
Locally Secreted Semaphorin 4D Is Engaged in Both Pathogenic Bone Resorption and Retarded Bone Regeneration in a Ligature-Induced Mouse Model of Periodontitis

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
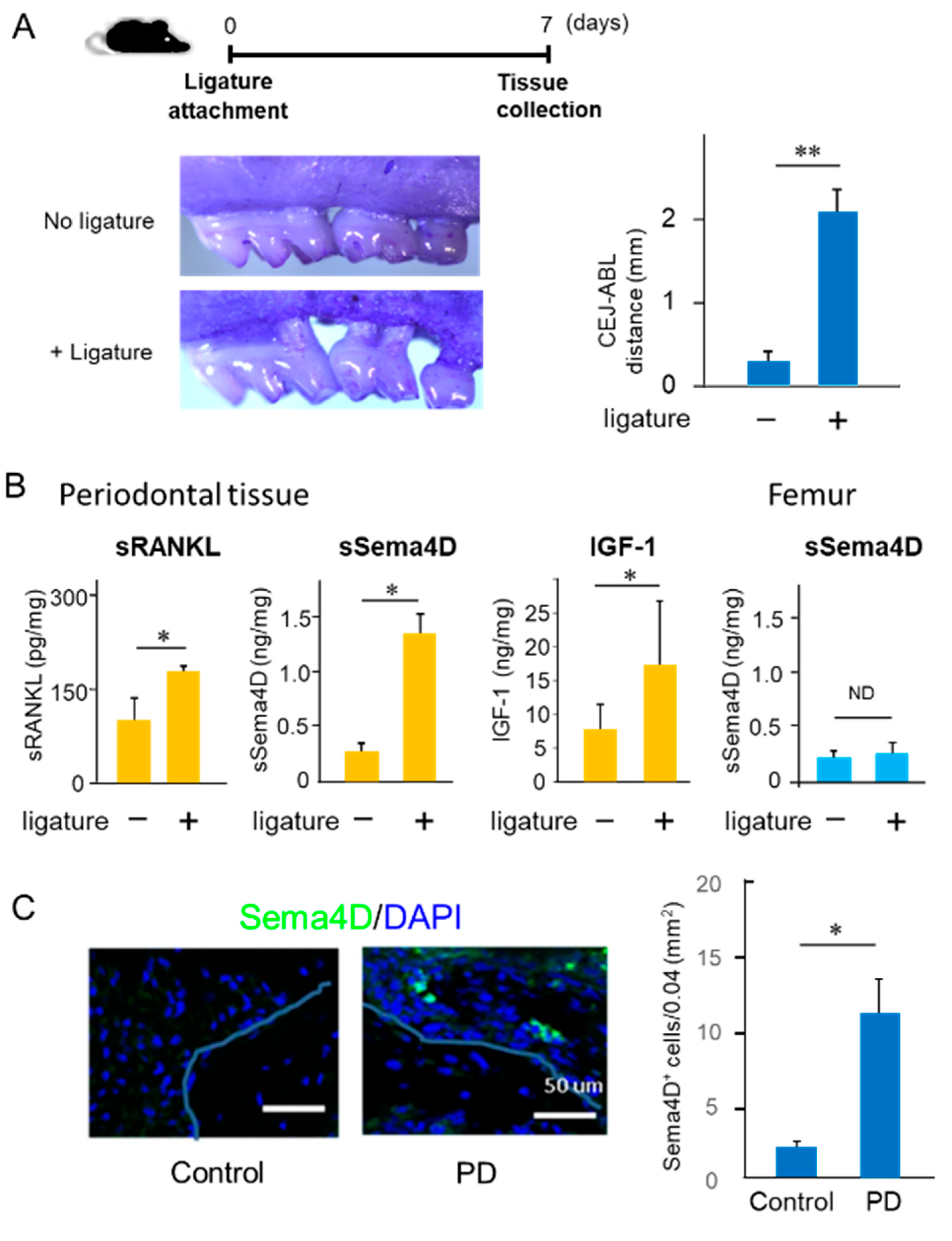
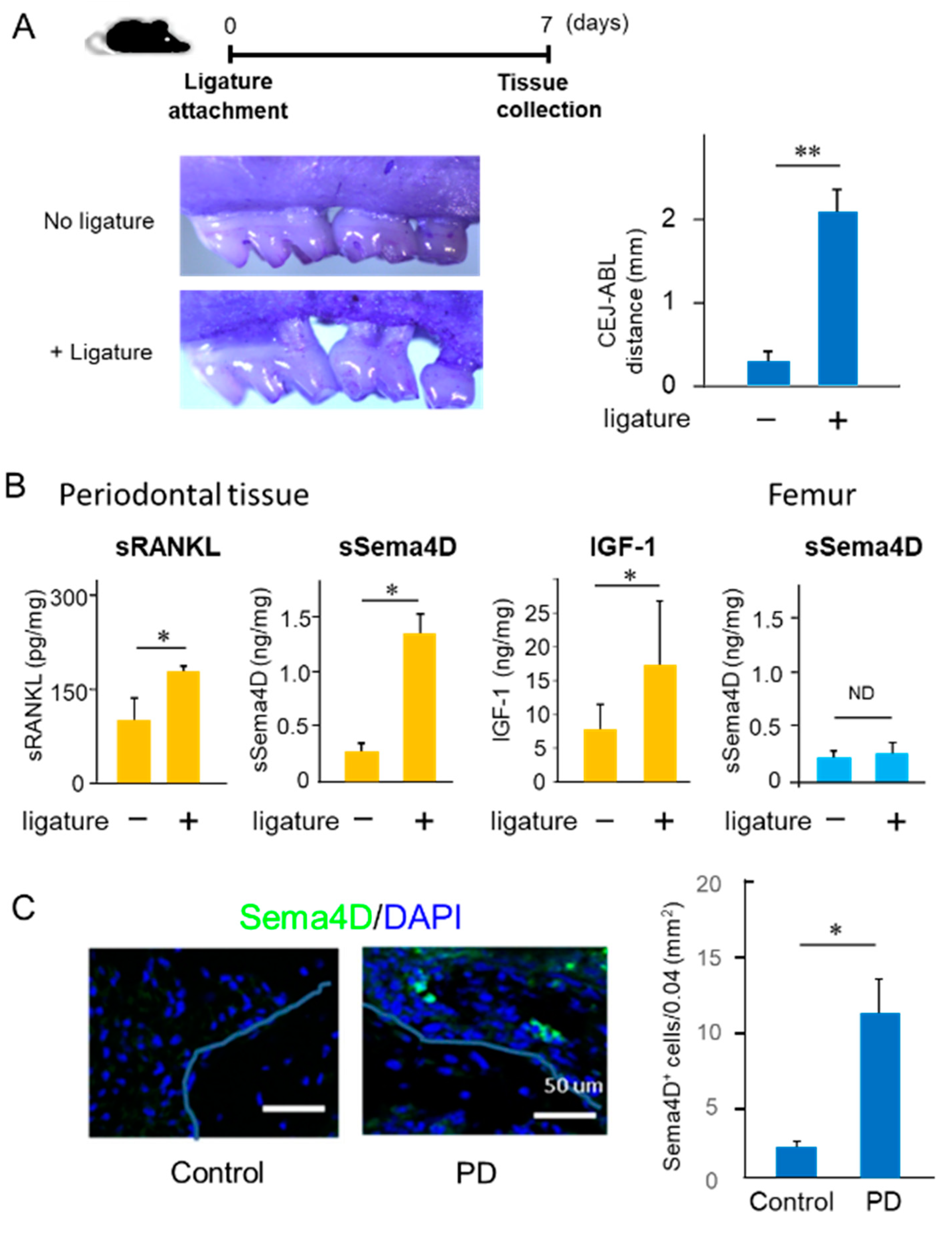
2.1. Increased Production of Sema4D in Mouse Periodontal Tissue of Experimentally Induced PD
2.2. Characteristics of Anti-Sema4D mAb in Reaction to Sema4D Produced by Osteoclasts
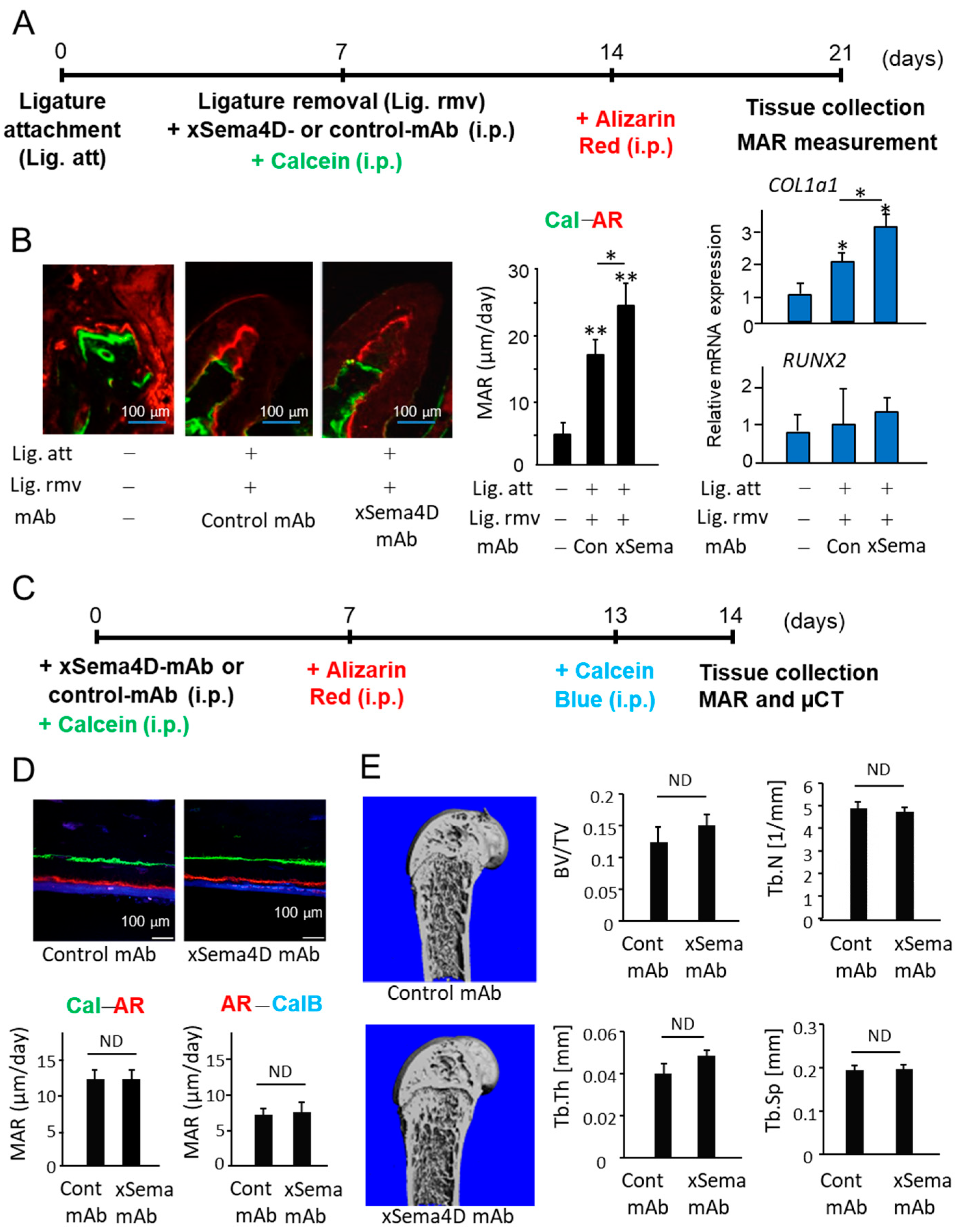
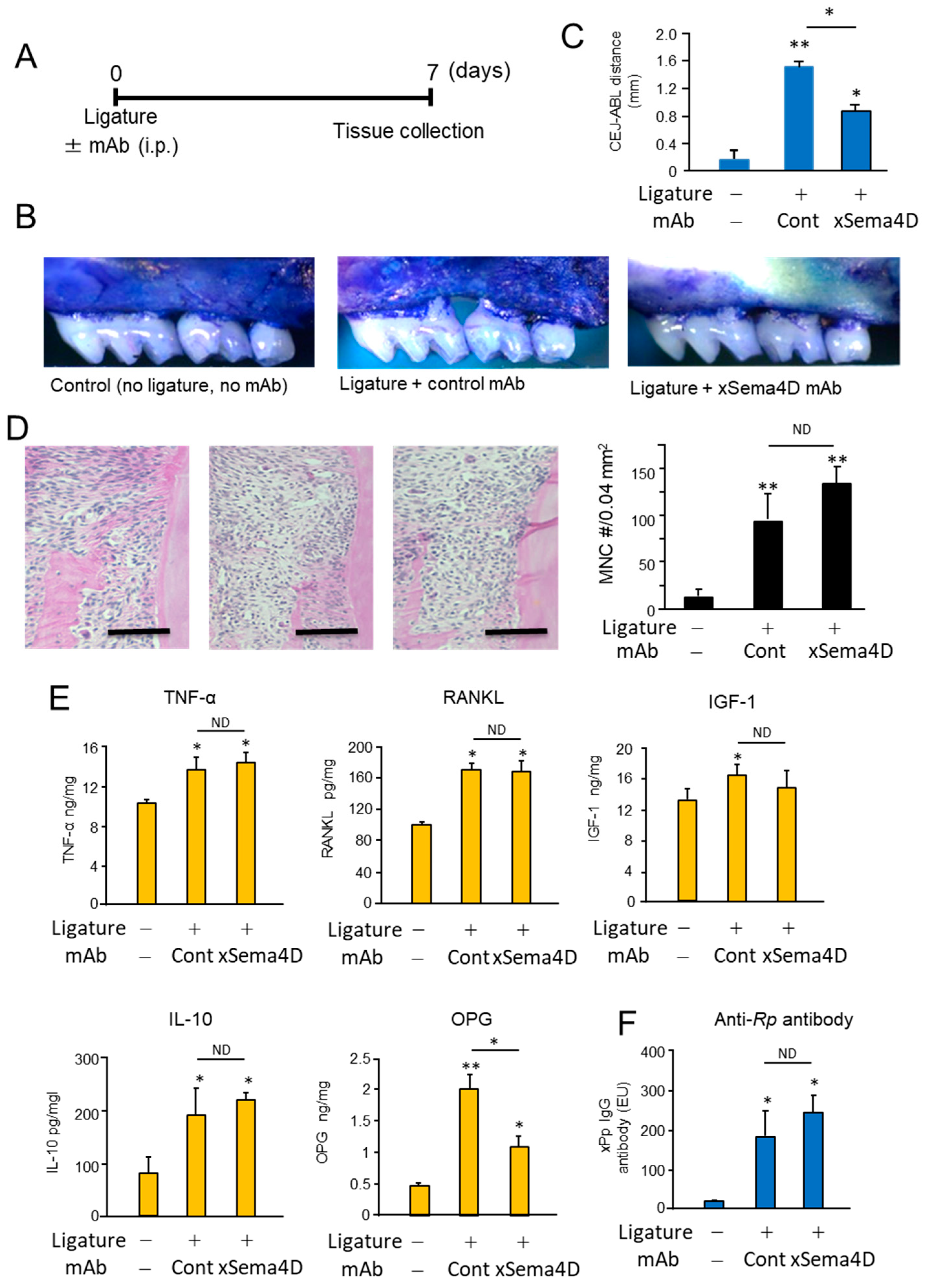
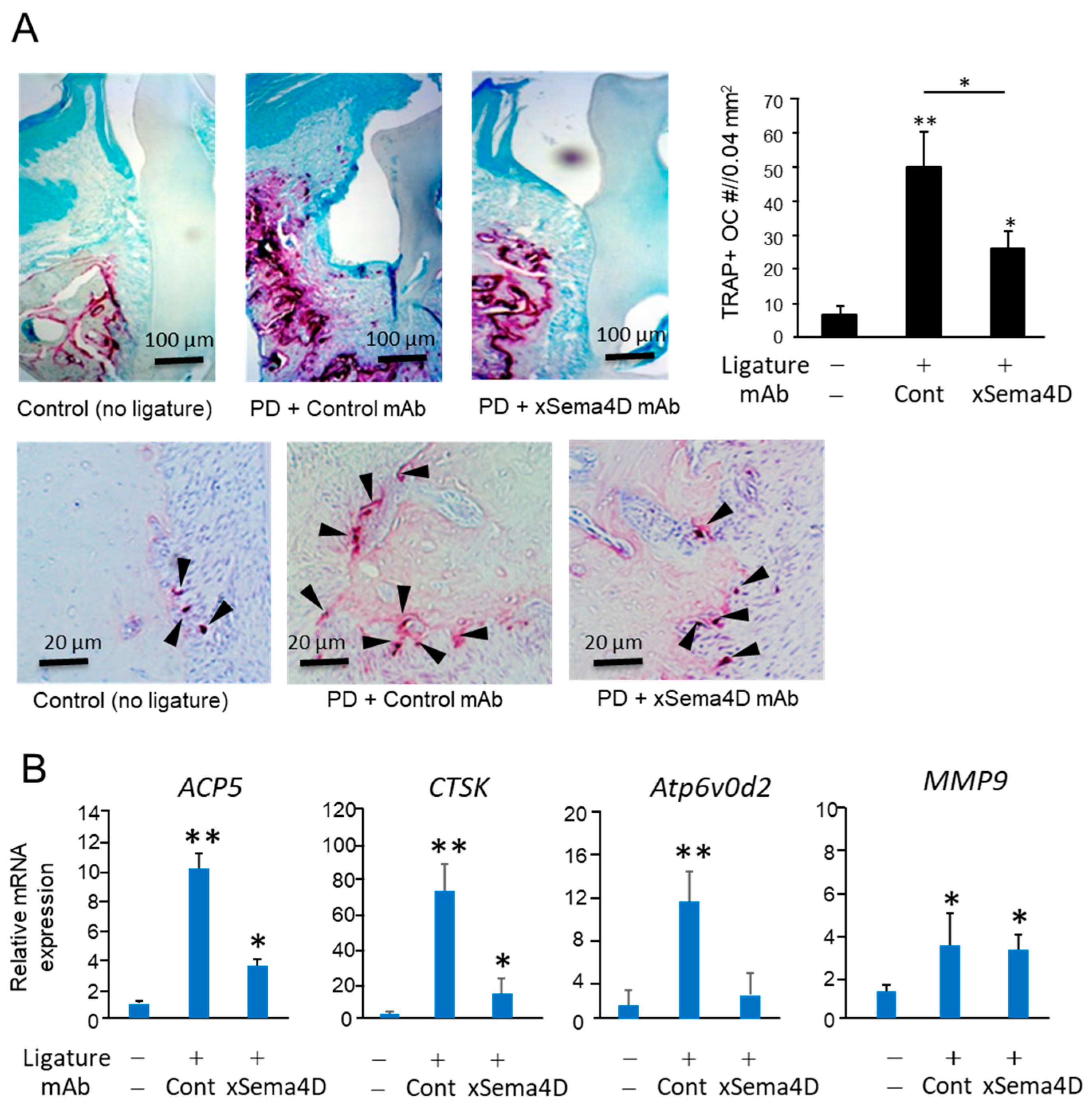
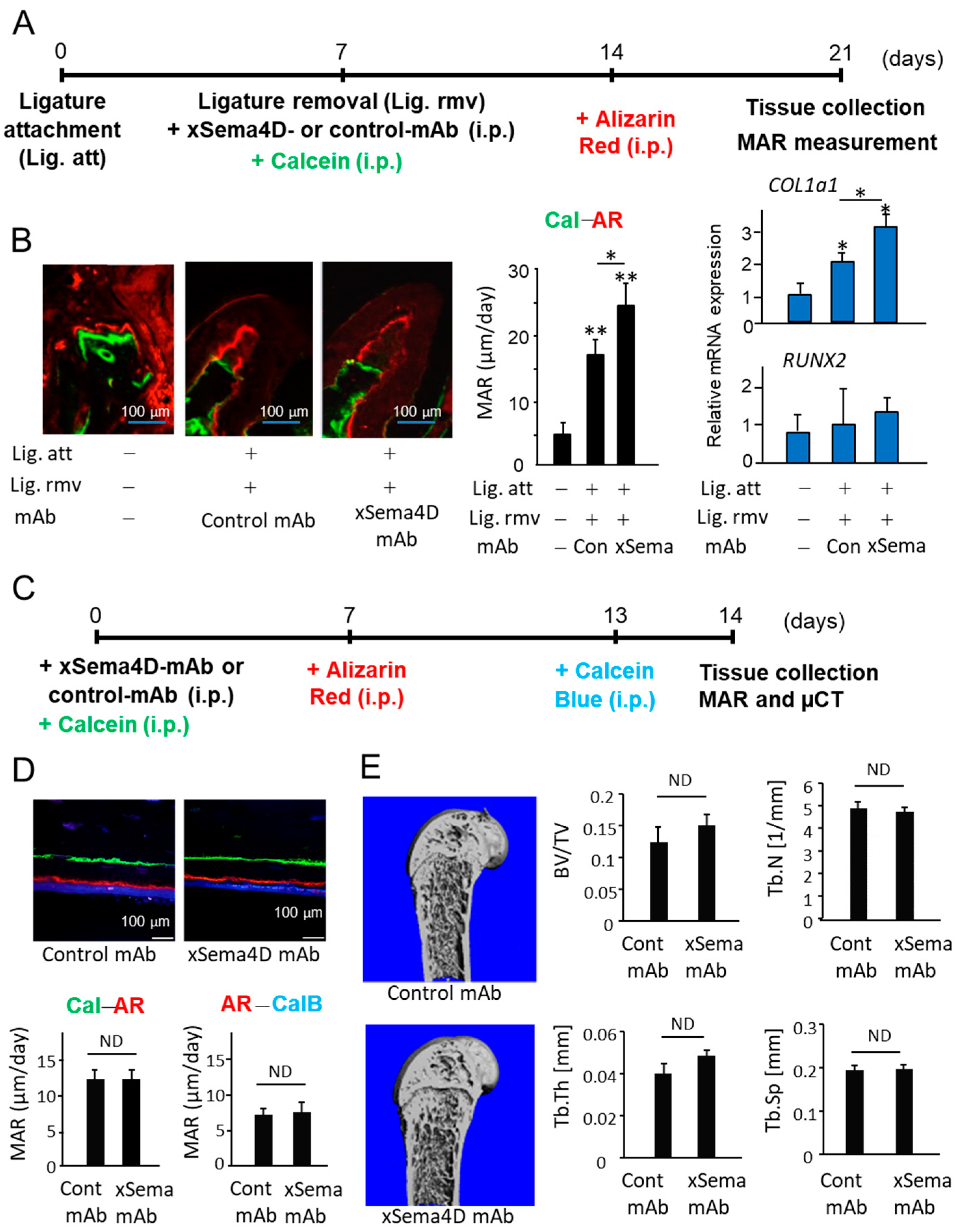
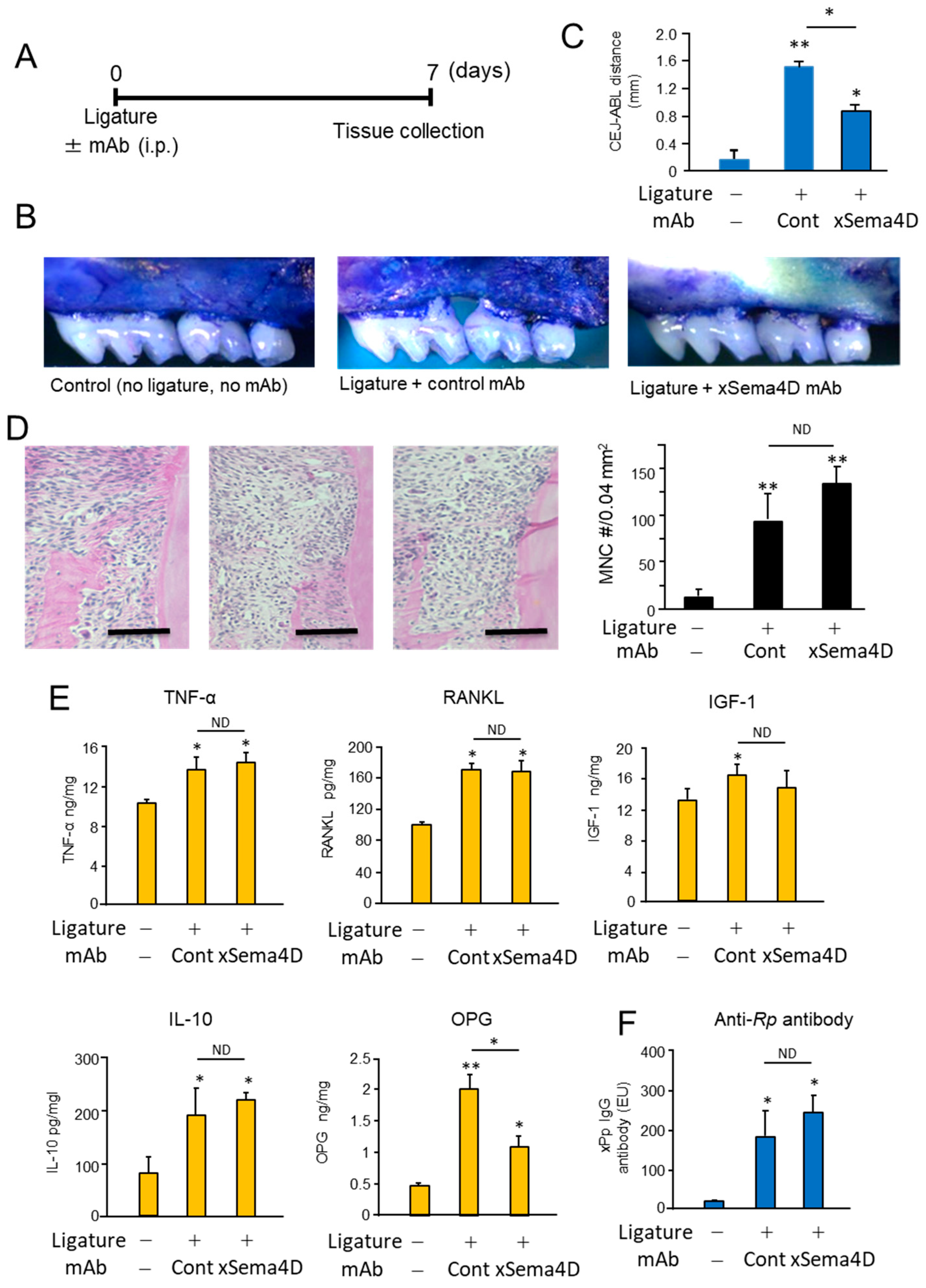
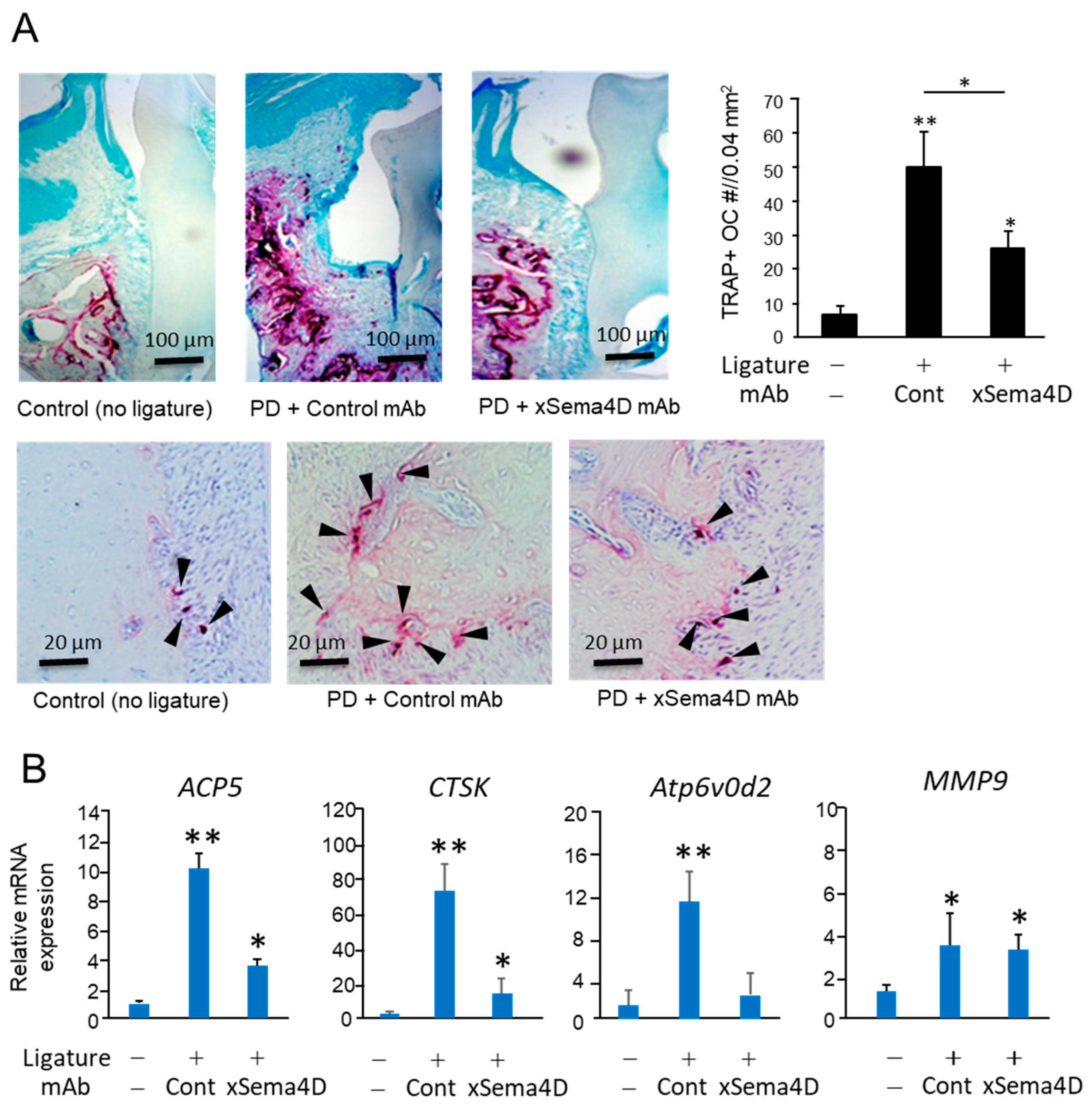
2.3. Locally Elevated Expression of Sema4D in PD Lesion May Be Engaged in the Upregulation of Osteoclast-Mediated Periodontal Bone Loss
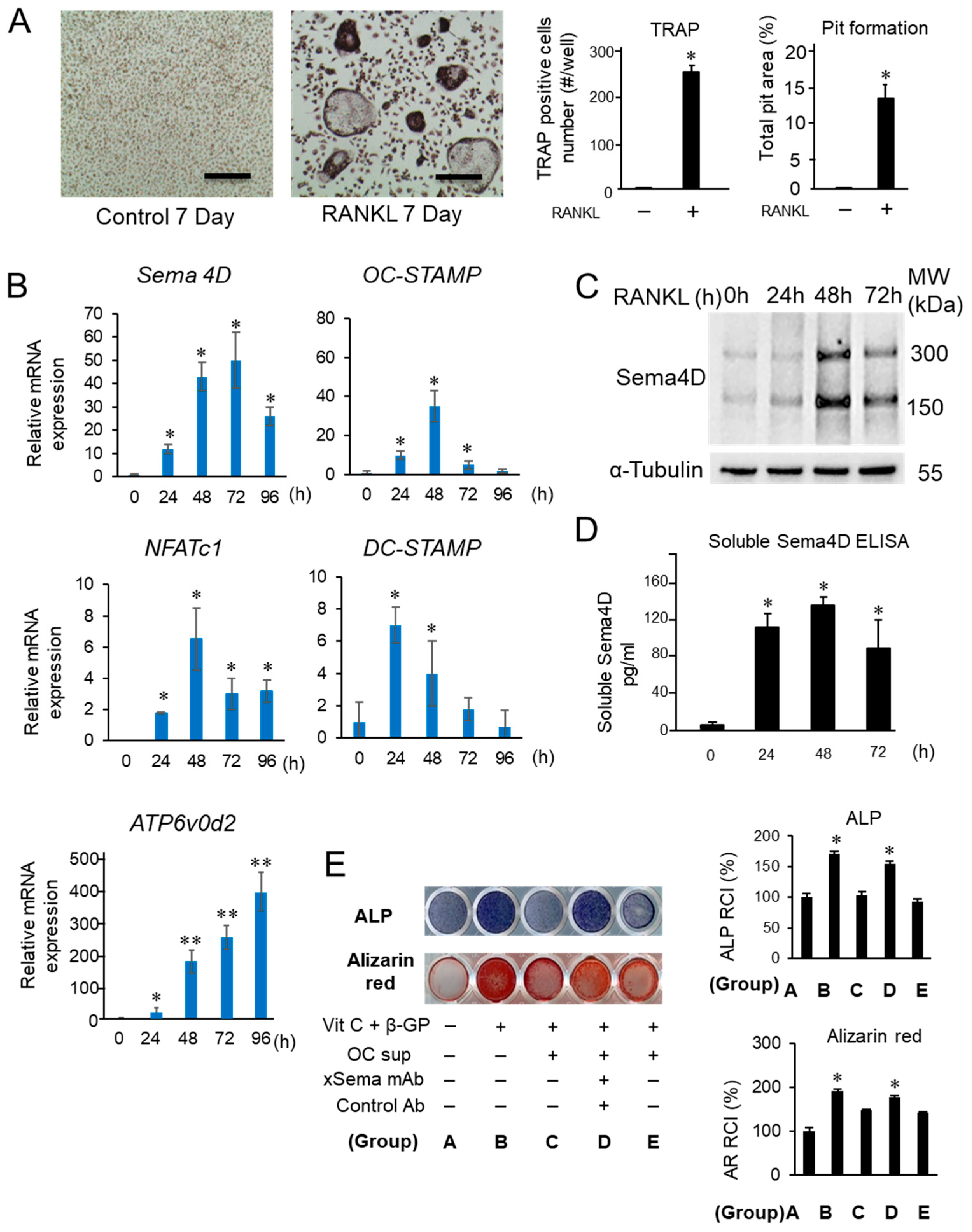
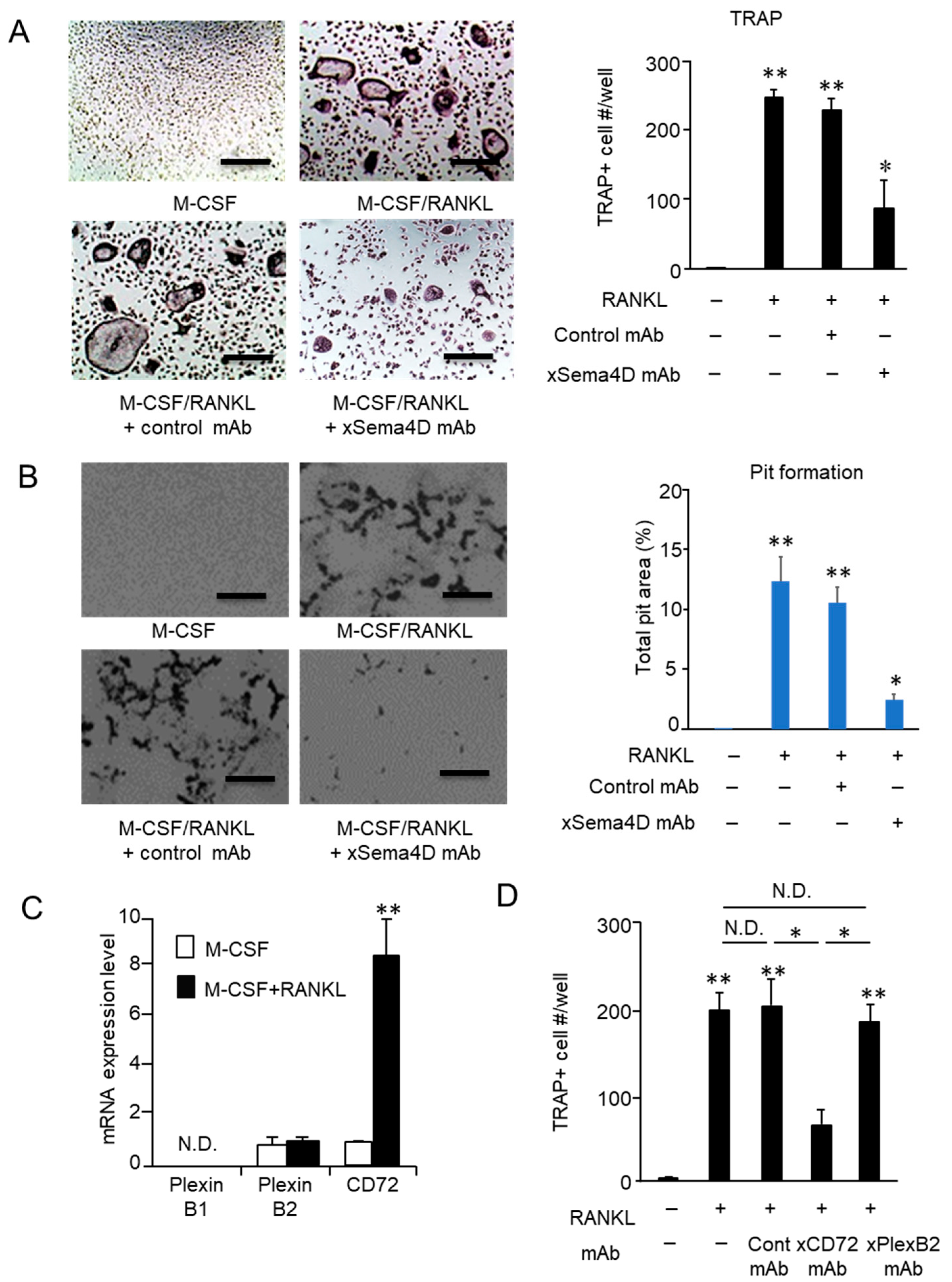
2.4. Sema4D Expressed by Osteoclasts Appeared to Facilitate the Upregulation of Osteoclastogenesis
2.5. Sema4D Upregulates Osteoclastogenesis via Ligation with CD72 Expressed on Osteoclasts
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Bone Marrow Cells Culture and Osteoclast Differentiation Assay
4.3. Osteoblastogenesis Assay
4.4. Real-Time RT-PCR
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Western Blotting
4.7. Ligature-Induced PD Model and Anti-Sema4D Antibody Injection
4.8. Assessment of Alveolar Bone Resorption, Histological Analysis of Periodontal Tissue Section and ELISA of Tissue Homogenate
4.9. Fluorescent Bone Labeling
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eke, P.I.; Borgnakke, W.S.; Genco, R.J. Recent epidemiologic trends in periodontitis in the USA. Periodontol. 2000 2019, 82, 257–267. [Google Scholar] [CrossRef]
- Kim, J.; Amar, S. Periodontal disease and systemic conditions: A bidirectional relationship. Odontology 2006, 94, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, F.Q.; Almeida-Da-Silva, C.L.C.; Huynh, B.; Trinh, A.; Liu, J.; Woodward, J.; Asadi, H.; Ojcius, D.M. Association between periodontal pathogens and systemic disease. Biomed. J. 2019, 42, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Gigante, I.; Colucci, S.; Grano, M. Periodontal disease: Linking the primary inflammation to bone loss. Clin. Dev. Immunol. 2013, 2013, 503754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartold, P.; Cantley, M.; Haynes, D.R. Mechanisms and control of pathologic bone loss in periodontitis. Periodontol. 2000 2010, 53, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Könönen, E.; Gursoy, M.; Gursoy, U. Periodontitis: A Multifaceted Disease of Tooth-Supporting Tissues. J. Clin. Med. 2019, 8, 1135. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, Q.; Chen, Q. The cytokine network involved in the host immune response to periodontitis. Int. J. Oral Sci. 2019, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Matsuyama, T.; Hosokawa, Y.; Makihira, S.; Seki, M.; Karimbux, N.Y.; Goncalves, R.B.; Valverde, P.; Dibart, S.; Li, Y.-P.; et al. B and T Lymphocytes Are the Primary Sources of RANKL in the Bone Resorptive Lesion of Periodontal Disease. Am. J. Pathol. 2006, 169, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Kajiya, M.; Giro, G.; Taubman, M.A.; Han, X.; Mayer, M.P.A.; Kawai, T. Role of periodontal pathogenic bacteria in RANKL-mediated bone destruction in periodontal disease. J. Oral Microbiol. 2010, 2, 5532. [Google Scholar] [CrossRef]
- Alvarez, C.; Monasterio, G.; Cavalla, F.; Córdova, L.A.; Hernández, M.; Heymann, D.; Garlet, G.P.; Sorsa, T.; Pärnänen, P.; Lee, H.-M.; et al. Osteoimmunology of Oral and Maxillofacial Diseases: Translational Applications Based on Biological Mechanisms. Front. Immunol. 2019, 10, 1664. [Google Scholar] [CrossRef] [Green Version]
- Cavalla, F.; Letra, A.; Silva, R.M.; Garlet, G.P. Determinants of Periodontal/Periapical Lesion Stability and Progression. J. Dent. Res. 2021, 100, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Usui, M.; Onizuka, S.; Sato, T.; Kokabu, S.; Ariyoshi, W.; Nakashima, K. Mechanism of alveolar bone destruction in periodontitis—Periodontal bacteria and inflammation. Jpn. Dent. Sci. Rev. 2021, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Inflammatory bone destruction and osteoimmunology. J. Periodontal Res. 2005, 40, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Weivoda, M.M.; Chew, C.K.; Monroe, D.G.; Farr, J.N.; Atkinson, E.J.; Geske, J.R.; Eckhardt, B.; Thicke, B.; Ruan, M.; Tweed, A.J.; et al. Identification of osteo-clast-osteoblast coupling factors in humans reveals links between bone and energy metabolism. Nat. Commun. 2020, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Howard, G.A.; Bottemiller, B.L.; Turner, R.T.; Rader, J.I.; Baylink, D.J. Parathyroid hormone stimulates bone formation and resorption in organ culture: Evidence for a coupling mechanism. Proc. Natl. Acad. Sci. USA 1981, 78, 3204–3208. [Google Scholar] [CrossRef] [Green Version]
- Hathaway-Schrader, J.D.; Novince, C.M. Maintaining homeostatic control of periodontal bone tissue. Periodontol. 2000 2021, 86, 157–187. [Google Scholar] [CrossRef]
- Bougeret, C.; Mansur, I.G.; Dastot, H.; Schmid, M.; Mahouy, G.; Bensussan, A.; Boumsell, L. Increased surface expression of a newly identified 150-kDa dimer early after human T lymphocyte activation. J. Immunol. 1992, 148, 318–323. [Google Scholar]
- Kumanogoh, A.; Kikutani, H. Roles of the Semaphorin Family in Immune Regulation. Adv. Immunol. 2003, 81, 173–198. [Google Scholar] [CrossRef]
- Takamatsu, H.; Okuno, T.; Kumanogoh, A. Regulation of immune cell responses by semaphorins and their receptors. Cell. Mol. Immunol. 2010, 7, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, B.; Ma, Y.; Jin, B. Sema 4D/CD100-plexin B is a multifunctional counter-receptor. Cell. Mol. Immunol. 2013, 10, 97–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumanogoh, A.; Kikutani, H. The CD100–CD72 interaction: A novel mechanism of immune regulation. Trends Immunol. 2001, 22, 670–676. [Google Scholar] [CrossRef]
- Kumanogoh, A.; Kikutani, H. Biological functions and signaling of a transmembrane semaphorin, CD100/Sema4D. Cell. Mol. Life Sci. CMLS 2004, 61, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Kuklina, E. Semaphorin 4D as a guidance molecule in the immune system. Int. Rev. Immunol. 2021, 40, 268–273. [Google Scholar] [CrossRef]
- Elhabazi, A.; Marie-Cardine, A.; Chabbert-de Ponnat, I.; Bensussan, A.; Boumsell, L. Structure and function of the immune semaphorin CD100/SEMA4D. Crit. Rev. Immunol. 2003, 23, 65–81. [Google Scholar] [CrossRef]
- Dickson, B.J. Molecular Mechanisms of Axon Guidance. Science 2002, 298, 1959–1964. [Google Scholar] [CrossRef] [Green Version]
- Basile, J.R.; Castilho, R.M.; Williams, V.P.; Gutkind, J.S. Semaphorin 4D provides a link between axon guidance processes and tumor-induced angiogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 9017–9022. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, L.; Liu, W.X.; Wang, K. VEGF and SEMA4D have synergistic effects on the promotion of angio-genesis in epithelial ovarian cancer. Cell. Mol. Biol. Lett. 2018, 23, 2. [Google Scholar] [CrossRef] [Green Version]
- Dacquin, R.; Domenget, C.; Kumanogoh, A.; Kikutani, H.; Jurdic, P.; Machuca-Gayet, I. Control of Bone Resorption by Semaphorin 4D Is Dependent on Ovarian Function. PLoS ONE 2011, 6, e26627. [Google Scholar] [CrossRef] [Green Version]
- Negishi-Koga, T.; Shinohara, M.; Komatsu, N.; Bito, H.; Kodama, T.; Friedel, R.H.; Takayanagi, H. Suppression of bone formation by osteoclastic expression of semaphorin 4D. Nat. Med. 2011, 17, 1473–1480. [Google Scholar] [CrossRef]
- Liu, X.L.; Song, J.; Liu, K.J.; Wang, W.P.; Xu, C.; Zhang, Y.Z.; Liu, Y. Role of inhibition of osteogenesis function by Se-ma4D/Plexin-B1 signaling pathway in skeletal fluorosis in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2015, 35, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Veyisoğlu, G.; Savran, L.; Narin, F.; Yılmaz, H.E.; Avşar, C.; Sağlam, M. Gingival crevicular fluid semaphorin 4D and peptidylarginine deiminase-2 levels in periodontal health and disease. J. Periodontol. 2019, 90, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, E.; Xu, Y.; Wang, W.; Zhang, T.; Xiao, L.; Chen, R.; Lin, Y.; Chen, D.; Lin, L.; et al. Serum Sema4D levels are associated with lumbar spine bone mineral density and bone turnover markers in patients with postmenopausal osteoporosis. Int. J. Clin. Exp. Med. 2015, 8, 16352–16357. [Google Scholar] [PubMed]
- Yoshida, Y.; Ogata, A.; Kang, S.; Ebina, K.; Shi, K.; Nojima, S.; Kimura, T.; Ito, D.; Morimoto, K.; Nishide, M.; et al. Semaphorin 4D Contributes to Rheumatoid Ar-thritis by Inducing Inflammatory Cytokine Production: Pathogenic and Therapeutic Implications. Arthritis Rheumatol. 2015, 67, 1481–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleki, K.T.; Cornillet, M.; Björkström, N.K. Soluble SEMA4D/CD100: A novel immunoregulator in infectious and inflammatory diseases. Clin. Immunol. 2016, 163, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, T.; Mitoma, H.; Harashima, S.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-alpha: Structure, func-tion and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shindo, S.; Savitri, I.J.; Ishii, T.; Ikeda, A.; Pierrelus, R.; Heidari, A.; Okubo, K.; Nakamura, S.; Kandalam, U.; Rawas-Qalaji, M.; et al. Dual-Function Semaphorin 4D Released by Platelets: Suppression of Osteoblastogenesis and Promotion of Osteoclastogenesis. Int. J. Mol. Sci. 2022, 23, 2938. [Google Scholar] [CrossRef]
- Movila, A.; Mawardi, H.; Nishimura, K.; Kiyama, T.; Egashira, K.; Kim, J.Y.; Villa, A.; Sasaki, H.; Woo, S.-B.; Kawaiah, T.; et al. Possible pathogenic engagement of soluble Semaphorin 4D produced by gammadeltaT cells in medication-related osteonecrosis of the jaw (MRONJ). Biochem. Biophys. Res. Commun. 2016, 480, 42–47. [Google Scholar] [CrossRef]
- Zhu, L.; Bergmeier, W.; Wu, J.; Jiang, H.; Stalker, T.J.; Cieslak, M.; Fan, R.; Boumsell, L.; Kumanogoh, A.; Kikutani, H.; et al. Regulated surface expression and shedding support a dual role for semaphorin 4D in platelet responses to vascular injury. Proc. Natl. Acad. Sci. USA 2007, 104, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Bi, L.; Yu, X.; Kawai, T.; Taubman, M.A.; Shen, B.; Han, X. Porphyromonas gingivalis Exacerbates Ligature-Induced, RANKL-Dependent Alveolar Bone Resorption via Differential Regulation of Toll-like Receptor 2 (TLR2) and TLR4. Infect. Immun. 2014, 82, 4127–4134. [Google Scholar] [CrossRef] [Green Version]
- Graves, D.T.; Fine, D.; Teng, Y.-T.A.; Van Dyke, T.E.; Hajishengallis, G. The use of rodent models to investigate host-bacteria interactions related to periodontal diseases. J. Clin. Periodontol. 2008, 35, 89–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Deng, Z.; Seneviratne, C.J.; Cheung, G.S.; Jin, L.; Zhao, B.; Zhang, C. Enterococcus faecalis promotes osteoclasto-genesis and semaphorin 4D expression. Innate Immun. 2015, 21, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Takada, H.; Ibaragi, S.; Eguchi, T.; Okui, T.; Obata, K.; Masui, M.; Morisawa, A.; Takabatake, K.; Kawai, H.; Yoshioka, N.; et al. Semaphorin 4D promotes bone invasion in head and neck squamous cell carcinoma. Int. J. Oncol. 2017, 51, 625–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufeld, G.; Lange, T.; Varshavsky, A.; Kessler, O. Semaphorin Signaling in Vascular and Tumor Biology. Adv. Exp. Med. Biol. 2007, 600, 118–131. [Google Scholar] [CrossRef]
- Takegahara, N.; Kumanogoh, A.; Kikutani, H. Semaphorins: A new class of immunoregulatory molecules. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Li, J.; Gao, Q.; Ye, F. The role of Sema4D/CD100 as a therapeutic target for tumor microenvironments and for autoimmune, neuroimmune and bone diseases. Expert Opin. Ther. Targets 2016, 20, 885–901. [Google Scholar] [CrossRef]
- Filipowska, J.; Tomaszewski, K.; Niedźwiedzki, Ł.; Walocha, J.A.; Niedźwiedzki, T. The role of vasculature in bone development, regeneration and proper systemic functioning. Angiogenesis 2017, 20, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Sousa, S.; Clézardin, P. Bone-Targeted Therapies in Cancer-Induced Bone Disease. Calcif. Tissue Res. 2018, 102, 227–250. [Google Scholar] [CrossRef]
- Deeks, E.D. Denosumab: A Review in Postmenopausal Osteoporosis. Drugs Aging 2017, 35, 163–173. [Google Scholar] [CrossRef]
- Anagnostis, P.; Paschou, S.A.; Mintziori, G.; Ceausu, I.; Depypere, H.; Lambrinoudaki, I.; Mueck, A.; Pérez-López, F.R.; Rees, M.; Senturk, L.M.; et al. Drug holidays from bisphosphonates and denosumab in postmenopausal osteoporosis: EMAS position statement. Maturitas 2017, 101, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Yin, R.V.; Hirpara, H.; Lee, N.C.; Lee, A.; Llanos, S.; Phung, O.J. Increased risk for atypical fractures associated with bisphosphonate use. Fam. Pract. 2015, 32, 276–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A.; Morrison, A.; Hanley, D.A.; Felsenberg, D.; McCauley, L.K.; O’Ryan, F.; Reid, I.R.; Ruggiero, S.L.; Taguchi, A.; Tetradis, S.; et al. Diagnosis and Management of Osteonecrosis of the Jaw: A Systematic Review and International Consensus. J. Bone Miner. Res. 2015, 30, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, S.L.; Dodson, T.B.; Fantasia, J.; Goodday, R.; Aghaloo, T.; Mehrotra, B.; O’Ryan, F. American Association of Oral and Maxillofacial Surgeons Position Paper on Medication-Related Osteonecrosis of the Jaw—2014 Update. J. Oral Maxillofac. Surg. 2014, 72, 1938–1956. [Google Scholar] [CrossRef] [PubMed]
- Ishida, I.; Kumanogoh, A.; Suzuki, K.; Akahani, S.; Noda, K.; Kikutani, H. Involvement of CD100, a lymphocyte semaphorin, in the activation of the human immune system via CD72: Implications for the regulation of immune and inflammatory responses. Int. Immunol. 2003, 15, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Kumanogoh, A.; Kikutani, H. Semaphorins and their receptors in immune cell interactions. Nat. Immunol. 2008, 9, 17–23. [Google Scholar] [CrossRef]
- Van de Velde, H.; von Hoegen, I.; Luo, W.; Parnes, J.R.; Thielemans, K. The B-cell surface protein CD72/Lyb-2 is the ligand for CD5. Nature 1991, 351, 662–665. [Google Scholar] [CrossRef]
- Brown, M.H.; Lacey, E. A ligand for CD5 is CD5. J. Immunol. 2010, 185, 6068–6074. [Google Scholar] [CrossRef]
- Kumanogoh, A.; Watanabe, C.; Lee, I.; Wang, X.; Shi, W.; Araki, H.; Hirata, H.; Iwahori, K.; Uchida, J.; Yasui, T.; et al. Identification of CD72 as a lymphocyte receptor for the class IV semaphorin CD100: A novel mechanism for regulating B cell signaling. Immunity 2000, 13, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Tsubata, T. CD72 is a Negative Regulator of B Cell Responses to Nuclear Lupus Self-antigens and Development of Systemic Lupus erythematosus. Immune Netw. 2019, 19, e1. [Google Scholar] [CrossRef]
- Ishii, T.; Ruiz-Torruella, M.; Ikeda, A.; Shindo, S.; Movila, A.; Mawardi, H.; Albassam, A.; Kayal, R.A.; Al-Dharrab, A.A.; Egashira, K.; et al. OC-STAMP promotes osteoclast fu-sion for pathogenic bone resorption in periodontitis via up-regulation of permissive fusogen CD9. FASEB J. 2018, 32, 4016–4030. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Movila, A.; Kataoka, S.; Wisitrasameewong, W.; Ruiz Torruella, M.; Murakoshi, M.; Albassam, A.; Kayal, R.A.; Al-Dharrab, A.A.; Egashira, K.; et al. Proinflammatory M1 Macrophages Inhibit RANKL-Induced Osteoclastogenesis. Infect. Immun. 2016, 84, 2802–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisitrasameewong, W.; Kajiya, M.; Movila, A.; Rittling, S.; Ishii, T.; Suzuki, M.; Matsuda, S.; Mazda, Y.; Torruella, M.; Azuma, M.; et al. DC-STAMP Is an Osteoclast Fusogen Engaged in Periodontal Bone Resorption. J. Dent. Res. 2017, 96, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Paster, B.J.; Komatsuzawa, H.; Ernst, C.W.; Goncalves, R.B.; Sasaki, H.; Ouhara, K.; Stashenko, P.P.; Sugai, M.; Taubman, M.A. Cross-reactive adaptive im-mune response to oral commensal bacteria results in an induction of receptor activator of nuclear factor-kappaB ligand (RANKL)-dependent periodontal bone resorption in a mouse model. Oral Microbiol. Immunol. 2007, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Movila, A.; Suzuki, M.; Kajiya, M.; Wisitrasameewong, W.; Kayal, R.; Hirshfeld, J.; Aldharrab, A.; Savitri, I.J.; Mira, A.; et al. A novel method of sampling gingival crevicular fluid from a mouse model of periodontitis. J. Immunol. Methods 2016, 438, 21–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishii, T.; Ruiz-Torruella, M.; Yamamoto, K.; Yamaguchi, T.; Heidari, A.; Pierrelus, R.; Leon, E.; Shindo, S.; Rawas-Qalaji, M.; Pastore, M.R.; et al. Locally Secreted Semaphorin 4D Is Engaged in Both Pathogenic Bone Resorption and Retarded Bone Regeneration in a Ligature-Induced Mouse Model of Periodontitis. Int. J. Mol. Sci. 2022, 23, 5630. https://doi.org/10.3390/ijms23105630
Ishii T, Ruiz-Torruella M, Yamamoto K, Yamaguchi T, Heidari A, Pierrelus R, Leon E, Shindo S, Rawas-Qalaji M, Pastore MR, et al. Locally Secreted Semaphorin 4D Is Engaged in Both Pathogenic Bone Resorption and Retarded Bone Regeneration in a Ligature-Induced Mouse Model of Periodontitis. International Journal of Molecular Sciences. 2022; 23(10):5630. https://doi.org/10.3390/ijms23105630
Chicago/Turabian StyleIshii, Takenobu, Montserrat Ruiz-Torruella, Kenta Yamamoto, Tsuguno Yamaguchi, Alireza Heidari, Roodelyne Pierrelus, Elizabeth Leon, Satoru Shindo, Mohamad Rawas-Qalaji, Maria Rita Pastore, and et al. 2022. "Locally Secreted Semaphorin 4D Is Engaged in Both Pathogenic Bone Resorption and Retarded Bone Regeneration in a Ligature-Induced Mouse Model of Periodontitis" International Journal of Molecular Sciences 23, no. 10: 5630. https://doi.org/10.3390/ijms23105630
APA StyleIshii, T., Ruiz-Torruella, M., Yamamoto, K., Yamaguchi, T., Heidari, A., Pierrelus, R., Leon, E., Shindo, S., Rawas-Qalaji, M., Pastore, M. R., Ikeda, A., Nakamura, S., Mawardi, H., Kandalam, U., Hardigan, P., Witek, L., Coelho, P. G., & Kawai, T. (2022). Locally Secreted Semaphorin 4D Is Engaged in Both Pathogenic Bone Resorption and Retarded Bone Regeneration in a Ligature-Induced Mouse Model of Periodontitis. International Journal of Molecular Sciences, 23(10), 5630. https://doi.org/10.3390/ijms23105630