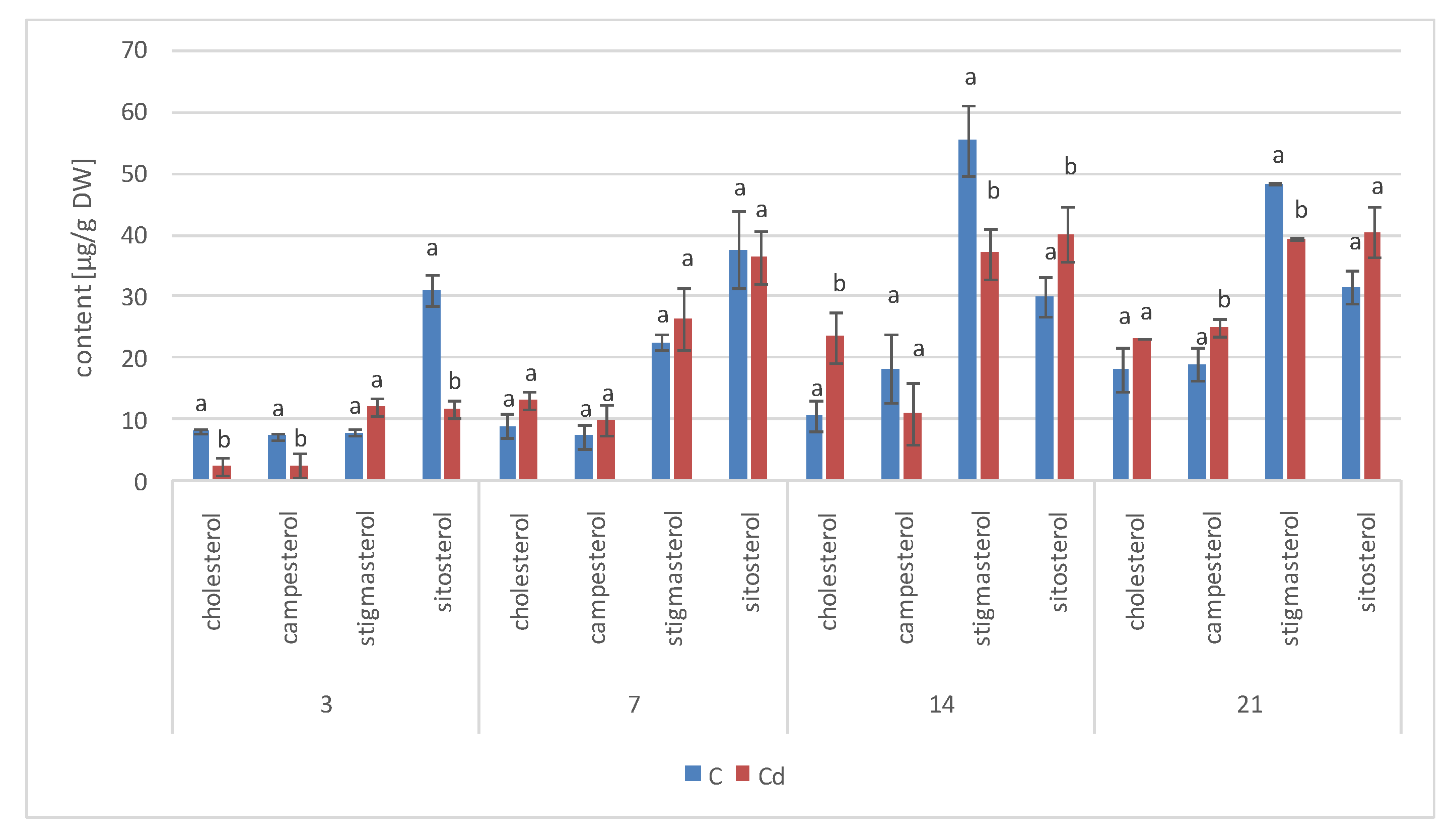
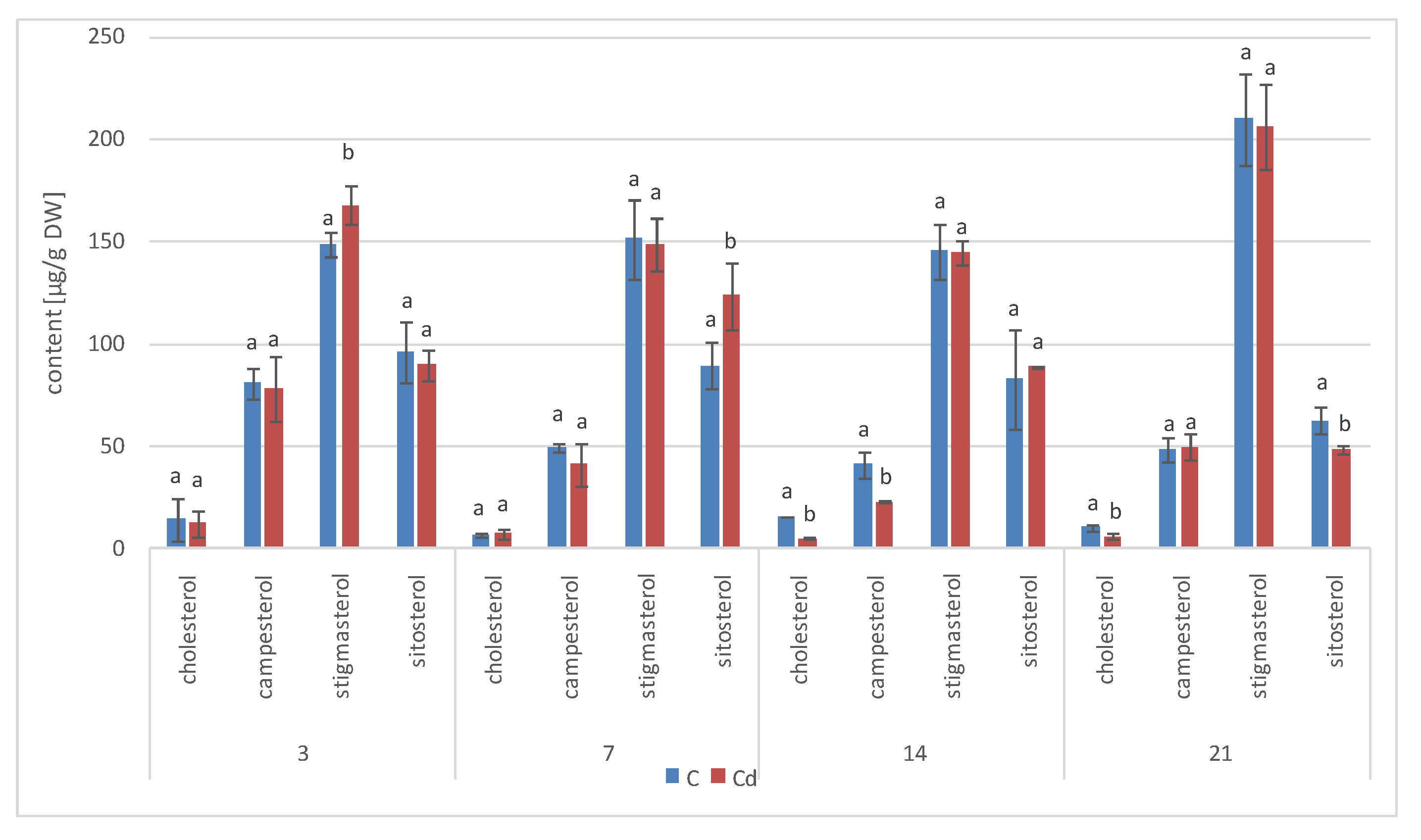
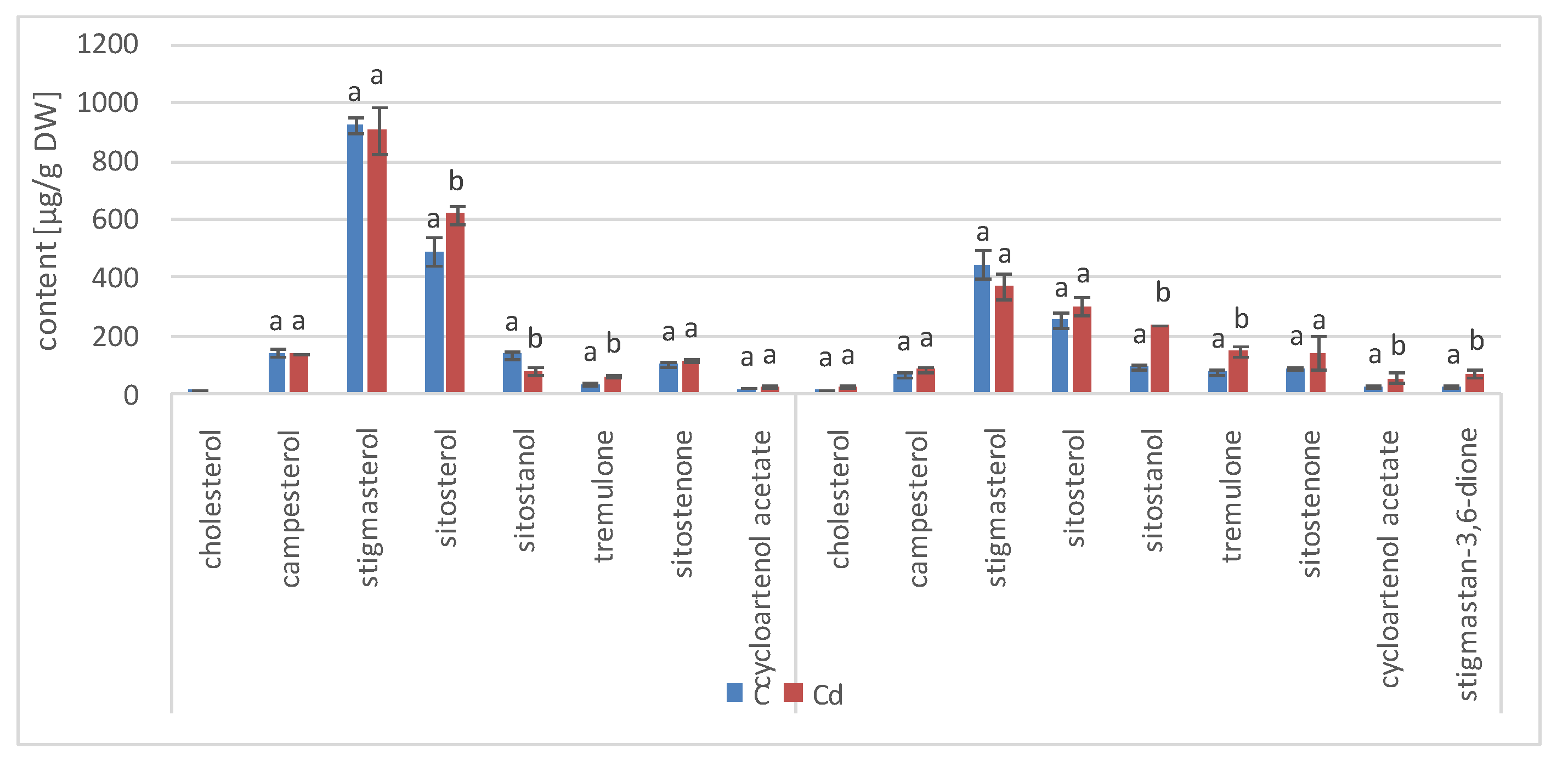
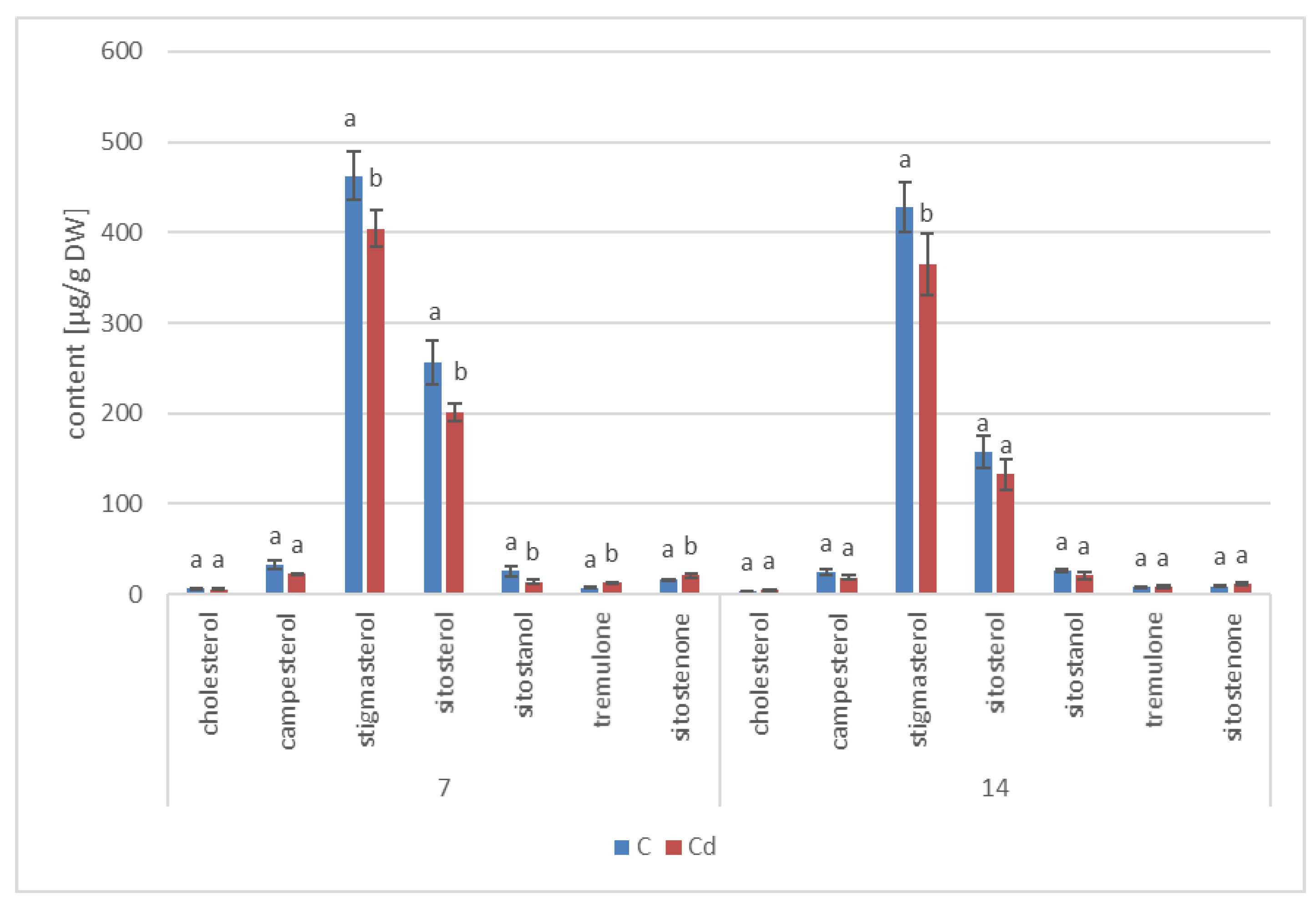
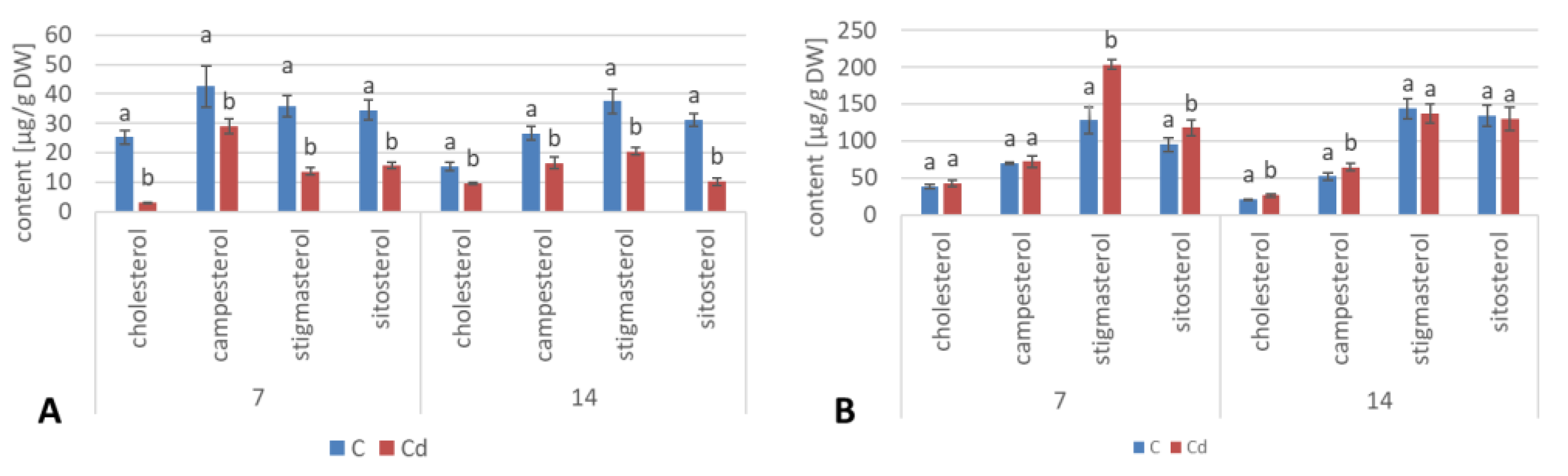
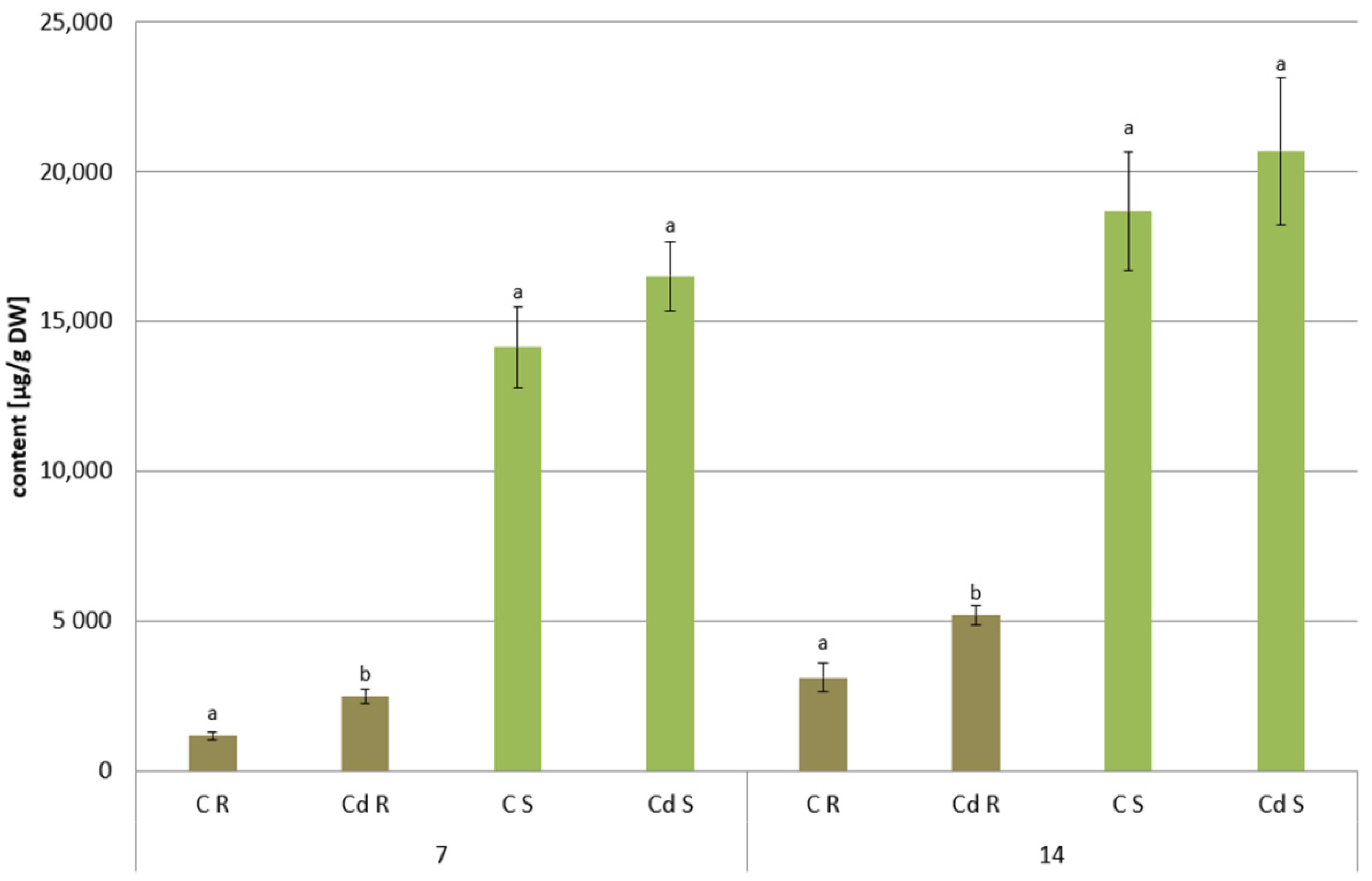
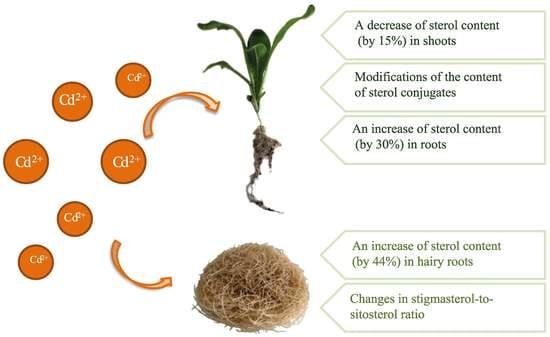
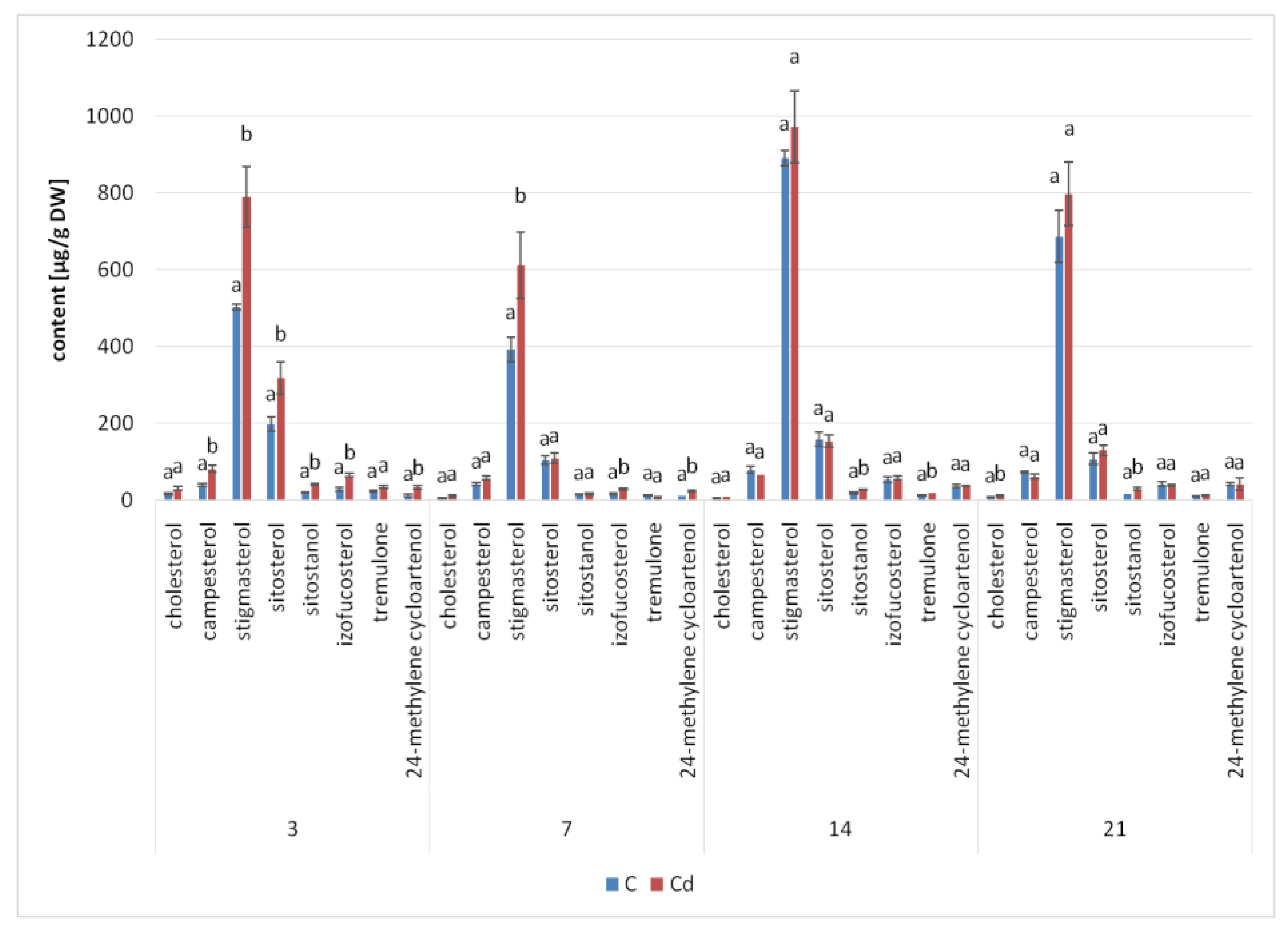
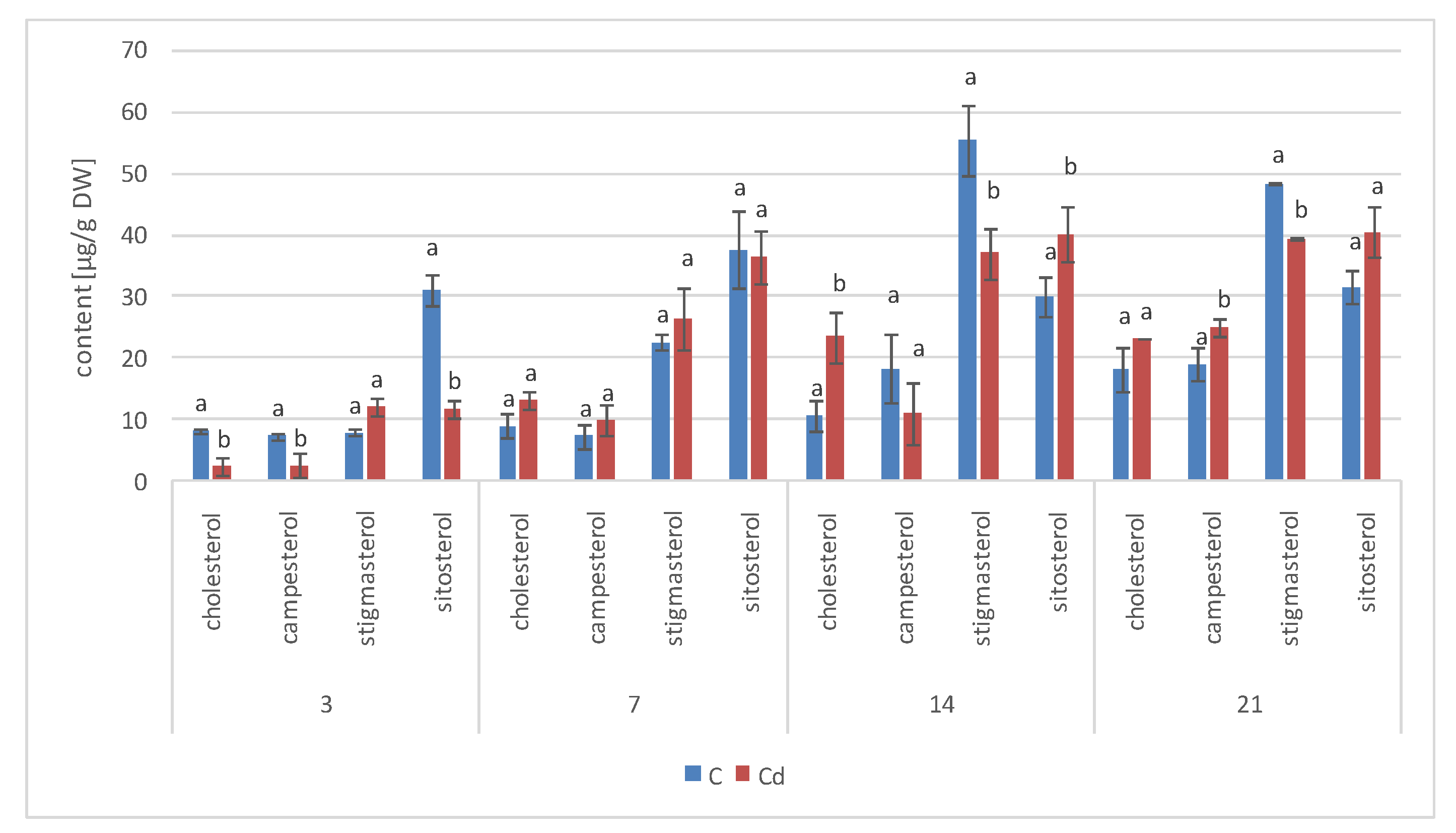
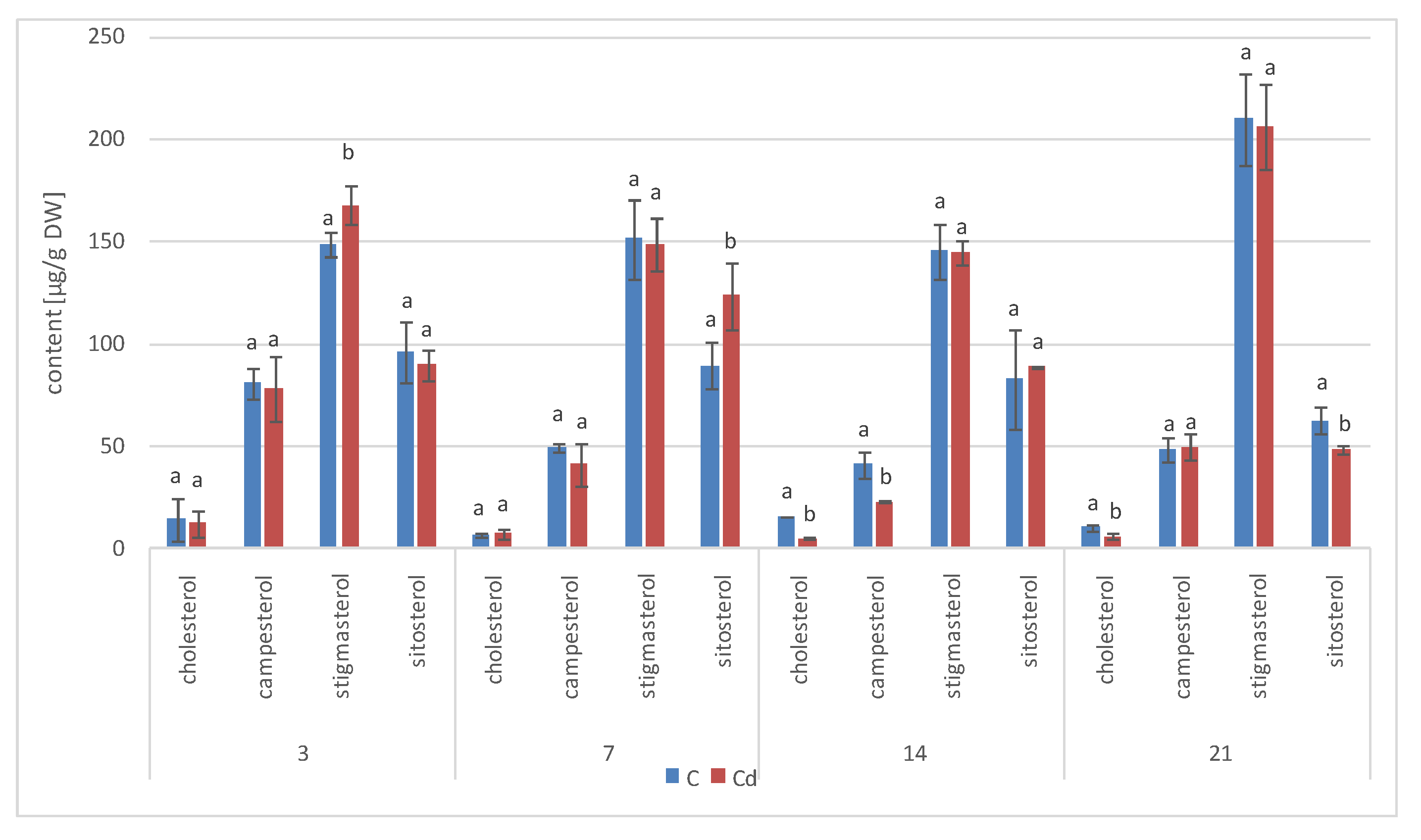
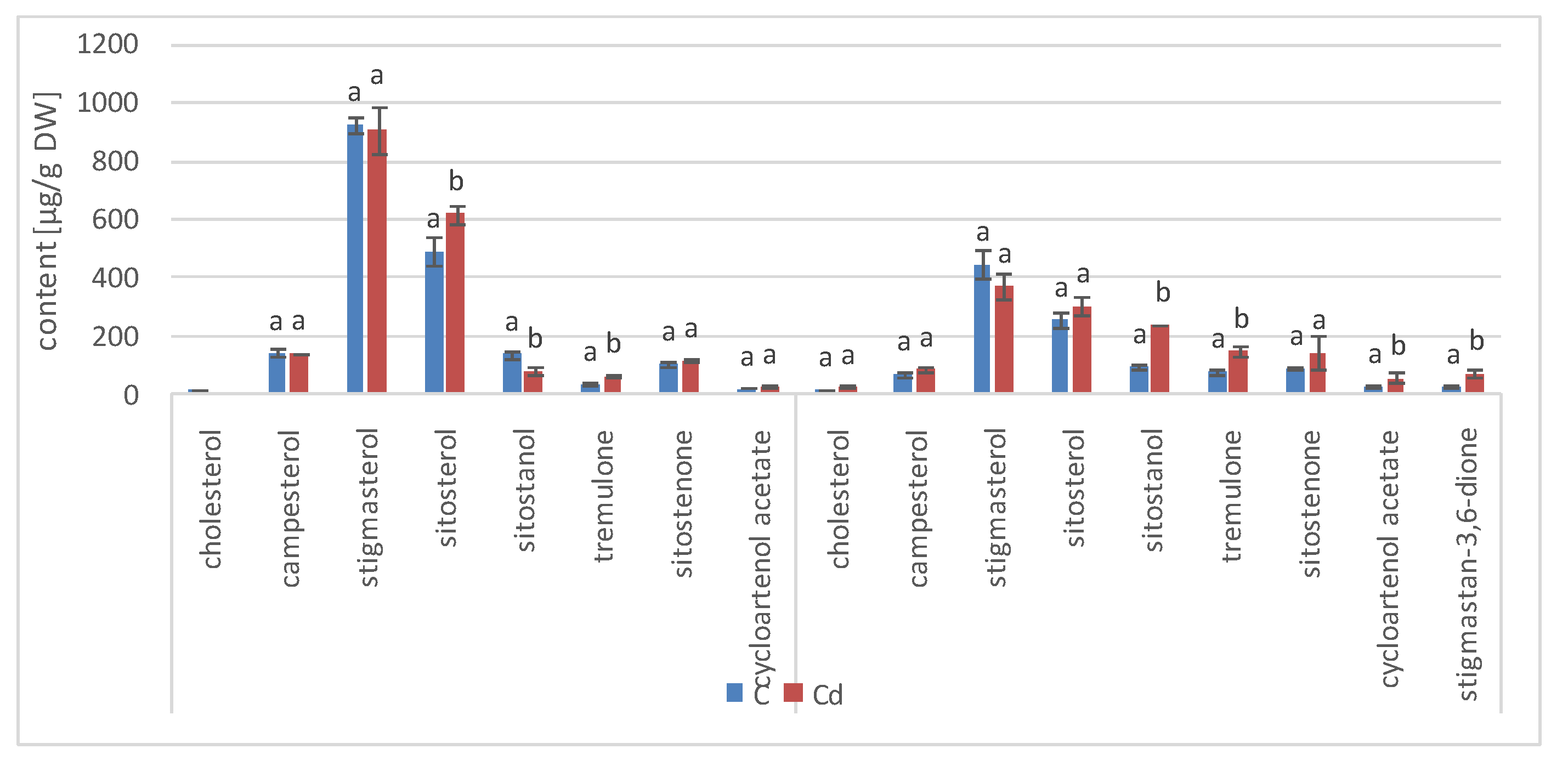
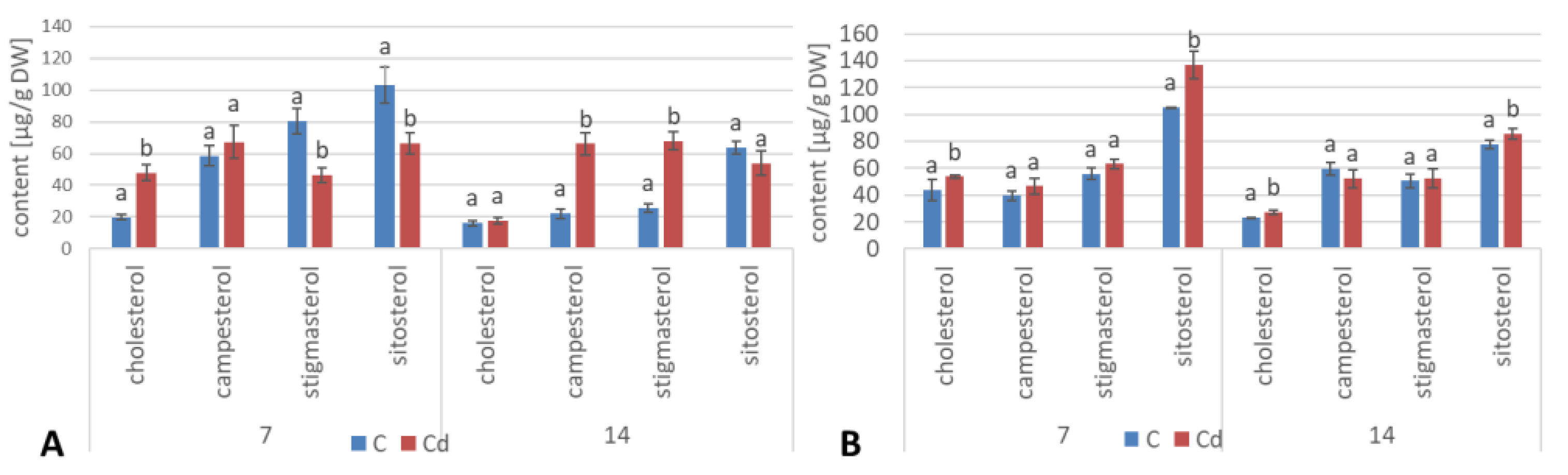
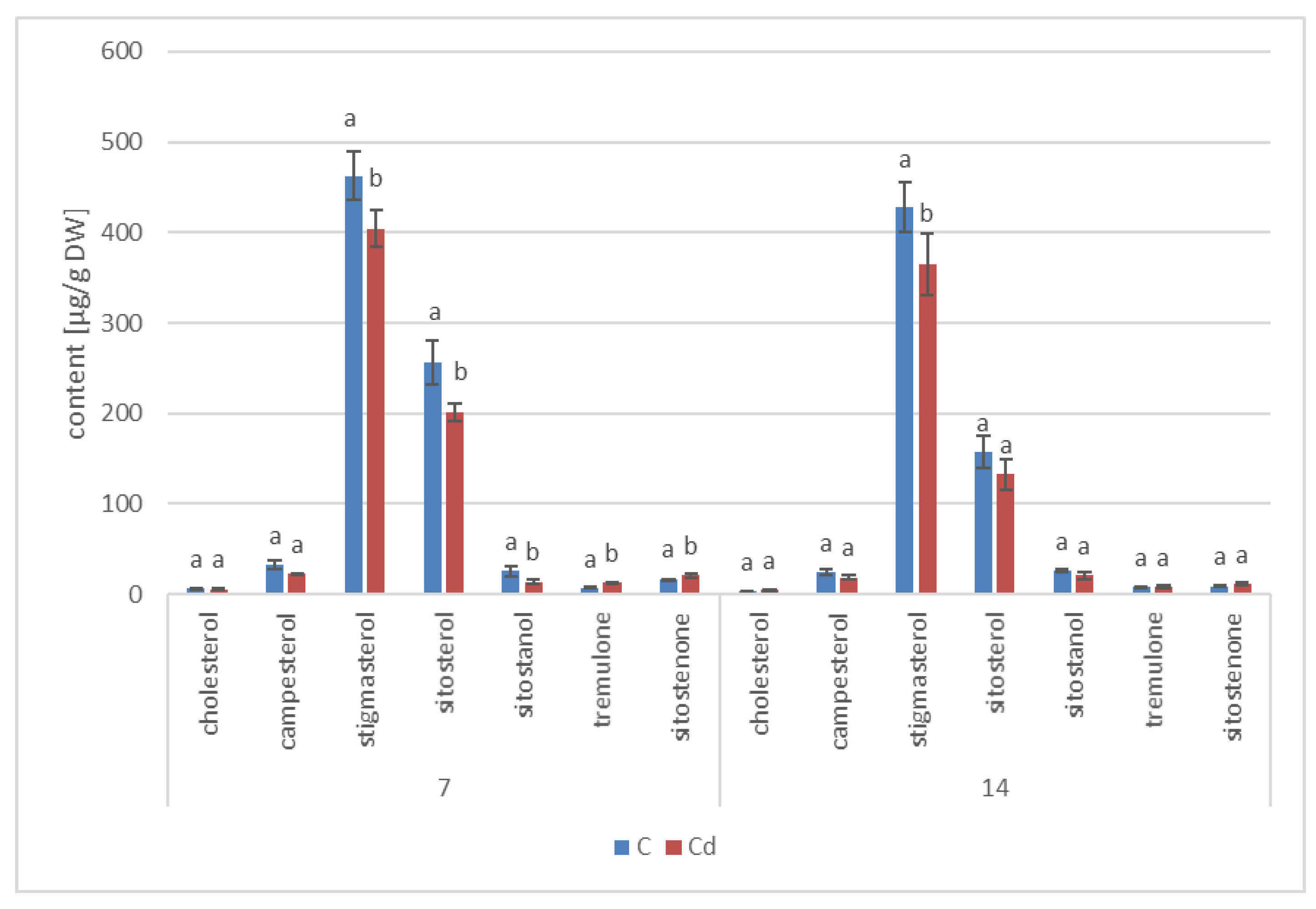
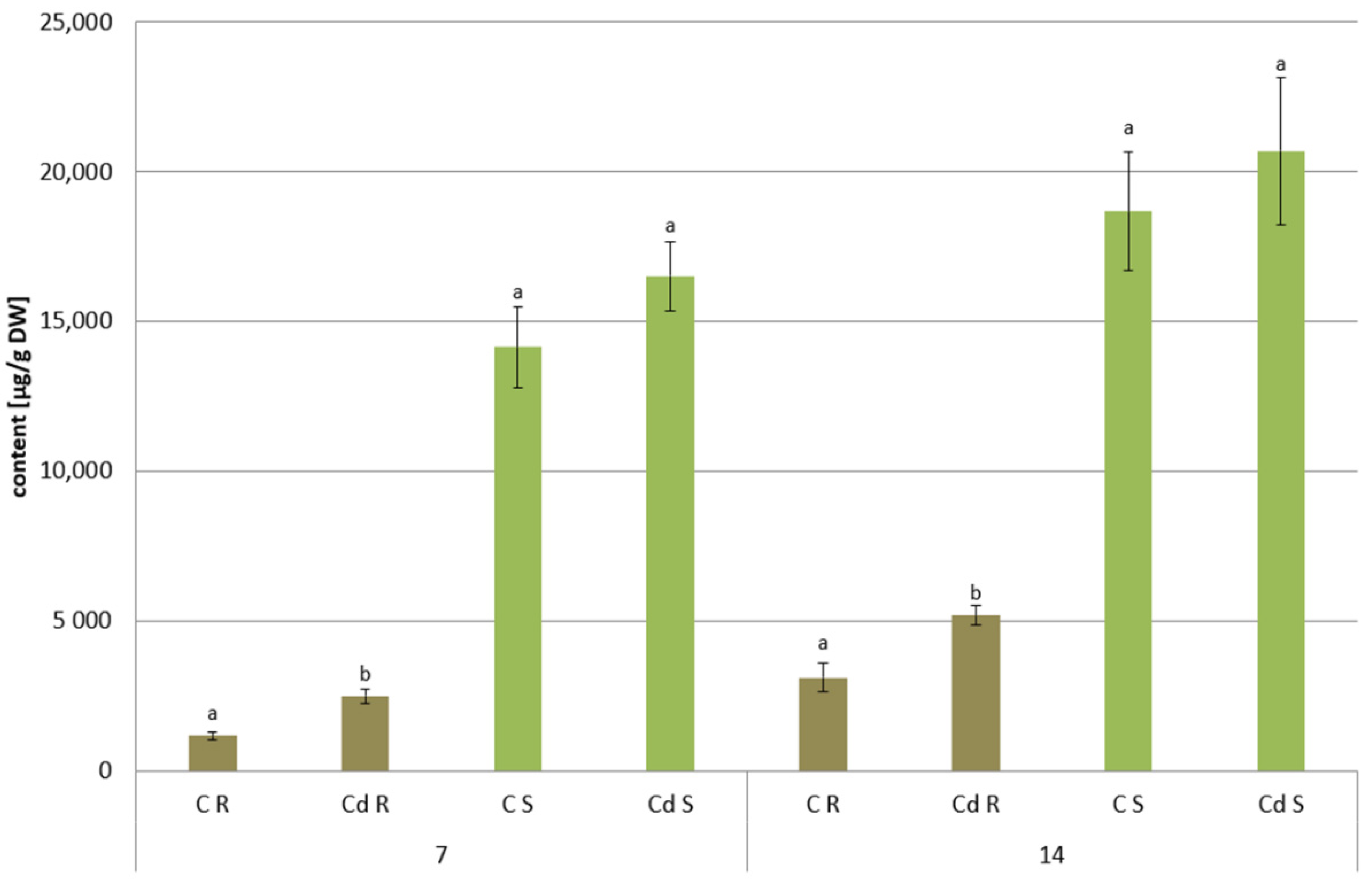
The results obtained in the present study clearly indicate that Cd stress significantly altered the contents of steroids, including sterols, as well as the proportions among individual constituents. However, it was observed that various organs of the same plant can react differently to Cd stress. The total sterol content increased in roots and hairy root culture and decreased in shoots; these effects were inversely correlated with Cd-induced growth suppression. A lack of a direct correlation between the changes in the growth and total sterol content has also been observed in previous studies, e.g., on
O. sativa plants exposed to Cd stress, where the reduced growth of both shoots and roots was accompanied with an approximately 30% increase in the sterol content. The growth suppress parallel to the stimulation of sterol biosynthesis can be explained by the restructuring of the membrane composition due to the accumulation of the excess sterols, leading to substantial changes in membrane properties, such as fluidity, permeability, and perception of stress signals; such effects have frequently been observed in plants and in particular in in-vitro cultures exposed to various stressors or elicitors [
20,
28]. Regarding the essential role of sterols as plasma membrane constituents, the explanation of the adverse phenomenon, i.e., the correlation between increased growth and a decrease in the sterol content, seems to be less obvious; however, the increase in biomass of plants and in-vitro cultures might not occur exclusively as a result of the increased number of cells. Indeed, in a previous study, the reported increase in the dry weight of mercury-treated algal cells appeared to be uncoupled from cell division [
25]. Moreover, the inhibition of sterol biosynthesis, according to the “plant Cornelian dilemma” of the growth-defense trade-off [
24], can significantly induce the biosynthesis of triterpenoids or other compounds excessively accumulating in cells under stress conditions
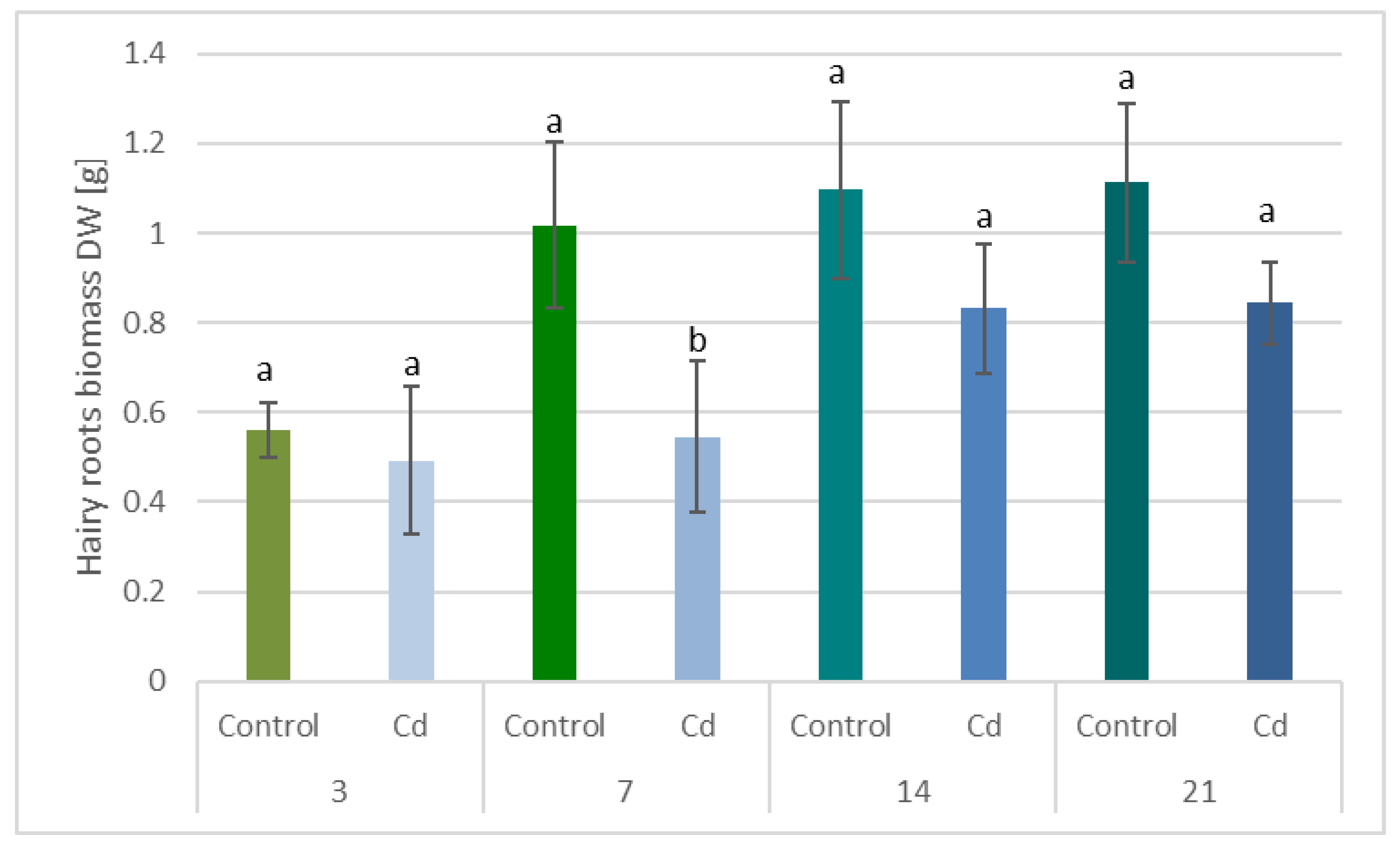
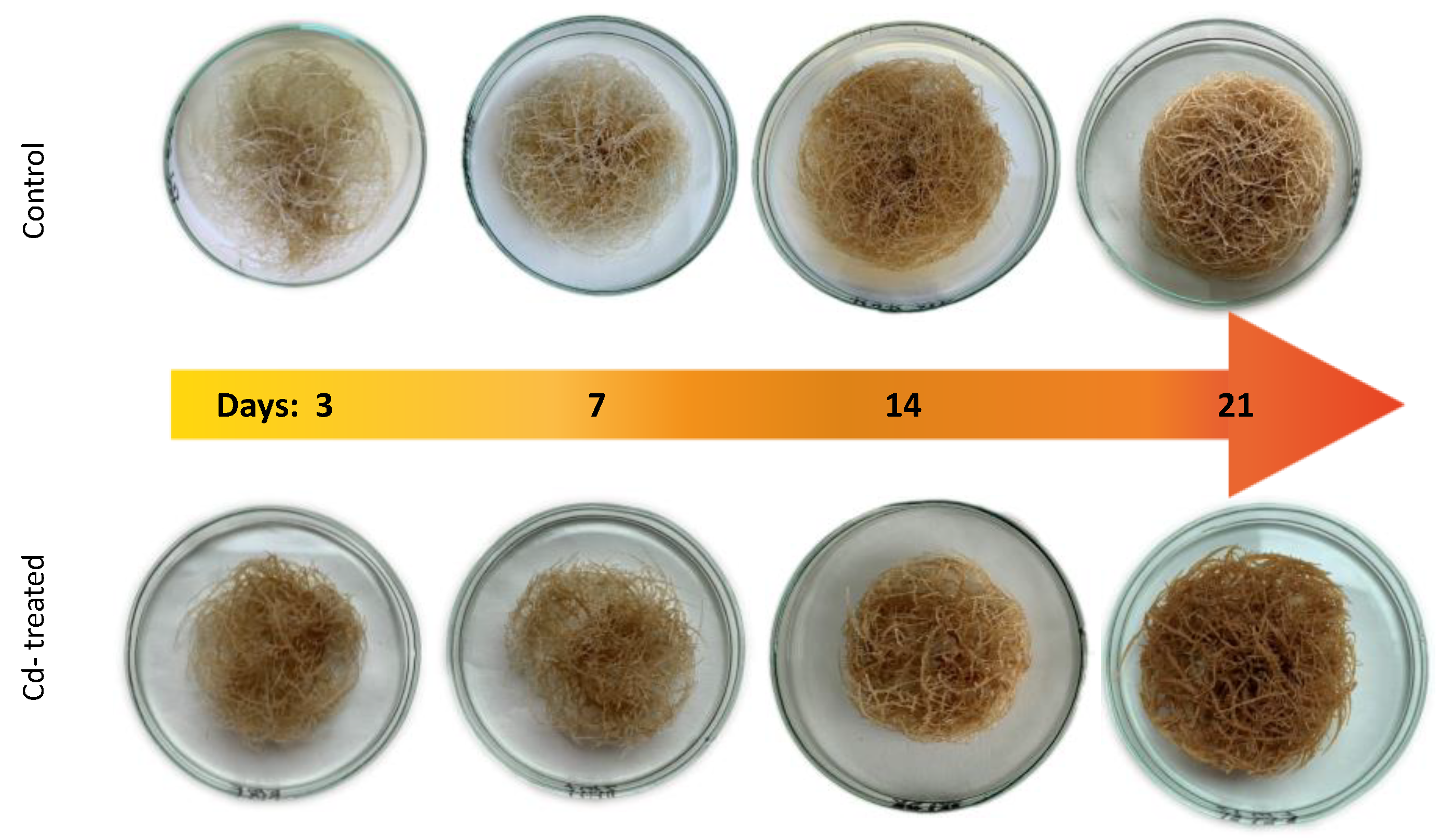
The observed decrease in both
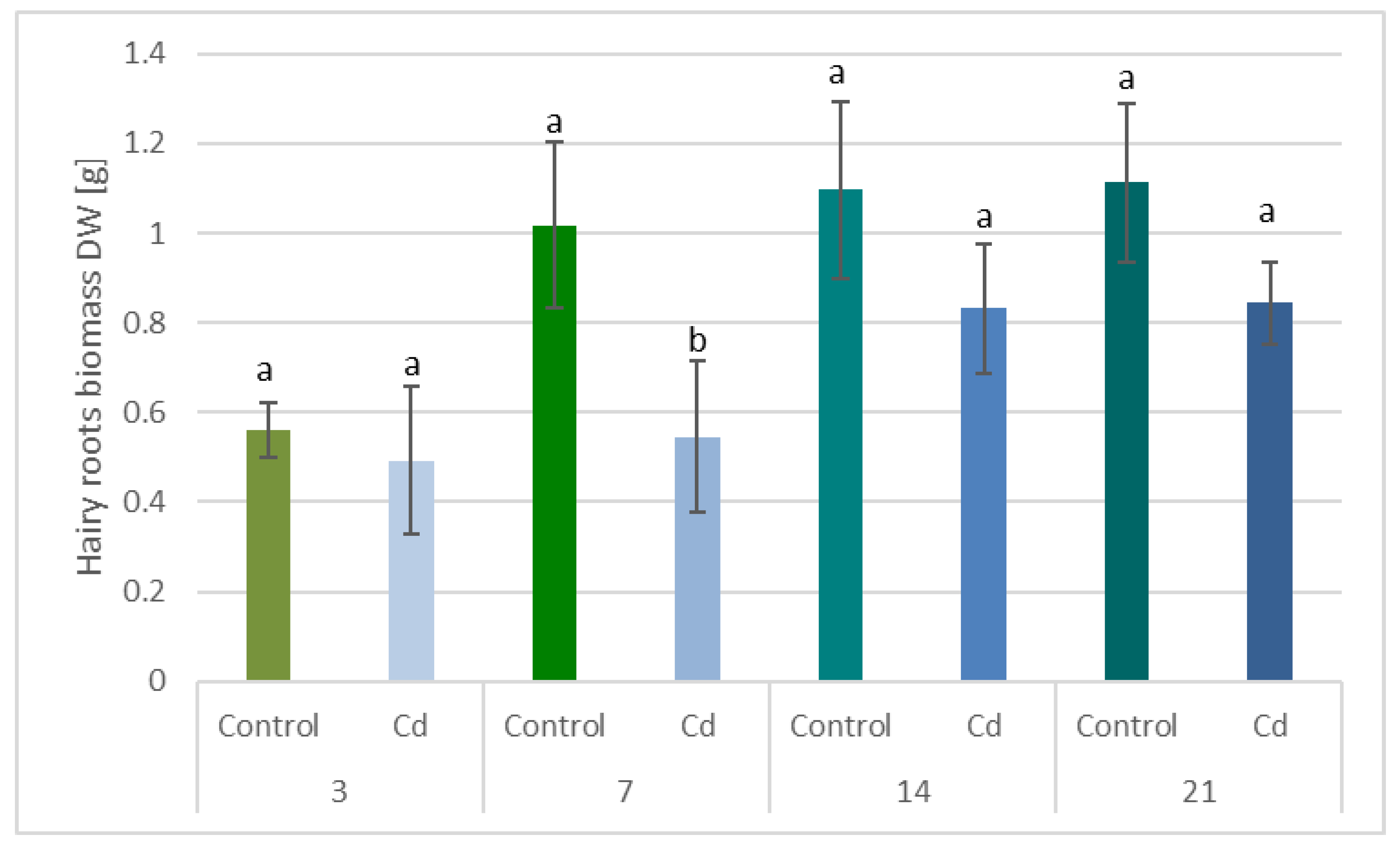
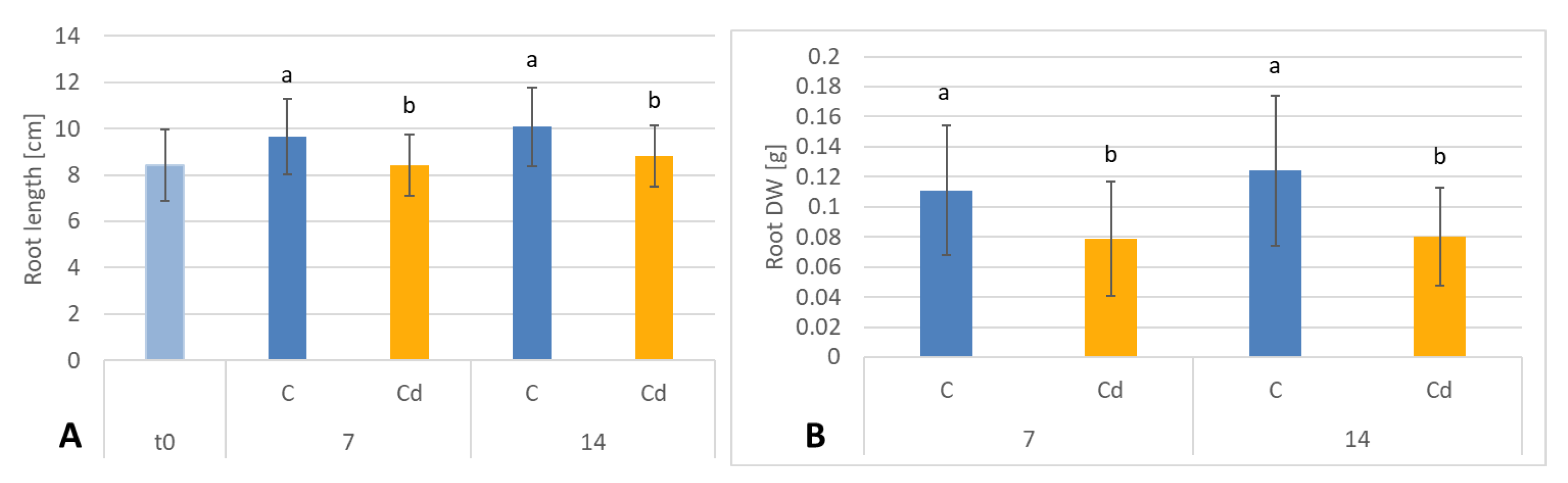
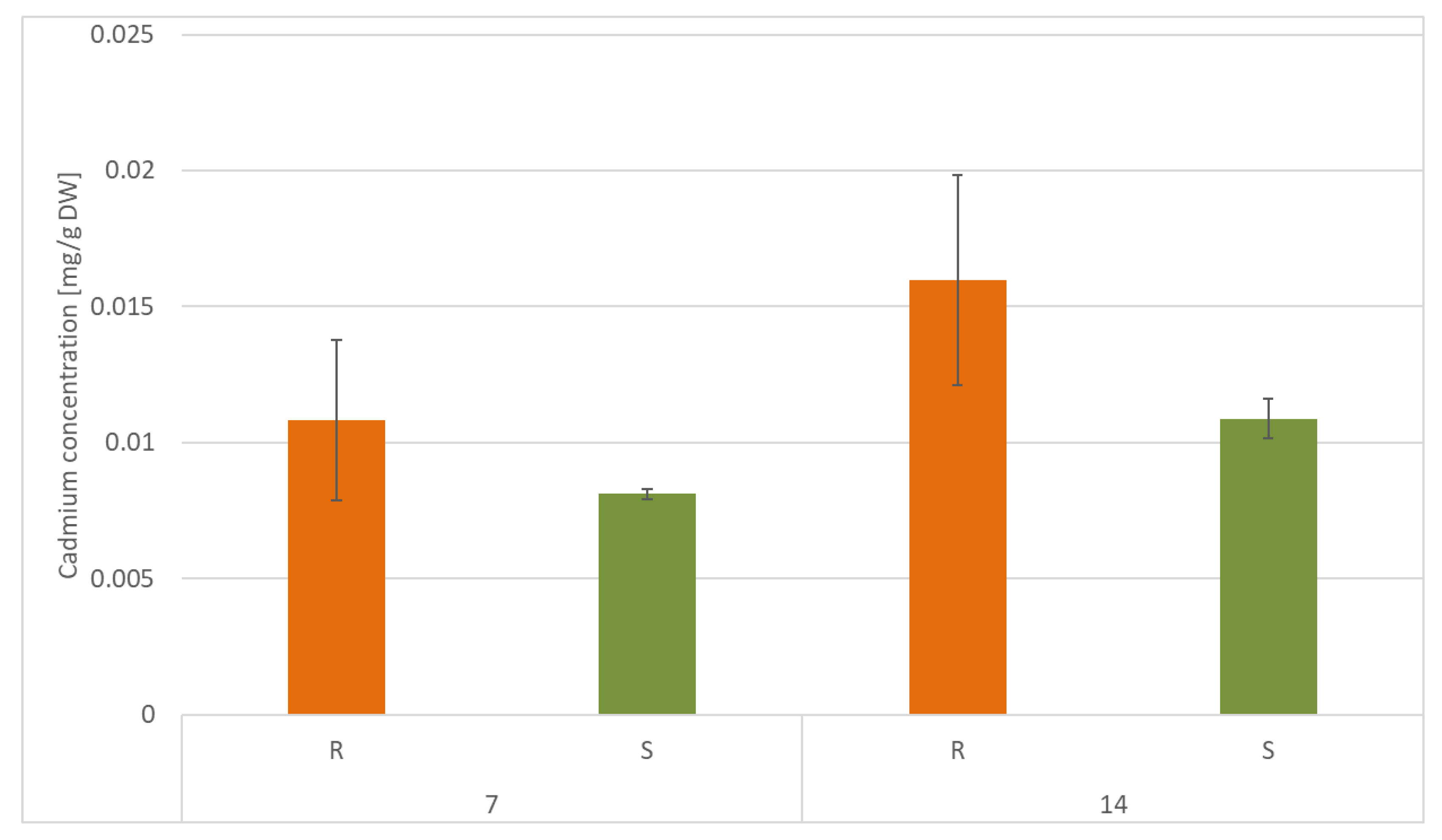
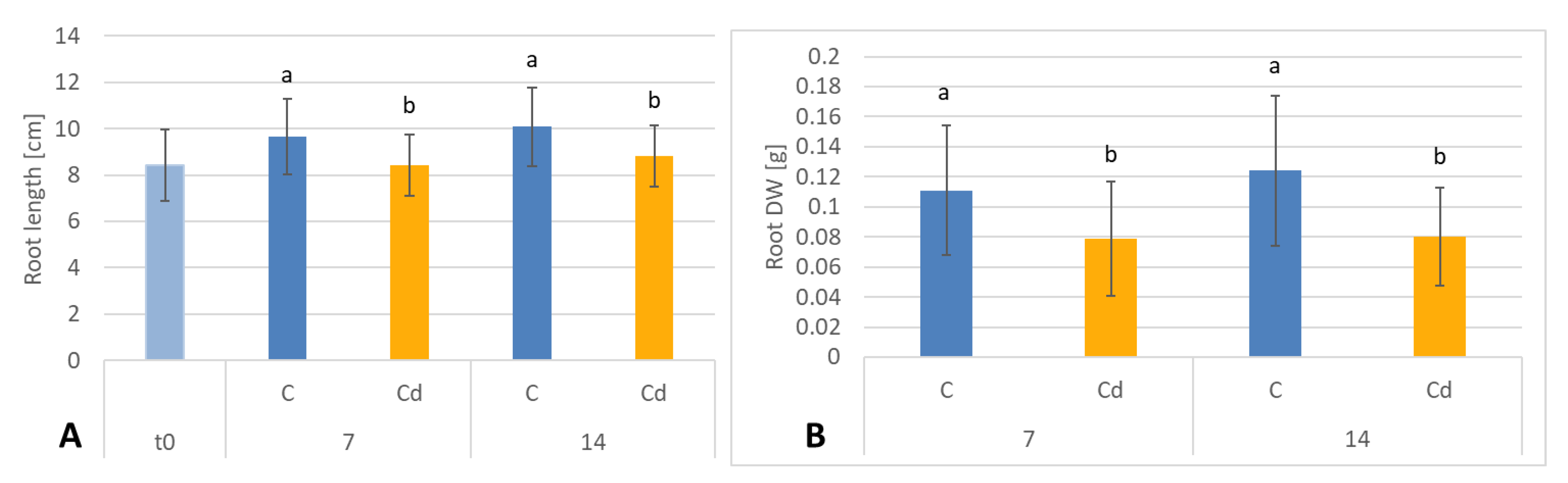
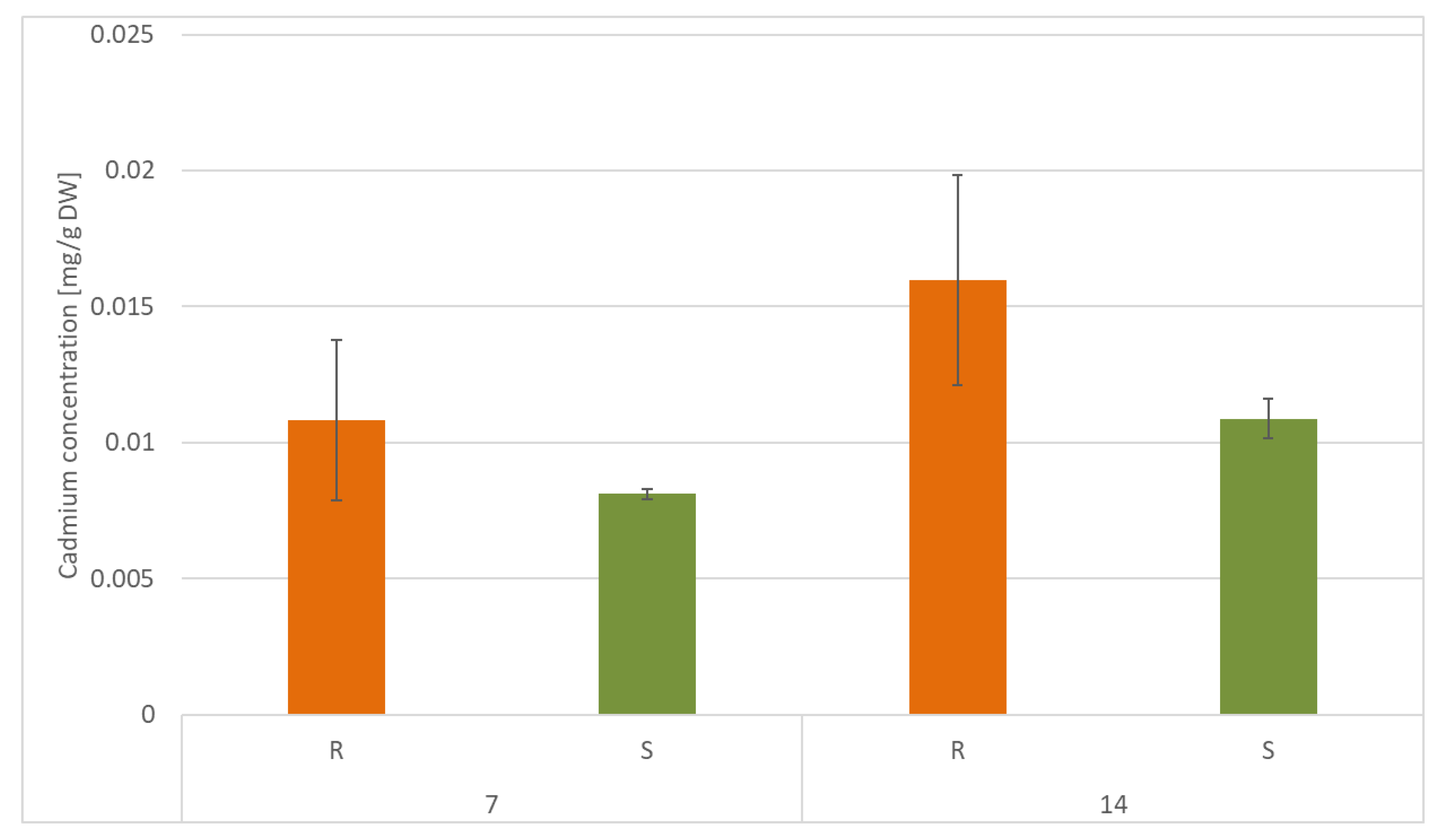
C. officinalis plant roots and hairy root culture growth is in accordance with most of the studies on heavy metal influences on plant development. The reduction of the biomass in plants and plant in vitro cultures exposed to Cd stress can be attributed to the accumulation of this element in the cell wall and its entry into the cytoplasm, leading to the disruption of the normal metabolism, the reduction of the uptake and distribution of other essential elements and nutrients (thus inducing mineral deficiencies and interrupting the activity of important enzymes), and reduction in the cellular turgor and cell division. Plant roots are the first organs directly affected by heavy metals present in soil, and the exerted phytotoxic effects are clearly visible as root growth inhibition and alterations in root morphology. Roots are particularly affected by Cd-induced stress, because Cd can be readily absorbed into the rhizodermis and root cortex, either through apoplastic or symplastic pathways, and then through the plasma membrane of the endodermis, root cell walls are the major sites of Cd storage, and Cd accumulation is, therefore, generally greater in roots compared with the plant’s aerial parts [
22,
29]. The present study clearly shows that in
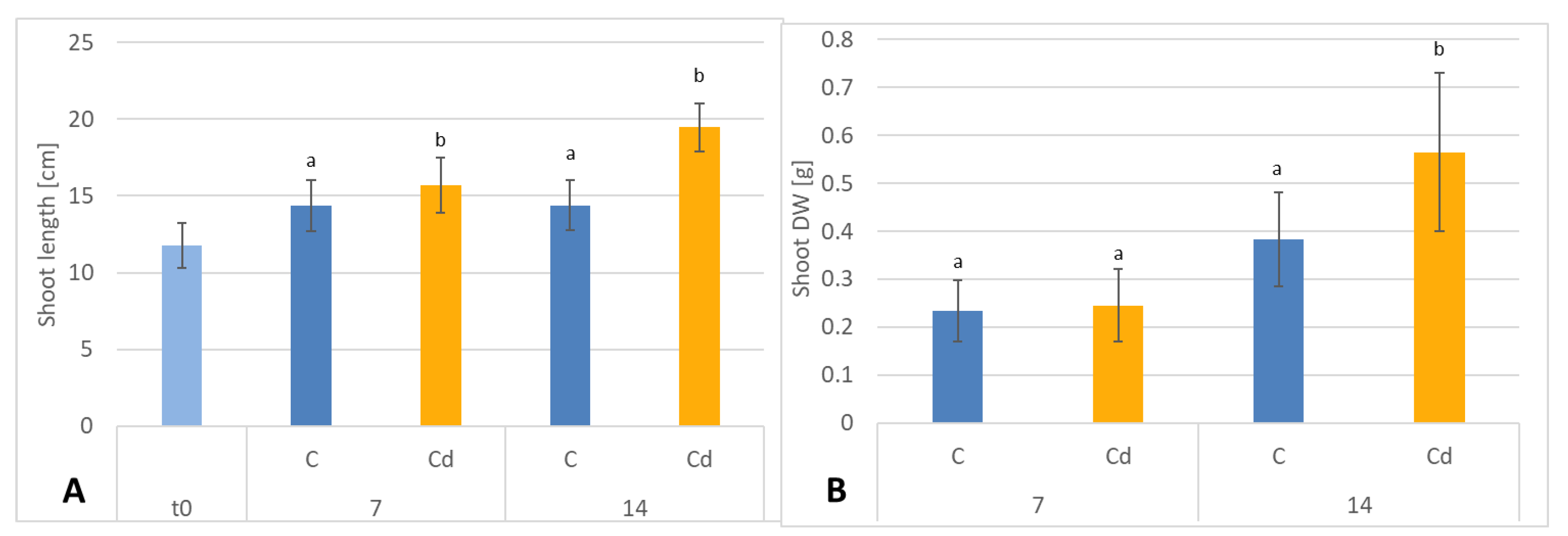
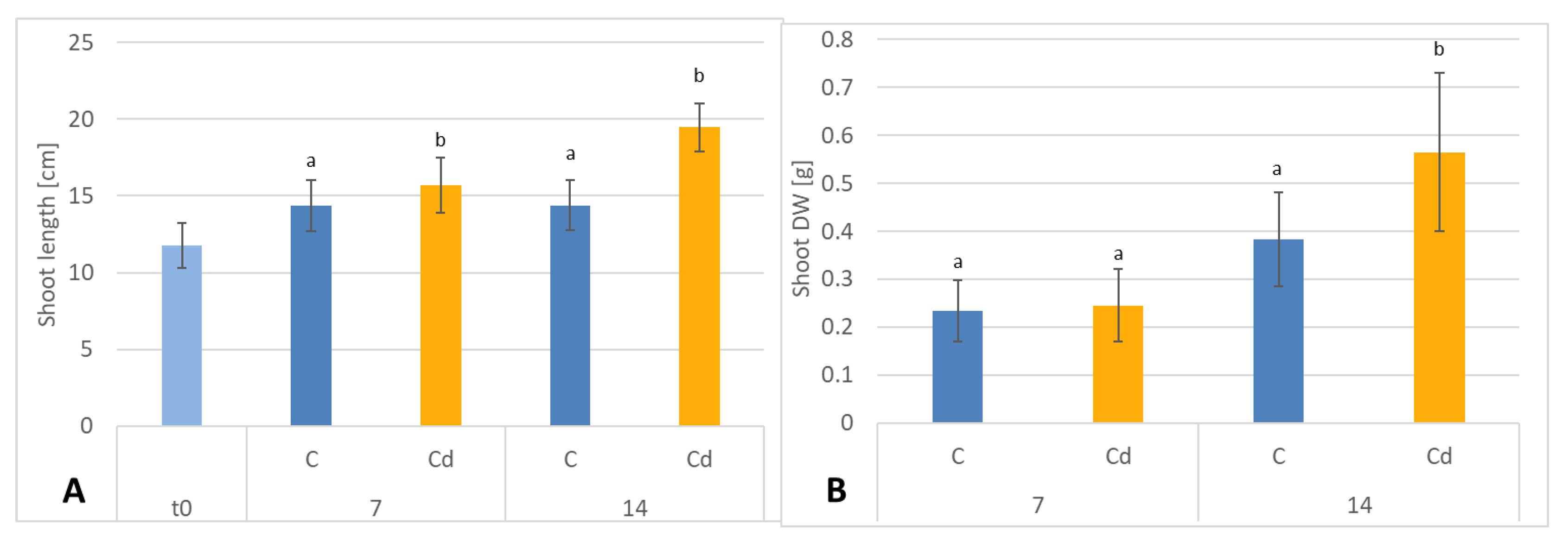
C. officinalis plants, the root is a direct target of Cd stress and is more affected than the shoot. Furthermore, the aerial part of the plants exposed to Cd might not display symptoms of growth reduction; on the contrary, both the biomass and the height of shoots of plants cultivated in Cd-contaminated soil were greater than those of untreated plants. Similar observations have been reported for
C. officinalis exposed to Cd stress, indicating that relatively low Cd concentrations in the soil could stimulate plant growth [
18]. This phenomenon has also been reported in other studies and has been termed “hormesis” by Jia et al. (2015) [
30]. The present study confirms earlier findings that
C. officinalis plants show high tolerance to Cd contamination; as this species is largely used as an ornamental plant and therefore does not reach the human food chain, it might be used for phytoremediation purposes [
18,
19]. However, this implies the distinct understanding that it should not be harvested for other purposes, e.g., as a herbal plant in traditional medicine.
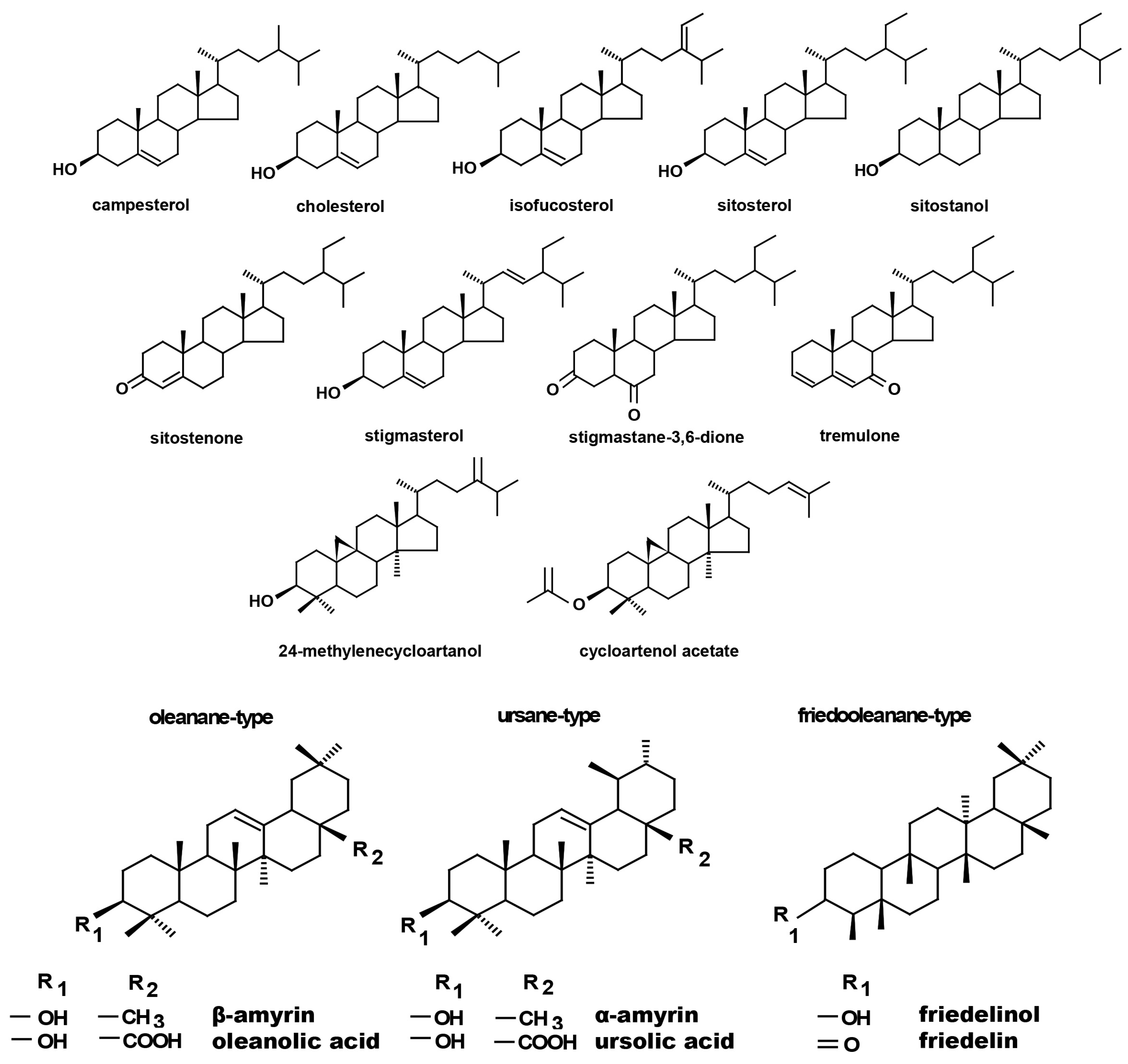
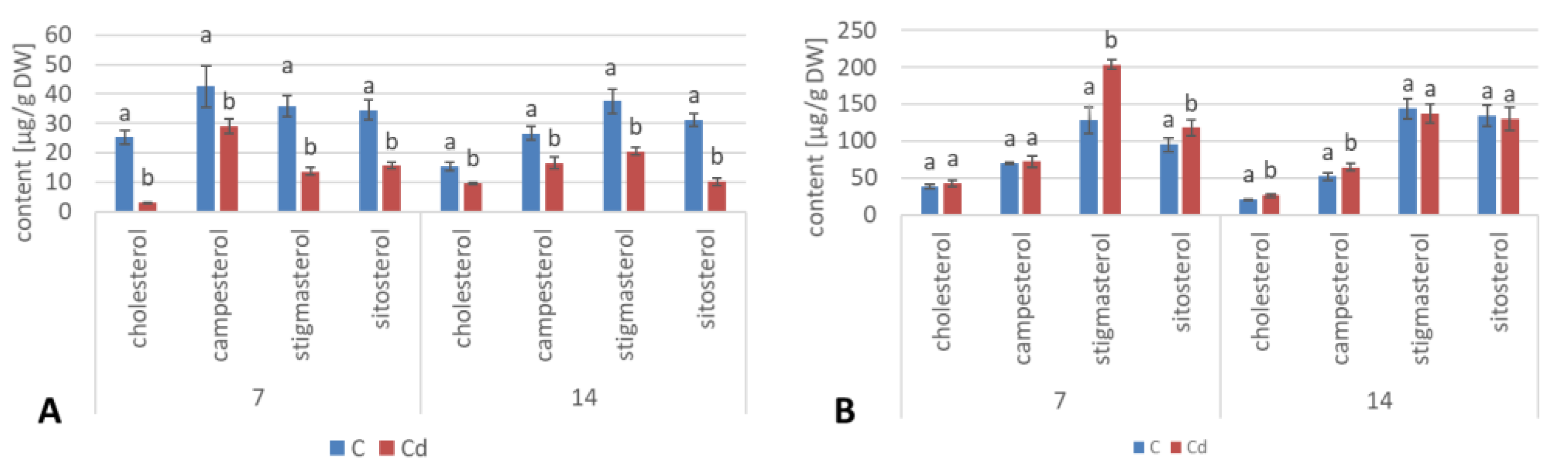
Regardless of the Cd influence on the total sterol content, the present study revealed that Cd stress affected the proportions among individual compounds and the formation of ester and glycoside conjugates. The main observations concerned the changing ratio between stigmasterol and sitosterol, resulting from modulations of sterol side-chain hydrogenation as well as fluctuations in the campesterol content, indicating the diverse stimulation of competing branches in sterol biosynthesis, i.e., 24-methyl sterols versus 24-ethyl sterols. Changes in the sitosterol-to-stigmasterol ratio are frequently observed in plants during adaptation to unfavorable environmental conditions, e.g., under drought stress or salinity [
11,
12]. Stigmasterol, formed from sitosterol in a reaction catalyzed by C-22-sterol desaturase, has an additional double bond in the side chain and can induce substantial changes in the properties of cell membranes correlated with altered activities of membrane-associated enzymes and activation of proton pumps such as H
+-ATPases, the including PM (plasma membrane) proton pump, which are responsible for pH regulation and the production of gradients necessary to maintain cellular homeostasis [
31]. It has been suggested that sterols improve the resistance of plants through reinforcing the cohesion of the cell membrane and may preserve the active conformation of PM H
+-ATPase, modulating its interaction with other constituents, e.g., phospholipids. Although the modulation of PM H
+-ATPase by sterols might be a rather “slow response” because of the complexity of the sterol metabolism, it could nevertheless represent an important mechanism of adaptation to stress conditions [
12,
23]. Ester and glycoside conjugates of sterols are also considered to play a key role in maintaining cell membrane homeostasis, although their correlation with particular stress responses is not always evident [
11]. In the present study, the increase in the content of glycoside conjugates and the decrease of ester forms were noticed, both in roots and hairy root culture, as well as in shoots. However, this phenomenon might not be universal for all plants and requires further studies. Moreover, it is still difficult to ascertain whether the observed changes in the content of sterols and their conjugates appear as a direct effect of Cd toxicity, and, thus, they constitute the primary mechanism of the response to Cd stress; or they are generated as a secondary effect of an interference with other metabolic pathways and modified activity of respective enzymes.
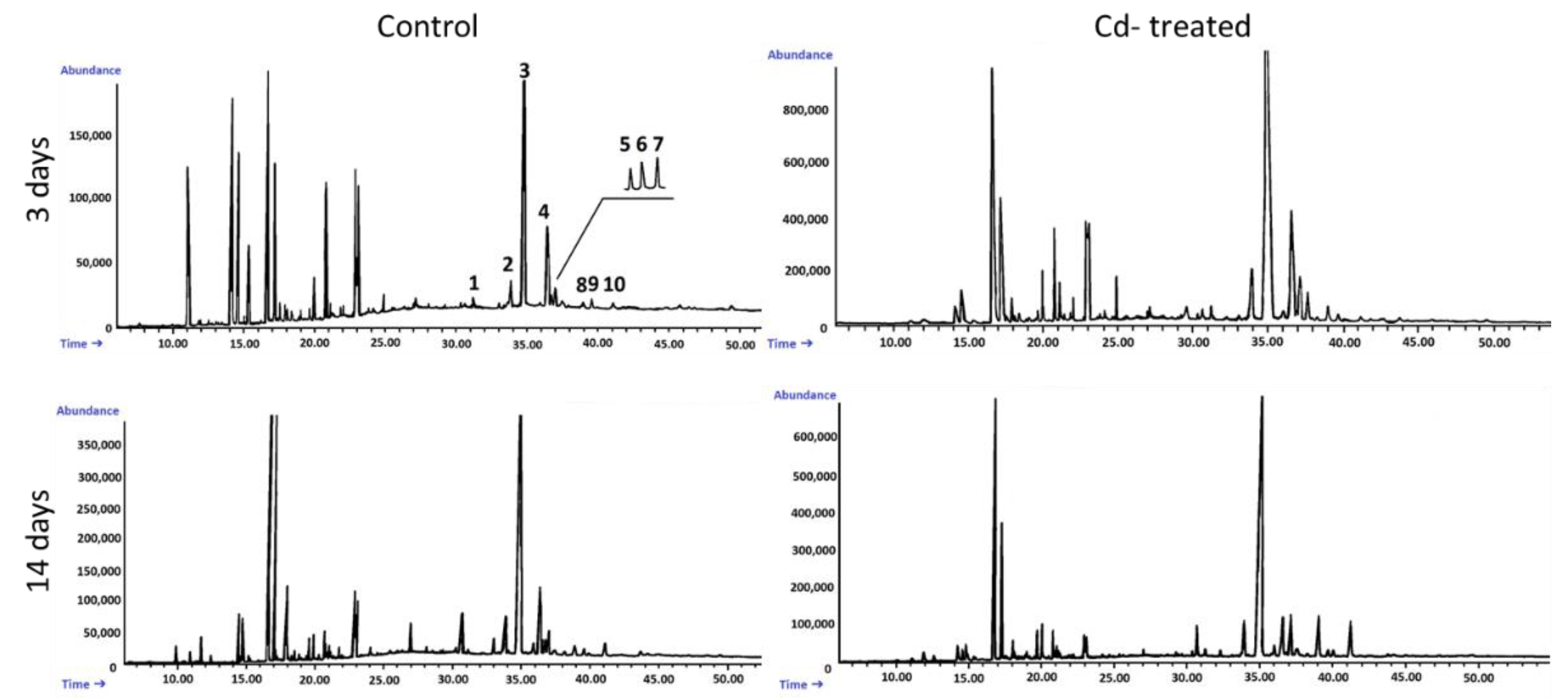
The obtained results clearly demonstrate that the metabolic modifications occurring during stress responses are not constant and equally intensive in time. Some of the observed phenomena could even have an inverse course, first increasing than decreasing, which was particularly evident during a relatively long experiment performed on the hairy root culture. This observation is important to understand that the time-course of plant stress responses is not uniform, the metabolism is a net of pathways influencing each other, and every change can have further consequences for the regulation of the biosynthesis of other compounds. The present study indicates that the majority of the observed modifications finally seem to tend to return to the normal physiological levels, e.g., the sharp boost of sterol biosynthesis observed in plant roots and hairy root culture decreased in time, and the differences between the Cd-treated and control samples largely decreased. However, it should be clearly noticed that the time-course and intensity of stress response differed in whole plants and hairy root cultures; usually the metabolic fluctuations appeared earlier in hairy roots, and were more instantaneous and profound.
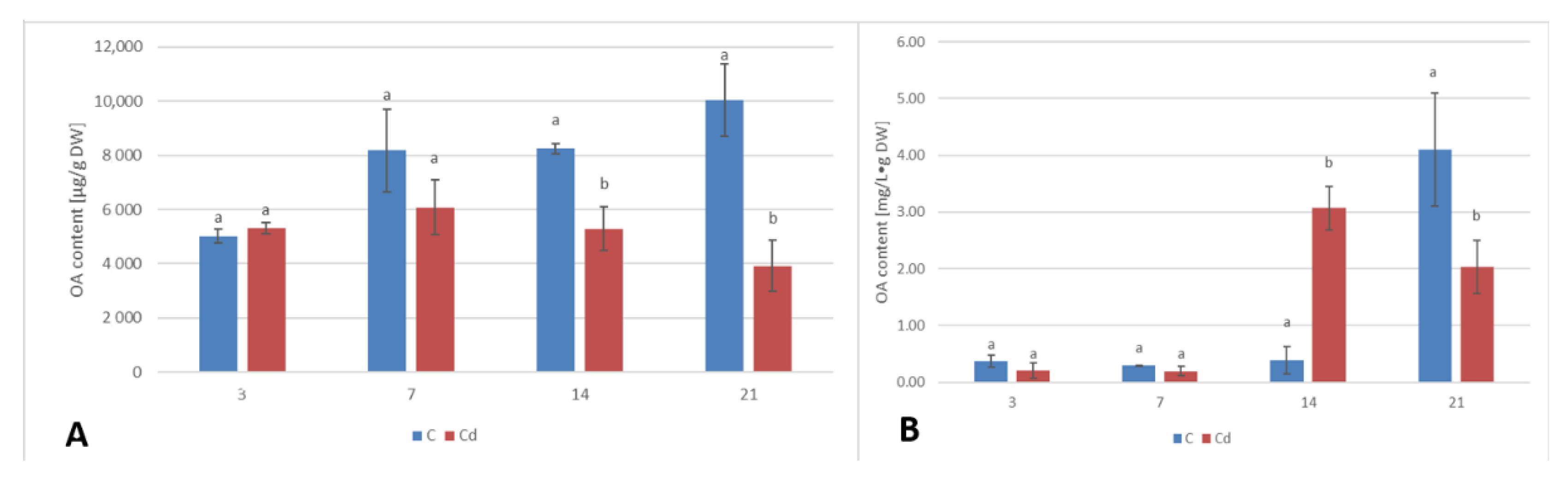
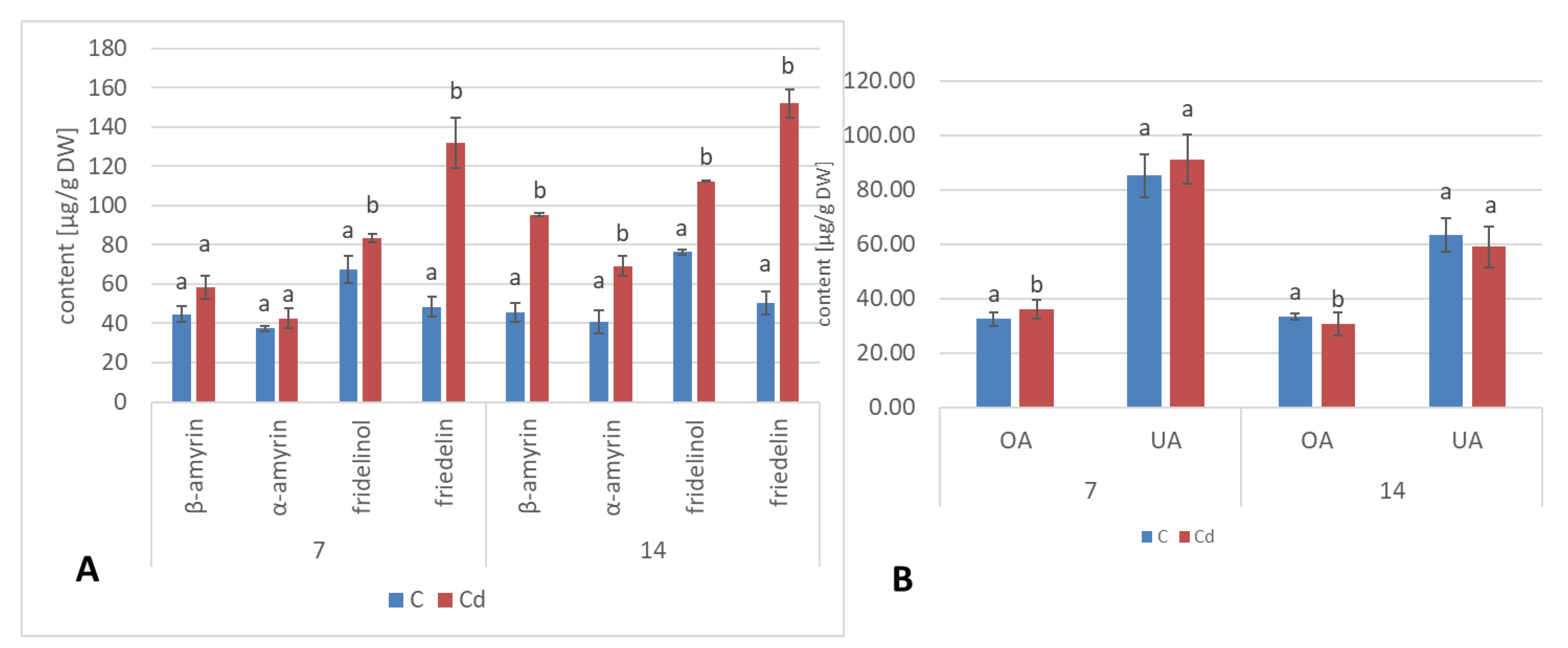
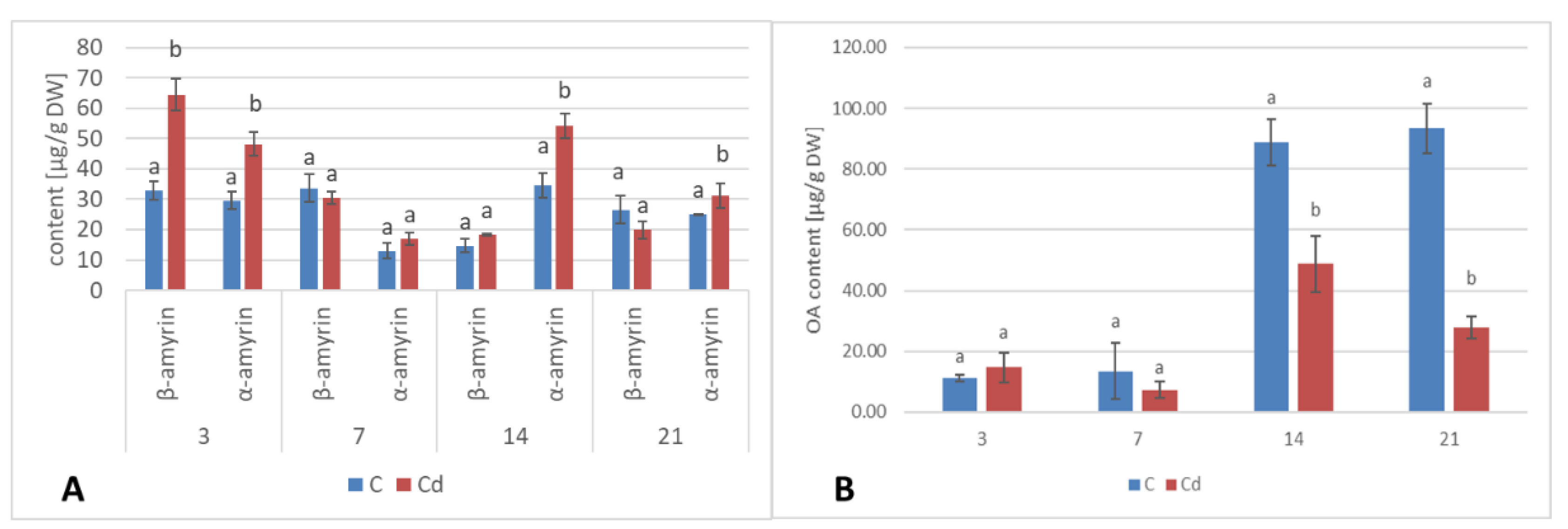
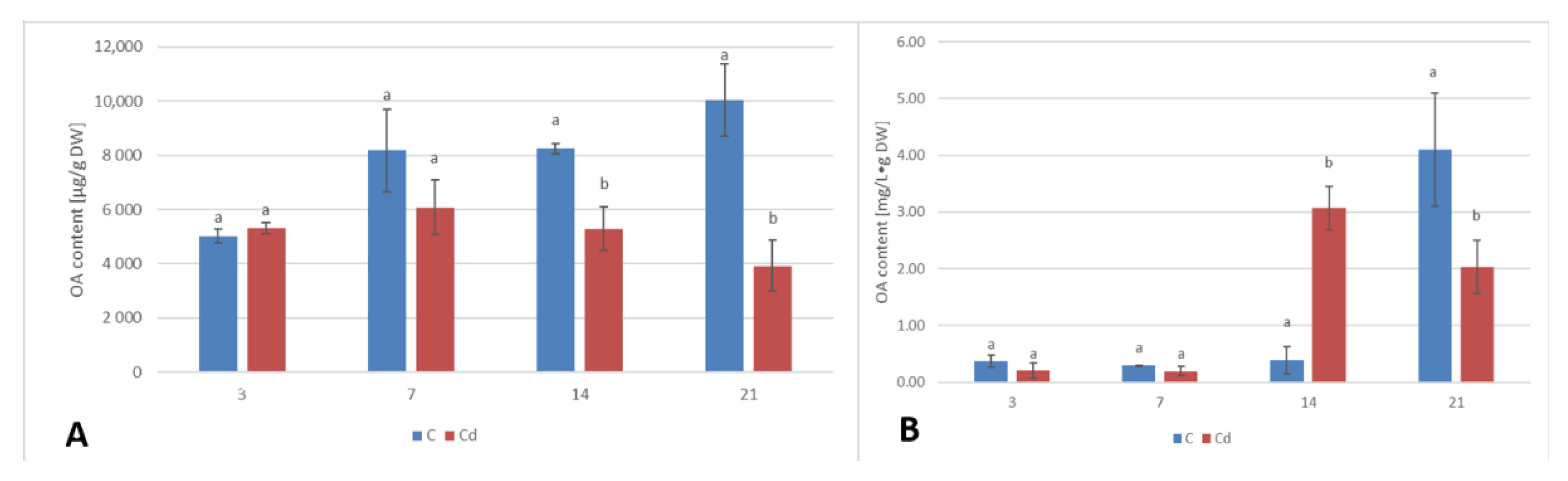
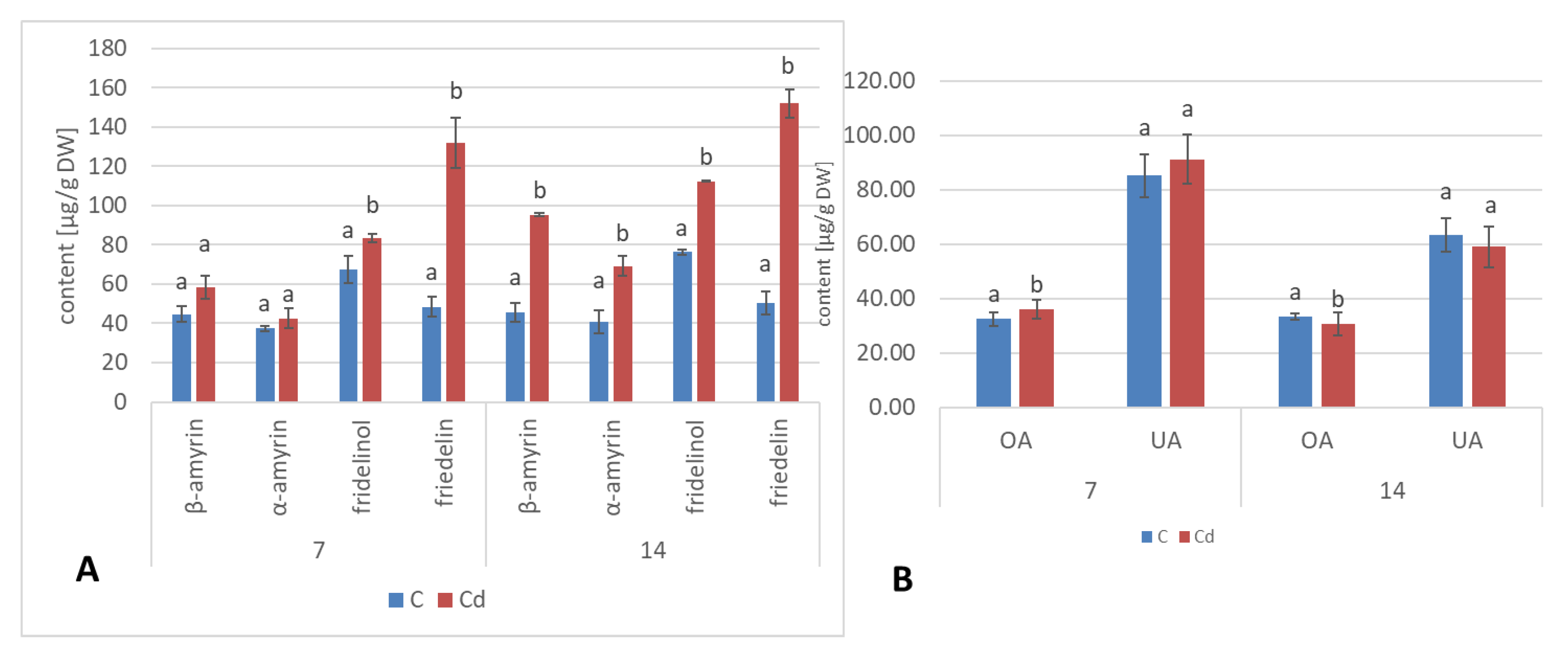
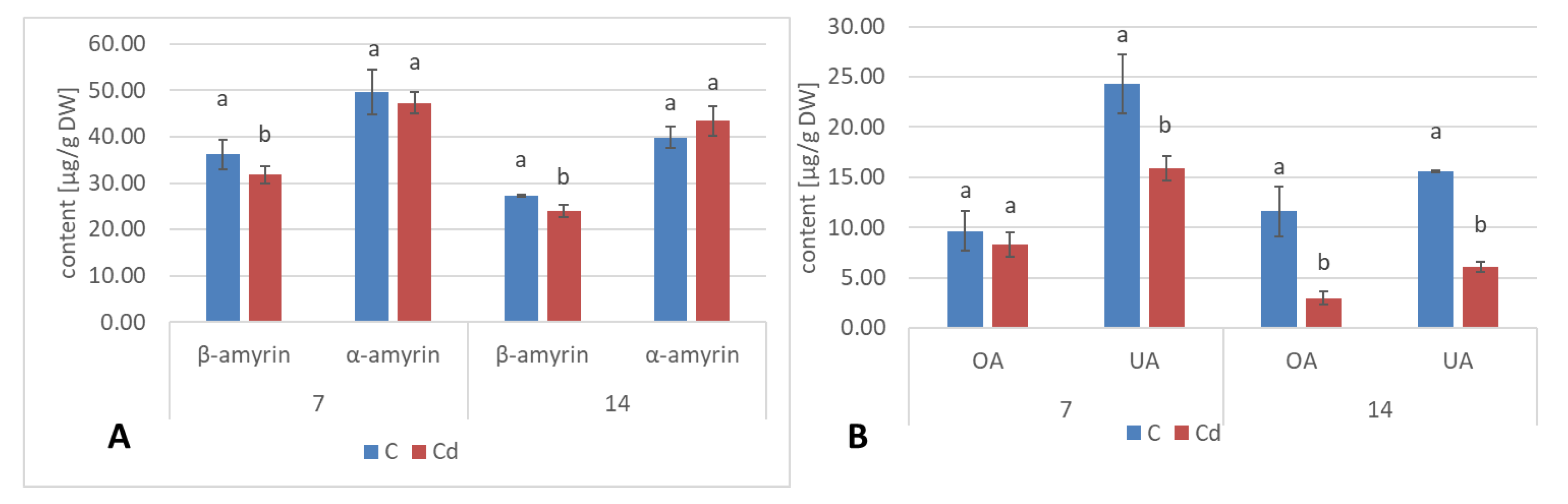
A common phenomenon observed in plants subjected to stress conditions is the competitiveness of the primary/general and secondary/specialized metabolic pathways, including those of steroids and triterpenoids with a common precursor, namely, squalene [
24]. Some symptoms of this competition could be observed in the present study, i.e., the increase in sterol biosynthesis parallel to the decrease in the triterpenoid content in
C. officinalis plant roots and hairy root culture, as well as the inverse phenomenon, namely, the decrease in the sterol content accompanied by the stimulation of triterpenoid biosynthesis in shoots. Effects such as the significant increase in sterol content, changes in the proportion of stigmasterol to sitosterol, and a decrease in the biosynthesis of free oleanolic acid (however, it should be noted that the decreased level of oleanolic acid was coupled with a simultaneous increase in saponin secretion) have also been observed in a preliminary study on the possibility of the application of heavy metals as elicitors for the enhancement of the triterpenoid productivity in
C. officinalis hairy root cultures [
20]. In this sense, the obtained results suggest that sterols and their forms seem to play a greater role in the response to stress associated with the presence of Cd ions than specialized triterpenoid compounds, particularly in roots. This hypothesis is supported by the fact that the changes in sterol content, composition, and proportion between conjugated forms may affect the activity of H
+-ATPases (regardless the mechanism of this modification, i.e., whether it occurs by a direct interaction of sterols with the proton pump, or by indirect alteration of the physical properties of the lipid bilayer), and, hence, influence the membrane transport and ion homeostasis, the processes essential for root cells. In turn, the increased level of α-tocopherol in shoots suggests that an enhanced synthesis of antioxidants is an important mechanism involved in a defensive response to Cd stress in aerial part of the plant.
Another aspect of the present study was the questioned suitability of hairy root cultures as an experimental model to investigate the mechanisms of plant stress responses. Heavy metals including Cd have been applied as efficient elicitors to enhance the biotechnological production of various valuable phytochemicals in plant hairy root cultures, e.g., phytoestrogenic isoflavones daidzein and genistein in
Psoralea corylifolia [
32] or ajmalicine in
Catharanthus roseus [
33]. However, various types of plant in vitro cultures were also often considered as useful experimental models for physiological research, particularly when the main aim was to examine the mechanisms and metabolic responses of plants rather than their agronomic characteristics. Experimental conditions in plant in vitro cultures can be more easily controlled than for soil-growing plants; the relative homogeneity of cultured cells or organs compared with whole plants (in plus to variability between individual plants) improves the reproducibility of results; additionally, the time required for experimental investigations is usually reduced [
34,
35]. The similarity of the metabolic modifications observed in the present study in
C. officinalis plant roots and hairy root culture seems to be encouraging; however, as it has been frequently discussed in various reports, it is important to recognize the limitations associated with the use of in vitro study systems and to be careful before extrapolating the obtained results to plants under field conditions.
4. Materials and Methods
4.1. Plant Material
4.1.1. Hairy Root Cultures
C. officinalis hairy roots line CC16 (derived from cotyledon explant) was obtained according to a previously described procedure [
36]. The roots were cultivated in a ½ Murashige–Skoog liquid medium, at 23–25 °C, in the darkness on a rotary shaker at 120 rpm. Subcultures were performed every 3–4 weeks by transferring the 1–2 cm pieces of the young-branched root to 100 mL of a fresh medium.
4.1.2. Pot Cultures
C. officinalis seeds (PNOS, Ożarów Mazowiecki, Poland) were sown into rectangular plastic pots (dimensions: 80 × 18 × 14 cm) filled with universal flower soil “Athena” (the details of physical and chemical characterization in
Table S14). Germination of seed started within 3 days of sowing. The plants were cultivated for 3 weeks in the greenhouse under controlled conditions (16/8 h day/night fotoperiod, 52 ± 2% humidity, temperature 20 °C with a light intensity of 120 ± 10 µmol/m
2·s).
4.2. Treatment with Cadmium Ions
4.2.1. Supplementation of Hairy Roots
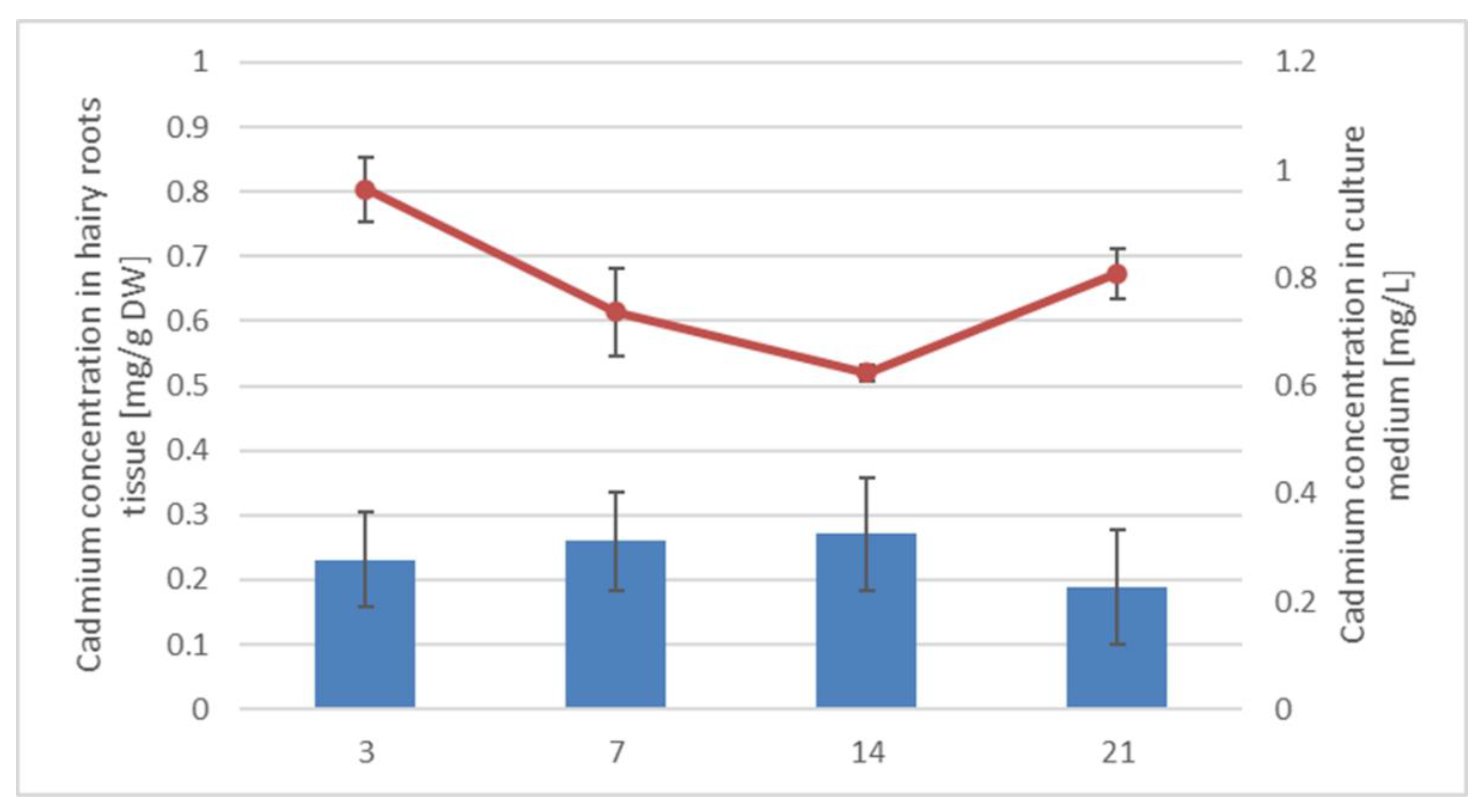
Freshly subcultured roots were incubated for 21 days to obtain at least 1.5 g of fresh weight. Afterwards, they were weighed and transferred to 100 mL of fresh medium five days prior to elicitation. A 50 mM stock solution of CaCl2 was prepared, then the solution was sterilized by filtration through a 0.22 m syringe filter (Millipore, Bionovo, Legnica, Poland) and added to the culture medium to obtain the final concentrations of 25 µmol/L. The cultures were cultivated with cadmium ions for 3, 7, 14, and 21 days.
4.2.2. Supplementation of Pot-Cultures
Soil conditioning with cadmium was conducted by the addition of cadmium chloride salt solution (CaCl2), so that the final concentration of this element in soil was 5 mg Cd/kg dry weight [d.w]. After cadmium contamination, the soil was thoroughly mixed and incubated for 7 days at room temperature. During this time, the soil was mixed and watered every day in order to keep the humidity of the soil stable at 40–45%. Three-week-old seedlings were transferred to cadmium-contaminated soil and cultivated either for 1 week or for 2 weeks in the greenhouse under conditions described above.
4.3. Extraction and Fractionation
4.3.1. Extraction of the Hairy Roots and the Culture Medium
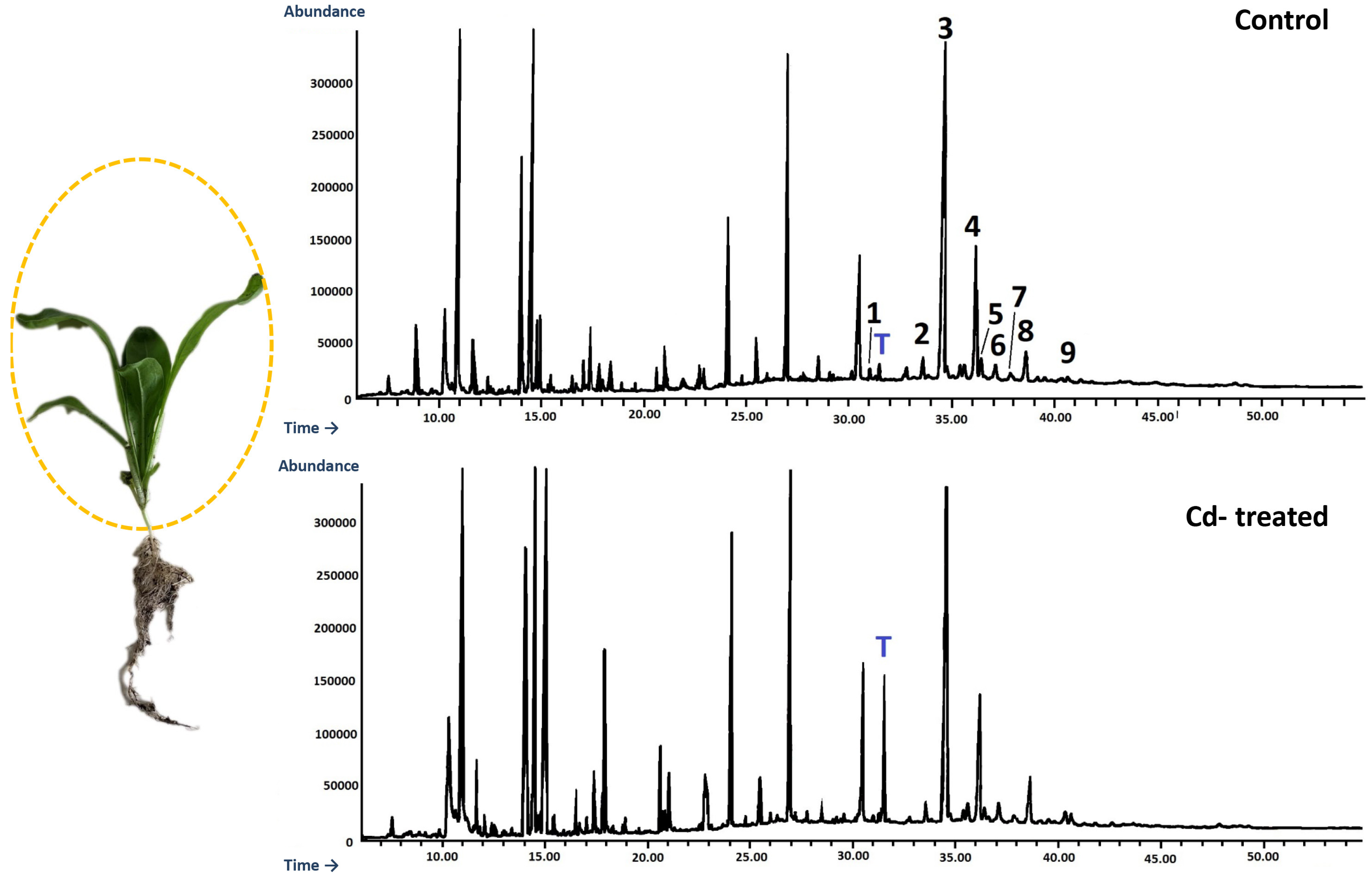
After 3, 7, 14, and 21 days of incubation with cadmium, the culture media were filtered from the hairy roots. The harvested hairy roots were air-dried at room temperature before extraction, whereas the culture media were directly extracted 3 times with 40 mL portions of n-butanol to extract oleanolic acid saponins released to the medium. Dried hairy roots were powdered and extracted using a Soxhlet apparatus for 8 h with diethyl ether to obtain fractions of free sterols, conjugated sterols (esters and low polar glycosides), free triterpenoid acids and alcohols, and then 8 h with methanol to extract more polar sterol glycosides and oleanolic acid saponins. The obtained extracts were evaporated to dryness under reduced pressure on a rotary evaporator.
4.3.2. Extraction of the Plants
After 7 and 14 days of growing in cadmium-contaminated soil, plants were gently collected, then weighed, and lengths of the root and the shoots (aerial) parts were measured. Roots and aerial parts were left to dry in the dark and airy place, afterwards weighed again, grinded in mortar to fine powder and extracted in Soxhlet apparatus for 8 h with diethyl ether and then 8 h with methanol. The obtained extracts were evaporated to dryness under reduced pressure on a rotary evaporator.
4.4. Fractionation of Diethyl Ether Extracts
Evaporated diethyl ether extracts obtained from the hairy roots, as well as roots and aerial parts of native plants, were fractionated by adsorption preparative TLC on 20 cm × 20 cm glass plates coated manually with silica gel 60H (Merck, Darmstadt, Germany). The solvent system chloroform: methanol 97:3 (
v/
v) was applied for developing. Four fractions were obtained as described by Sykłowska-Baranek et al. (2022): (i) esters (Rf, retention factor, of 0.9–1); (ii) free steroids and neutral triterpenoids (Rf of 0.3–0.9); (iii) free triterpenoid acids (Rf 0.2–0.3); and (iv) glycosides (Rf of 0–0.2) [
28]. Fraction (ii) containing free steroids and neutral triterpenoids (alcohols) was directly analyzed using GC–MS, gas chromatography-mass spectrometer (Agilent Technologies 7890A); the triterpenoid acid fraction (iii) was methylated with diazomethane prior to GC-MS analysis as described previously [
37]; the ester fraction (i) was subjected to alkaline hydrolysis to release the steroid core from ester forms; and the glycoside fraction (iv) to acidic hydrolysis to release the aglycones.
4.5. Alkaline Hydrolysis
The ester fractions were subjected to alkaline hydrolysis with 10% NaOH in 80% methanol at 80 °C for 3 h. Subsequently, 5 volumes of water were added to each hydrolysate, the pH was neutralized with 5% acetic acid, and the obtained mixtures were extracted with diethyl ether (3 × 20 mL) in a separation funnel. The extracts were evaporated and fractionated by preparative TLC (as described in
Section 4.4) to obtain the fraction of sterols released from their ester forms.
4.6. Acidic Hydrolysis
The glycoside fractions obtained from the diethyl ether extracts, evaporated methanol extracts and n-butanol extracts from the culture medium were hydrolyzed by 11% HCl in 80% methanol during 2 h on a heating mantle under reflux [
28,
37]. Subsequently, the hydrolysates were diluted with distilled water, methanol was evaporated in a rotary evaporator, and the obtained aqueous remnants were extracted 3 times with 40 mL portions of diethyl ether in a separation funnel. The obtained extracts were washed with distilled water 3 times and evaporated to dryness.
4.7. Fractionation of Acidic Hydrolysates
The dried extracts obtained from acidic hydrolysates were divided by preparative TLC on 20 cm × 20 cm glass plates, manually coated with silica gel 60H (Merck, Darmstadt, Germany). The solvent system chloroform: methanol 95:5 (v/v) was used for developing the plates. From hydrolyzed methanol extracts, two fractions were obtained: sterols and oleanolic acid. From hydrolyzed n-butanol extracts, only one fraction was obtained: oleanolic acid. Purified oleanolic acid was methylated with diazomethane.
4.8. Derivatization of Triterpenoid Acids
Nitrosomethylurea (2.06 g) was added to a mixture of 20 mL of diethyl ether and 6 mL of 50% aqueous KOH, and the organic layer was then separated from the aqueous layer. Samples containing triterpenoid acids were dissolved in 2 mL of the obtained solution of diazomethane in diethyl ether, and held at 2 °C for 24 h.
4.9. Quantification of Oleanolic Acid by GC
The samples were solved in the suitable portion of the mixture diethyl ether: methanol 3:1 (
v/
v). Quantitative measurement of oleanolic acid (in the form of methyl ester) was performed by gas-liquid chromatography (GLC) at 270 °C on a Shimadzu GC-2014 instrument equipped with a flame ionization detector. Samples were applied by split injection 1:5 on a ZB-1 30 m × 0.25 mm × 0.25 μm column (Phenomenex, Torrance, CA, USA). The temperature of the injector and detector was 290 °C. Nitrogen was used as the carrier gas at a flow rate of 1.2 mL/min. Peak identification and quantification of oleanolic acid were carried out by referring to a calibration curve prepared with an authenticated sample of methylated oleanolic acid as the standard [
38].
4.10. Identification and Quantification of Triterpenoids by GC–MS/FID
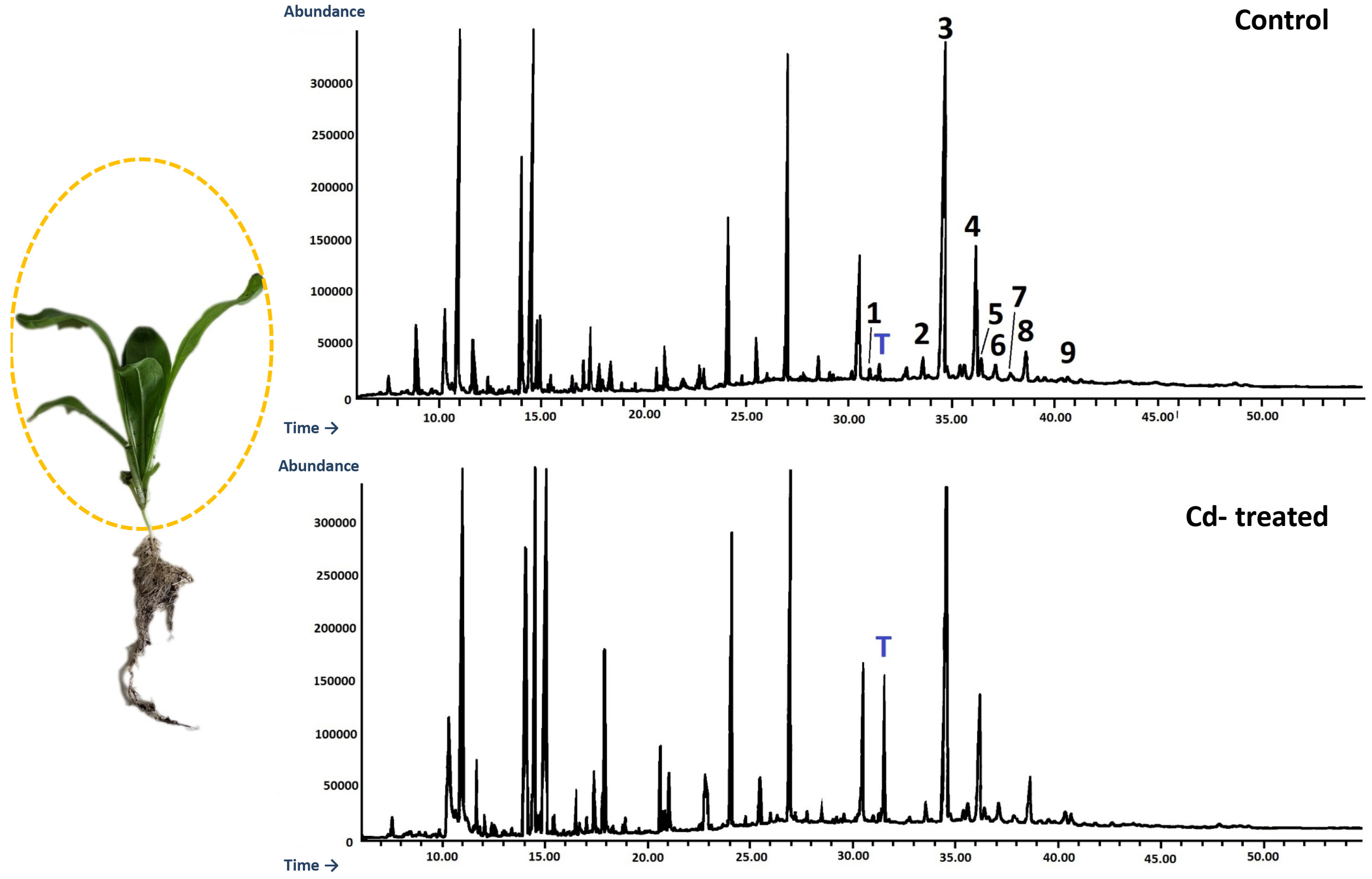
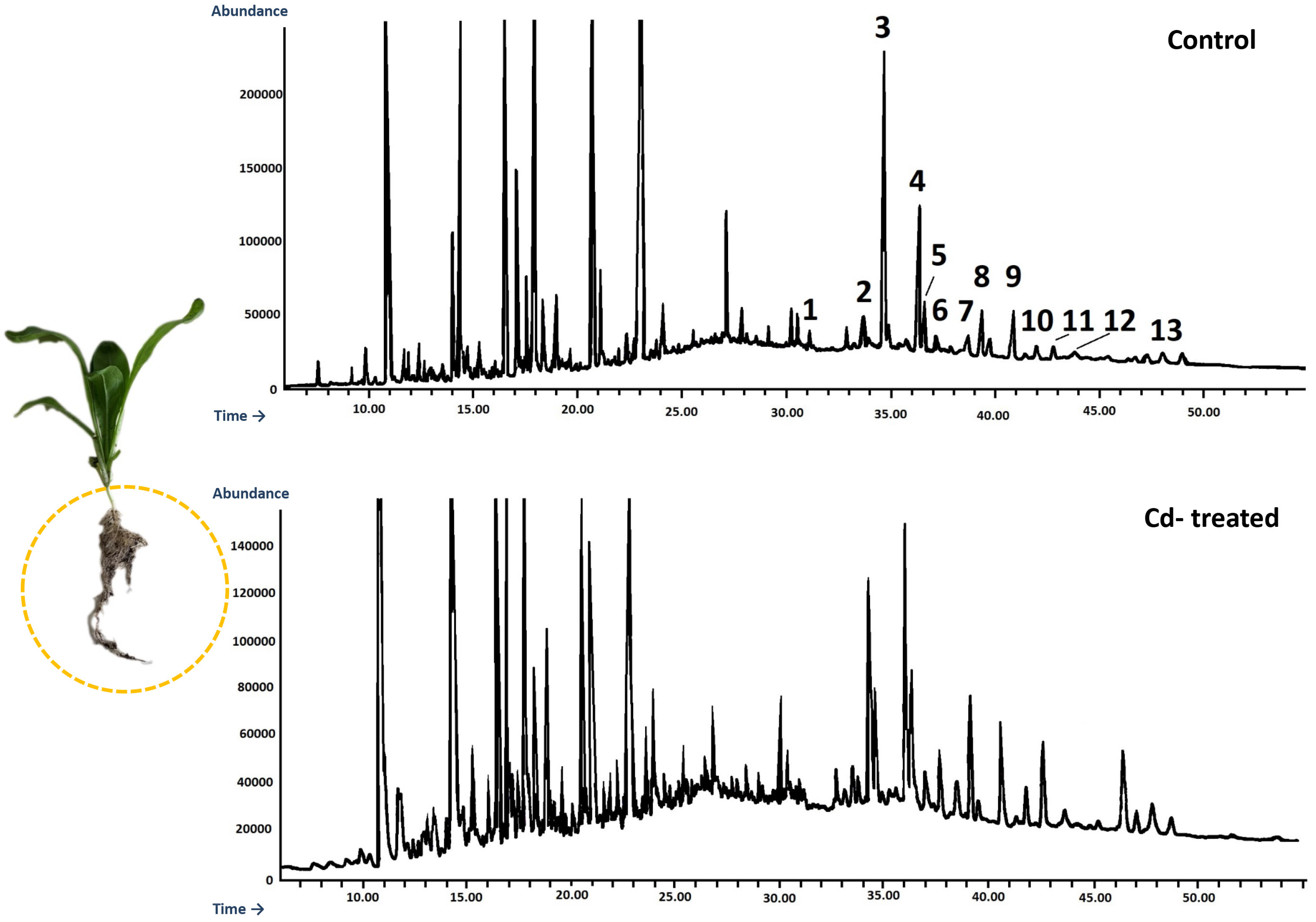
An Agilent Technologies 7890 A gas chromatograph equipped with a 5975C mass spectrometric detector was used for qualitative and quantitative analyses. Samples dissolved in diethyl ether:methanol (5:1, v/v) were applied (in a volume of 1–4 μL) using 1:10 split injection. The column used was a 30 m × 0.25 mm i.d., 0.25-μm, HP-5MS UI (Agilent Technologies, Santa Clara, CA, USA). Helium was used as the carrier gas at a flow rate of 1 mL/min. The separation was made either under isothermal conditions at 280 °C or in the temperature programmed: initial temperature of 160 °C held for 2 min, then increased to 280 °C at 5 °C/1 min and the final temperature of 280 °C held for further 44 min. The other employed parameters were as follows: inlet and FID (flame ionization detector) temperature 290 °C; MS transfer line temperature 275 °C; quadrupole temperature 150 °C; ion source temperature 230 °C; EI 70 eV; m/z range 33–500; FID gas (H2) flow 30 mL·min−1 (hydrogen generator); and air flow 400 mL·min−1. Individual compounds were identified by comparing their mass spectra with library data from Wiley 9th ED. and NIST 2008 Lib. SW Version 2010 or previously reported data and by comparison of their retention times and corresponding mass spectra with those of authentic standards, when available. Quantitation was performed using an external standard method based on calibration curves determined for the compounds belonging to representative triterpenoid classes: α-amyrin for triterpene alcohols, oleanolic acid methyl ester for triterpene acid methyl esters, and sitosterol for steroids.
4.11. Determination of Total Cadmium Concentration in Plant Tissues
To determine the total cadmium concentration in tissues, approximately 0.3 g of sample was digested in 10 mL of the oxidizing mixture: 9 mL of 69% HNO3 (Merck, Darmstadt, Germany) and 1 mL H2O2 (35%) at 180 °C for 30 min using the closed microwave system (Milestone Ethos Plus). Digested samples were transferred to plastic tubes and stored below 4 °C before measurement. The amount of cadmium was measured by Flame Atomic Absorption Spectroscopy (FAAS) using a Thermo Scientific—SOLAAR M Series (TJA Solution, SOLAAR M, UK) at the primary wavelength 228.8 nm. In FAAS the gas mixture was air and acetylene. The calibration curve range was 0–5 mg L−1 and the lower limit of quantification was 0.1 mg L−1. For background correction was used deuterium lamp (TJA Solution, SOLAAR M, UK). Cadmium standards solutions (Merck, Darmstadt, Germany) were prepared in 3% HNO3. The analysis was performed in Laboratory of Environmental Instrumental Analyzes at Faculty of Biology, University of Warsaw.
After determining the Cd concentrations in plants, the translocation factor (
TF) was calculated using following equation:
TF allows to determine heavy metal accumulation potential of plant. In plants showing features of hyperaccumulators the
TF > 1 [
18].
4.12. Statistical Analysis of Data
All data are presented as the means ± standard deviation of three independent samples analyzed in triplicate. For statistical analysis, t-Student’s test was applied using Microsoft Excel by Microsoft, Redmont, WA, USA, and STATISTICA by TIBCO Software Inc., Palo Alto, CA, USA. Statistical significance was considered at p < 0.05.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}