Abstract
Arsenic (As), distributed widely in the natural environment, is a toxic substance which can severely impair the normal functions in living cells. Research on the genetic determinants conferring functions in arsenic resistance and metabolism is of great importance for remediating arsenic-contaminated environments. Many organisms, including bacteria, have developed various strategies to tolerate arsenic, by either detoxifying this harmful element or utilizing it for energy generation. More and more new arsenic resistance (ars) determinants have been identified to be conferring resistance to diverse arsenic compounds and encoded in ars operons. There is a hazard in mobilizing arsenic during gold-mining activities due to gold- and arsenic-bearing minerals coexisting. In this study, we isolated 8 gold enrichment strains from the Zijin gold and copper mine (Longyan, Fujian Province, China) wastewater treatment site soil, at an altitude of 192 m. We identified two Brevundimonas nasdae strains, Au-Bre29 and Au-Bre30, among these eight strains, having a high minimum inhibitory concentration (MIC) for As(III). These two strains contained the same ars operons but displayed differences regarding secretion of extra-polymeric substances (EPS) upon arsenite (As(III)) stress. B. nasdae Au-Bre29 contained one extra plasmid but without harboring any additional ars genes compared to B. nasdae Au-Bre30. We optimized the growth conditions for strains Au-Bre29 and Au-Bre30. Au-Bre30 was able to tolerate both a lower pH and slightly higher concentrations of NaCl. We also identified folE, a folate synthesis gene, in the ars operon of these two strains. In most organisms, folate synthesis begins with a FolE (GTP-Cyclohydrolase I)-type enzyme, and the corresponding gene is typically designated folE (in bacteria) or gch1 (in mammals). Heterologous expression of folE, cloned from B. nasdae Au-Bre30, in the arsenic-hypersensitive strain Escherichia coli AW3110, conferred resistance to As(III), arsenate (As(V)), trivalent roxarsone (Rox(III)), pentavalent roxarsone (Rox(V)), trivalent antimonite (Sb(III)), and pentavalent antimonate (Sb(V)), indicating that folate biosynthesis is a target of arsenite toxicity and increased production of folate confers increased resistance to oxyanions. Genes encoding Acr3 and ArsH were shown to confer resistance to As(III), Rox(III), Sb(III), and Sb(V), and ArsH also conferred resistance to As(V). Acr3 did not confer resistance to As(V) and Rox(V), while ArsH did not confer resistance to Rox(V).
1. Introduction
Arsenic (As) is a metalloid occurring all over the world, and ranks at the top of the US Priority List of Hazardous Substances (http://www.atsdr.cdc.gov/SPL/index.html, accessed on 15 December 2021). The concentrations of arsenic by weight in the crustal crust and sea water are around 2100 and 2.3 μg/kg, respectively (http://www.webelements.com/arsenic/geology.html, accessed on 15 December 2021). Arsenic can be accumulated and transferred via natural and anthropogenic activities, which finally enter the biosphere, in both terrestrial and oceanic environments, ending up as a toxin to organisms. It has been estimated that around 150 million people in the world are exposed to higher than 50 μg/L of As in their drinking water during daily life [1]. Exposure to arsenic leads to numerous diseases, such as diabetes, peripheral vascular disease, and even cancer [2,3].
In the environment, inorganic arsenic exists in various chemical forms, such as arsenate (As(V)), arsenite (As(III)), elemental arsenic, and arsine. Due to differences in Eh and pH, As(V) and As(III) are the predominant forms in the oxic and in the reducing environment, respectively [4]. As(III) is more toxic than As(V) owing to the high affinity and capability to form metallic compounds, and As(III) was present in significant amounts globally much earlier than As(V). Organic arsenic compounds also exist in various forms, such as pentavalent roxarsone (Rox(V)), methylarsenate (MAs(V)), and the trivalent roxarsone (Rox(III)) and methylarsenite (MAs(III)) [5]. Antimony (Sb) and As are group VA elements on the periodic table, and Sb shares some similar chemical and toxicological properties with As, which have been extensively studied [6]. Sb exists in four oxidation states (+V, +III, 0, and −III), of which pentavalent antimonate (Sb(V)) and trivalent antimonite (Sb(III)) are the prevalent forms in the environment [7]. Microbial communities have been shown to play an important role in the arsenic and antimony global biocycles [8,9]. As(III) efflux and methylation are responsible for the majority of As(III) tolerance and detoxification pathways in most organisms [10,11]. More and more new arsenic resistance (ars) genes have been found to confer resistance to arsenic and various organic arsenic compounds and are encoded in ars operons [12,13,14,15,16,17,18]. Acr3 and ArsB are the most widespread ars operon-encoded gene products, which were shown to pump As(III) from the cytosol across the cytoplasmic membrane into the periplasm or extracellular medium [10]. ArsG, ArsJ, ArsK, ArsP, ArsQ and ArsW have been shown to be efflux transporters encoded on ars operons in various organisms, with each mediating transport of different types of organic arsenicals [12,13,14,19,20,21]. ArsC, an As(V) reductase, has been shown to quickly reduce intracellular As(V) to As(III) [22]. ArsE, ArsF, ArsM, ArsH, ArsI, ArsN, ArsO, ArsU, ArsV and ArsL were shown to confer resistance to organic arsenical [12,13,14,15,16,17,18,23,24].
In this study, Brevundimonas nasdae strains Au-Bre29 and Au-Bre30 were isolated from Zijin gold- and copper-mining soil, with high multiple heavy-metal(loid)s resistance, but behaving differently regarding secretion of extra-polymeric substances (EPS) upon As(III) stress. We further characterized different genes conferring resistance to various arsenic and antimony species and compounds.
2. Results
2.1. The Soil Sample from the Zijin Gold and Copper Mine Contained Heavy Metals and Metalloids at Very High Concentrations
The heavy-metal(loid)s concentrations in the sampling soil of the Zijin gold and copper mine were determined, respectively (Table 1). The concentrations of As, Cd, Cu and Pb in the sampling site were significantly higher than the third level of the environmental quality standard for soils of China (GB 15618-1995).

Table 1.
Heavy-metal concentrations in the sampling soil. Data are presented as means ± standard deviations (n = 3).
2.2. Eight Highly Arsenite-Resistant Strains Were Isolated and Identified
Eight strains, named Au-Aci5, Au-Bre4, Au-Bre29, Au-Bre30, Au-Lys6, Au-Mic3, Au-Pse14 and Au-Pse15, respectively, were isolated from the Au(III) enrichment on R2A plates containing 160 μM of Au(III). Three of the eight isolates were shown to belong to the genus of Brevundimonas after blasting the 16S ribosomal RNA (rRNA) sequence to the database of the National Center for Biotechnology Information (NCBI). Au-Aci5 showed 100% similarity to Acinetobacter seifertii CMCC(B)25090, Au-Bre4 had 99.92% similarity to Brevundimonas vesicularis DCB19, Au-Bre29 and Au-Bre30 had 99.85% and 99.78% similarity to Brevundimonas nasdae DSS_C4, Au-Lys6 had 99.93% similarity to Lysinibacillus fusiformis LSP22, Au-Mci3 showed 100% similarity to Microbacterium binotii CIP 102116, and Au-Pse14 and Au-Pse15 had 99.93% similarity to Pseudomonas frederiksbergensis JS1015, respectively. The 16S rRNA sequences of the eight isolates were deposited to the NCBI database under accession numbers: OK178958, OK178960, OK182900, OK182930, OK178961, OK178958, OK271131 and OK560628, respectively (Table 2). Based on the 16S rRNA alignment results and the phenotype of the eight isolates, strains Au-Bre29 and Au-Bre30 with high similarity to Brevundimonas nasdae were selected for the following study.
Table 2.
Eight Au enrichment isolates from the Zijin gold and copper mine.
2.3. High Arsenic Resistance and Heavy-Metal Resistance Were Determined in B. nasdae Strains Au-Bre29 and Au-Bre30
After incubation on R2A plates containing various concentrations of Au(III), Cu(II), Zn(II), Cd(II) and As(III) at 28 °C for 3 days, the minimum inhibitory concentration (MIC) of strains Au-Bre29 and Au-Bre30 to heavy metal(loid)s was determined. Different isolates displayed differing abilities to resist different metal(loid)s, but all the strains conferred a high resistance to Au(III) (>100 μM) (Table 3). Furthermore, strain Au-Bre29 was found to be more resistant to Cu(II) and less resistant to As(III) compared to strain Au-Bre30. The EPS production of strain Au-Bre30 on R2A plates containing As(III) was significantly higher than in strain Au-Bre29, indicating that strains Au-Bre30 and Au-Bre29 might have a partially different response to As(III) exposure.
Table 3.
MICs of strains Au-Bre29 and Au-Bre30 for selected heavy metal(loid)s.
2.4. The Growth Conditions for B. nasdae Strains Au-Bre29 and Au-Bre30 could Be Optimized
The growth of bacterial strains depends on growth parameters including the behavior of strains Au-Bre29 and Au-Bre30, and a set of experiments were conducted. As shown in Figure 1, the best medium for growing strains Au-Bre29 and Au-Bre30 is TY medium. Strain Au-Bre29 was able to grow in R2A medium with pH ranging from 6 to 11, and strain Au-Bre30 was able to tolerate a pH ranging from 5 to 11. Both strains were able to grow in R2A medium with additional NaCl concentrations ranging from 0 to 3%. All these results indicated that there was little influence of these parameters on the growth patterns of both B. nasdae strains, Au-Bre29 and Au-Bre30.
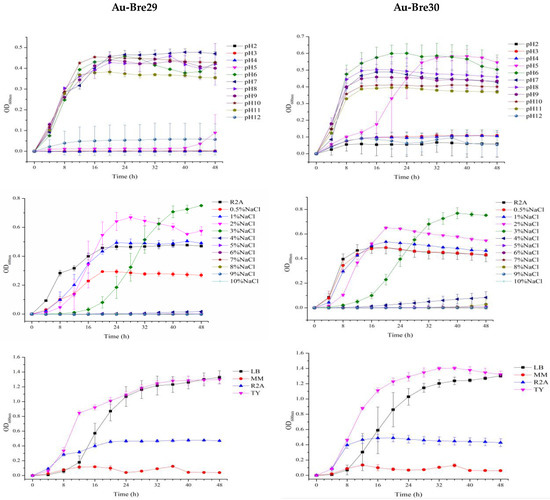
Figure 1.
Optimized growth patterns of B. nasdae strains Au-Bre29 and Au-Bre30. Au-Bre29 and Au-Bre30 were cultured individually in R2A medium with different pH levels (2–12, additional NaCl = 0%), different additional NaCl concentrations (0–10%, pH = 7.0), and different media (MM, R2A, TY and LB, pH = 7.0). Performed in the Bioscreen C microbiology reader at 28 °C for 48 h, and the optical densities at 600nm (OD600nm) were automatically measured at 4 h intervals. The growth curves display the results of three replicates, and error bars represent the standard error of the average values.
2.5. Genome Analysis of B. nasdae Strains Au-Bre29 and Au-Bre30
The complete genome was sequenced to be able to compare B. nasdae strains Au-Bre29 and Au-Bre30 with a focus on heavy-metal resistance, especially As(III) resistance determinants. The circular maps of the complete genome of both Au-Bre29 and Au-Bre30 are shown in Figure S1. Compared to strain Au-Bre30, strain Au-Bre29 contained one extra plasmid. Genome size (Kb), genes existing on the forward strand and reverse strand, repetitive sequence, annotated tRNA (blue) and rRNA (purple), GC content, and GC skew are also presented in Figure S1. As show in Table S1, the G+C content of Au-Bre29 and Au-Bre30 is 65.69% and 65.8%, respectively. Strain Au-Bre29 was shown to have 3665 coding sequence (CDS) and 53 tRNAs, while strain Au-Bre30 was shown to have 3577 CDS and 53 tRNAs. A total of 3731 genes were found on the genome of strain Au-Bre29, and 3618 genes were assigned with function prediction, 1772 genes were predicted to encode functions involved in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, and 531 genes were assigned to signal peptides. As in strain Au-Bre29, strain Au-Bre30 was shown to contain 3643 genes, with 3531, 3474, 1751 and 523 genes associated with protein coding sequence, function prediction, KEGG pathway, and signal peptides, respectively. Furthermore, the presence of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) loci has been shown to increase the genomic stability of strains, allowing them to adapt to various environments [25].
As shown in Figure S2, there are many hypothetical proteins encoded on the plasmid of Au-Bre29. Other genes encoded conjugative transfer system proteins, such as TraC, TraE, TraF, TraG and TraH. There was no arsenic resistance gene on the plasmid of Au-Bre29. As Figure S3 shows, the Mauve alignment showed high synteny and similarity among the genomes of B. nasdae strains Au-Bre29 and Au-Bre30.
2.6. ars Operons in B. nasdae Strains Au-Bre29 and Au-Bre30 Are Highly Similar
Three ars operons were identified on each of the chromosomes of strains Au-Bre29 and Au-Bre30: arsenic resistance protein1–arsR1–arsC1–acr31–arsN1–arsH1 (ars1), arsC2–acr32–arsR2–arsH2–folE–acyltransferase1–sugar transferase–mfs (ars2), and IS5 family transposase–acyltransferase2–copA–copB–arsenic resistance protein2–arsN2–arsH3–trbC (ars3) (Figure 2A). All of the ars operons were only found on the chromosome and not on the plasmid. Interestingly, a gene annotated as folE was located near arsH in the ars operon. Interestingly, the expression of folE was upregulated in strain Paenibacillus taichungensis NC1 exposed to 0.2 and 2 mM of As(III), indicating that FolE is predicted to have a role in conferring arsenic resistance in both strains [26]. After comparison of the genome sequences of the two strains, no major difference was found expect for the presence of the copA and copB genes adjacent to one of the ars operons (Figure 2B).
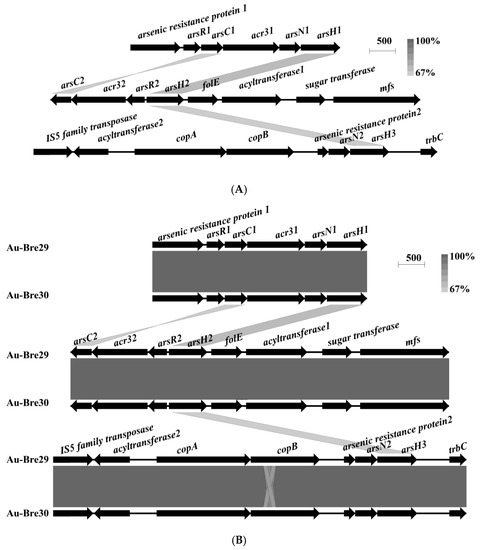
Figure 2.
ars operons in B. nasdae strain Au-Bre30 (A), and the alignment between the ars operons present on the chromosome of B. nasdae strains Au-Bre29 and Au-Bre30 (B). Physical maps were generated by using Easyfig.
2.7. Heterologous Expression of folE Confers Resistance to As(III), As(V), Rox(III), Rox(V), Sb(III), and Sb(V)
E. coli AW3110, the arsenic-hypersensitive strain with an arsRBC deletion, contained the control pTOPO plasmid or the pTOPO plasmid carrying folE to determine the function of FolE. E. coli AW3110 carrying plasmid pTOPO was able to grow at concentrations of As(III), Rox(III) and Sb(III) up to 0.15 mM, 6 μM, and 0.4 mM, respectively. E. coli AW3110 bearing plasmid pTOPO-FolE was able to grow when exposed to concentrations higher than 0.45 mM, 10 μM, and 0.6 mM of As(III), Rox(III), and Sb(III), respectively. E. coli AW3110 carrying plasmid pTOPO was able to tolerate As(V), Rox(V), and Sb(V) up to 1, 3, and 5 mM, respectively. E. coli AW3110 bearing plasmid pTOPO-FolE was able to grow when exposed to concentrations higher than 4, 5 and 10 mM of As(V), Rox(V) and Sb(V), respectively. Cells expressing folE were not only more resistant to organic and inorganic arsenic but also to trivalent and pentavalent antimony (Figure 3).
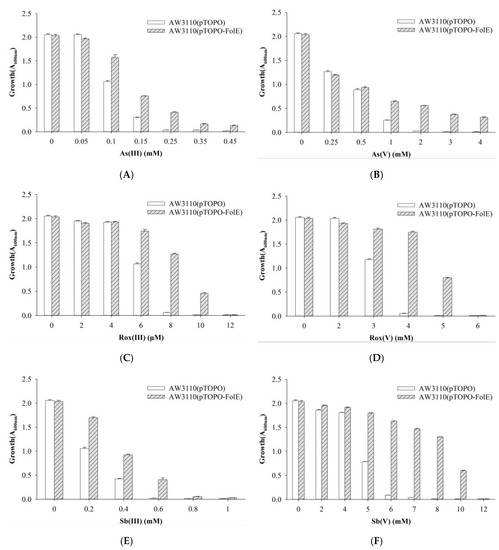
Figure 3.
Growth of E. coli AW3110 bearing pTOPO and pTOPO-FolE with the indicated concentrations of As(III) (A), As(V) (B), Rox(III) (C), Rox(V) (D), Sb(III) (E), and Sb(V) (F). The A600 nm was measured after 24 h of growth at 37 °C and 220 rpm. The graphs show the results of three replicates, and error bars represent the standard error of the average values.
To investigate the function of the acr3- and arsH-encoded gene products also present in the ars2 operon, acr3 and arsH from strain Au-Bre30 were cloned into the pTOPO plasmid. The arsenic-sensitive strain E. coli AW3110 carrying plasmid pTOPO-Acr3 was able to grow in 1.2 mM, 8 μM, 0.8 mM, and 10 mM of As(III), Rox(III), Sb(III), and Sb(V), respectively, while the strain containing the empty pTOPO did not grow under the same conditions. However, E. coli AW3110 carrying plasmid pTOPO-Acr3 or pTOPO did not display significant differences when exposed to As(V) and Rox(V). Expression of acr3 conferred very significant resistance to As(III), Sb(III), and Sb(V), conferred a small increase in resistance to Rox(III), but did not confer resistance to As(V) and Rox(V) (Figure 4). E. coli AW3110 carrying plasmid pTOPO-ArsH was able to grow in 0.45 mM, 8 μM, 3 mM, 2 mM and 7 mM of As(III), Rox(III), Sb(III), As(V) and Sb(V), respectively, while E. coli AW3110 containing pTOPO did not grow under the same conditions. However, E. coli AW3110 carrying plasmid pTOPO-ArsH and pTOPO showed no difference when exposed to As(V). Furthermore, we were able to show that expression of arsH conferred very significant resistance to As(III) and Sb(III), conferred significant resistance to Rox(III), As(V) and Sb(V), but did not confer resistance to Rox(V) (Figure 5).
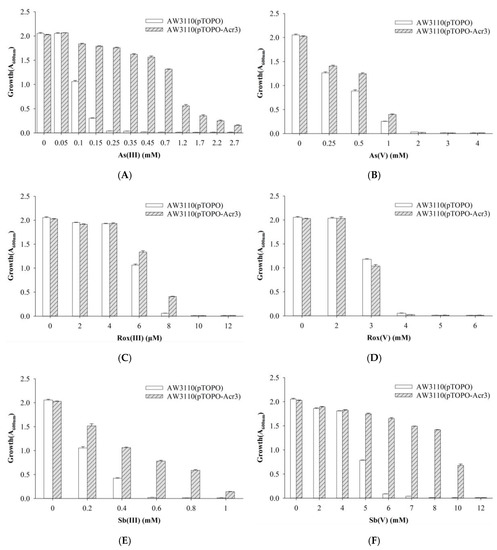
Figure 4.
Growth of E. coli AW3110 bearing pTOPO and pTOPO-Acr3 with the indicated concentrations of As(III) (A), As(V) (B), Rox(III) (C), Rox(V) (D), Sb(III) (E), and Sb(V) (F). The A600 nm was measured after 24 h of growth at 37 °C and 220 rpm. The graphs show the results of three replicates, and error bars represent the standard error of the average values.
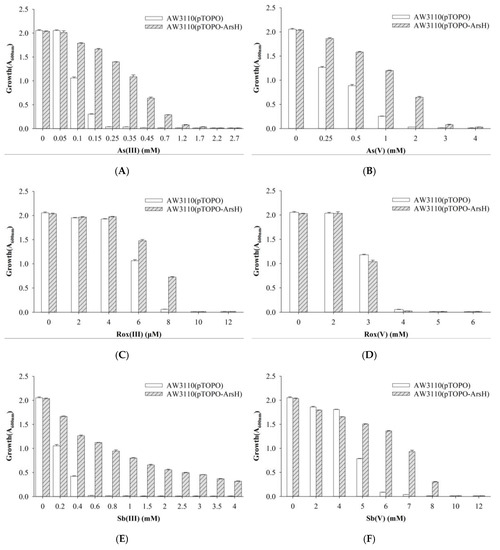
Figure 5.
Growth of E. coli AW3110 bearing pTOPO and pTOPO-ArsH with the indicated concentrations of As(III) (A), As(V) (B), Rox(III) (C), Rox(V) (D), Sb(III) (E), and Sb(V) (F). The A600 nm was measured after 24 h of growth at 37 °C and 220 rpm. The graphs show the results of three replicates, and error bars represent the standard error of the average values.
3. Discussion
Mine soil with extremely high concentrations of multiple heavy metals has been treated as one of the hot spots to study how organisms adapt and survive under these hazardous conditions. The Zijin gold and copper mine is the biggest gold mine in China, with a long history of mining indicating high levels of long-term heavy-metal contamination. In the present study, eight strains were isolated via Au-enrichment isolation, with three of the eight strains belonging to the genus Brevundimonas, with two of the three strains displaying high similarity to Brevundimonas nasdae but having a distinct phenotype on R2A plates, such as different levels of EPS secretion. This indicated the presence of genomic differences between B. nasdae strains Au-Bre29 and Au-Bre30 in their adaptation to heavy-metal-contaminated environments. Complete genome sequencing indicated that B. nasdae strains Au-Bre29 and Au-Bre30 both contain 3 ars operons conferring high arsenic resistance. These three ars operons, ars1, ars2 and ars3, with the ars3 operon in the vicinity of copA and copB, are located on conjugative transposons, and the ars2 operon contains a gene named folE adjacent to other genes with putative arsenic-related functions. Furthermore, these two strains show no difference in ars operons, as only one extra plasmid without any ars genes was found in B. nasdae Au-Bre29. Cloning of folE into an expression vector for heterologous expression in arsenic-hypersensitive strain AW3110 conferred resistance to As(III), As(V), Rox(III), Rox(V), Sb(III), and Sb(V). These results demonstrate that folE, encoding GTP-Cyclohydrolase I, is a novel arsenic resistance gene, that confers resistance to both trivalent and prevalent organic and inorganic arsenic.
Arsenic and antimony from both natural and anthropogenic sources contaminate the environment. As an element, arsenic and antimony cannot be destroyed and will always be present in the environment, but bio-transformations have been able to modify their physical location and chemical speciation. In T24 cells, which were derived from a human urinary bladder carcinoma lacking the ability of As(III) methylation, the relative toxicities were determined to be MAs(III) > DMAs(III) ≈ As(III) >> AsV > DMAs(V) > MAs(V) > Rox(V) [27]. The general order of toxicity for Sb species is presented as: Sb(III) > Sb(V) > organoantimonials [28]. MAs(III) and Rox(III) are likely oxidized by ArsH, resulting in reduced toxicity in E. coli AW3110 expressing arsH (Figure 5).
The arsH gene had previously been cloned from a rhizosphere bacterium, and its gene product, ArsH, was shown to be an NADPH-FMN-dependent oxidoreductase that oxidizes highly toxic MAs(III) to relatively nontoxic MAs(V) [18]. ArsH relieved the toxicity of As species by mediating the reduction of ROS produced in vivo upon exposure to the oxyanion, e.g., by generating FMNH2 to fuel ROS-quenching activities [29]. A recent study found that ArsH was also significantly upregulated in the oxidation process of Sb(III) [30], suggesting the possibility of its participation in the oxidation of Sb(III) in Klebsiella aerogenes X [31]. Pseudomonas putida arsH expressed in E. coli conferred resistance to MAs(III), Rox(III), and phenylarsenite (PhAs(III)) [25]. Our results show that arsH not only conferred resistance to Sb(III) and Rox(III), but also As(III), As(V) and Sb(V), strongly suggesting it not only oxidizes MAs(III) and Rox(III) to MAs(V) and Rox(V), respectively, but also protects against ROS (Figure 5).
GTP-Cyclohydrolase I (GTPCHI, EC 3.5.4.16) belongs to the cyclohydrolase family of enzymes, ranging from 190 to 250 amino acid residues, and is invariably present in many biological systems, including bacteria, fungi, plants, and higher animals, and shown to be conserved on eukaryotic as well as on bacterial genomes [32]. GTPCHI is the rate-limiting enzyme involved in the biosynthesis of tetrahydrobiopterin (BH4), a key cofactor necessary for nitric oxide synthase and for the hydroxylases that are involved in the production of catecholamines and serotonin [33]. GTPCHI requires zinc ions as a cofactor and involves two hydrolyses and an Amadori rearrangement, yielding formic acid as a byproduct [34]. GTPCHI is encoded by the folE gene. Folates are essential cofactors for one-carbon transfer reactions in most living organisms [35]. There is mounting evidence that folate influences arsenic metabolism [36], increasing methylation of inorganic to DMAs [37]. Overexpression of folE ensures a sufficient supply of folate. The expression of GTPCHI from E. coli in transgenic Arabidopsis resulted in a 1250-fold and 2- to 4-fold enhancement of pterins and folates, respectively [38]. Overexpression of the folKE gene in Lactococcus lactis increased folate production by almost 3-fold [39]. The presence of folate is essential for life and the folate biosynthesis pathway appears to be a main target of toxicity of various arsenical species. We therefore propose that overexpression of folE protects cells by ensuring that sufficient folate is produced even under exposure to various arsenic and antimony species (Figure 3).
Acr3 is an As(III) and Sb(III) efflux pump, which is a member of the BART (bile/arsenite/riboflavin transporter) superfamily and includes members found in bacteria, archaea, and fungi, and is more widely distributed than members of the ArsB family [40,41]. The Acr3 protein is present in the Saccharomyces cerevisiae plasma membrane and pumps As(III), but not As(V), Sb(III), tellurite (Te(III)), Cd(II), or phenylarsine oxide out of the cell in response to the proton motive force [42]. Cells of E. coli expressing AmAcr3-1 or CgAcr3-1 confer As(III) resistance. In addition, everted membrane vesicles prepared from Corynebacterium glutamicum expressing Acr3-1 were shown to transport As(III) but not Sb(III) using NADH and an electron donor for the respiratory chain [41,43]. Our results show that Acr3 not only conferred resistance to As(III), but also to Sb(III) (Figure 4). Resistance to high levels of Sb(V) might be due to low levels of reduction of Sb(V) to Sb(III) and subsequent efflux of Sb(III).
4. Materials and Methods
4.1. Strain Isolation
One gram of the soil from the Zijin gold and copper mine (Longyan, Fujian Province, China) wastewater treatment (25°09.724′ N, 116°23.258′ E), at an altitude of 192 m, was added into a 50 mL flask containing 9 mL of phosphate-buffered saline solution (pH 7.0, 0.2 M). The flask was incubated with shaking at 28 °C, 180 rpm for 6 h, to activate the microbial life inside the soil. After 6 h, 100 μL of the above soil solution was plated on R2A plates containing 10 μM of Au(III), and incubated at 28 °C until colonies appeared. Then, single colonies, with different phenotypes, were individually picked up and transferred to an R2A plate containing increasingly higher concentrations of Au(III). The Au(III) concentration ranged from 10 to 160 μM on the R2A plates. The strain grown on the R2A plate containing 160 μM of Au(III) was kept and purified by plating on the R2A plate with the same concentration of Au(III) 3–5 times. Finally, 8 strains were harvested and stored at −80 °C in 15% glycol (V:V). MIC results showed that both strains Au-Bre29 and Au-Bre30 had high heavy-metal resistance to Au(III), Cu(II) and As(III), but with differences in As(III) and Cu(II) resistance; furthermore, strains Au-Bre29 and Au-Bre30 belong to the species Brevundimonas nasdae, so these two strains were selected for further study.
4.2. Heavy-Metal Concentration Measurements of the Sampled Soil
The soil samples were digested by a microwave digestion system according to USEPA Method 3051A (USEPA, 2007) [44], and further determined by inductively coupled plasma mass spectrometry (ICP-MS) (PerkinElmer NexION 300X, Waltham, MA, USA).
4.3. Strain Characterization
4.3.1. Determination of Minimum Inhibitory Concentration (MIC) of Metal(loid)s on R2A Plates
R2A plates were used for the MIC test for isolates of different metal(loid)s since not all isolates can grow in MM medium, and the TY medium contains too many nutrients. First, isolates were activated from −80 °C on the R2A plate, at 28 °C, until a single colony appeared. Then, single colonies were individually transferred into R2A medium cultured at 28 °C, at 180 rpm overnight. The OD600nm of the above strain culture was adjusted to be 0.1, then diluted with R2A medium 100 times. Finally, the diluted culture was streaked on R2A plates containing 0.25–4 mM of Cu(II), 0.25–6 mM of Cd(II), and 0.25–6 mM of As(III), with a 0.25 increment, respectively. The plates were inoculated at 28 °C for 3 days. The R2A plate containing the lowest concentration of metal(loid)s, where no visible growth of strains was observed after 3 days, was recorded as the MIC.
4.3.2. Molecular Identification of Isolates
For molecular identification, the genomic DNA of strain Au-Bre30 was extracted using the TIANamp Bacteria DNA kit under the guidance of the manufacturer’s instructions (Tiangen Biotech, Beijing, China). The 16S rRNA gene was amplified using the primers (27F and 1492R) with genomic DNA as a template [45]. One percent agarose gel was used to verify the amplification of the 16S rRNA product by running gel, and after that, the confirmed PCR product was sent out to the Sangon Biotech company (Shanghai, China) for sequencing. Partial 16S rRNA gene sequences were uploaded and subjected to NCBI to identify the most related species based on the most homology. A phylogenetic tree was constructed using assembled sequences of 16S rRNA genes and the neighbor-joining algorithms via MEGA X [46].
4.3.3. Determination of Optimal Growth Conditions of B. nasdae Au-Bre29 and Au-Bre30
Single colonies of strains Au-Bre29 and Au-Bre30 that appeared on the R2A plate were transferred into liquid R2A medium and incubated with shaking at 28 °C. Thereafter, the strain culture was transferred into fresh R2A medium and cultured under the same conditions overnight. The OD600nm of the above strain culture was adjusted to be 1.0 for further study. One percent of strain culture with OD600nm adjusted was transferred into new R2A medium with different pH levels (2–12, additional NaCl = 0%), different additional NaCl (0–10%, pH = 7.0) concentrations, and different medium (LB, MM, R2A, and TY, pH = 7.0), individually. LB medium contained 10 g/L of tryptone, 5 g/L of yeast extract, and 10 g/L of NaCl, with pH adjusted to 7.0 by HCl or NaOH [47]. MM medium contained 2.0 g/L of sodium gluconate, 6.06 g/L of trimethylaminomethane, 4.68 g/L of NaCl, 1.49 g/L of KCl, 1.07 g/L of NH4Cl, 0.43 g/L of Na2SO4, 0.005 g/L of ferric ammonium citrate, 0.2 g/L of MaCl2·6H2O, 0.03 g/L of CaCl2·2H2O, 0.23 g/L of Na2HPO4·12H2O, 1.5 mg/L of FeCl2·4H2O, 0.19 mg/L of CoCl2·6H2O, 0.1 mg/L of MnCl2·4H2O, 62 μg/L of H3BO3, 70 μg/L of ZnCl2, 36 μg/L of Na2MoO4·2H2O, 24 μg/L of NiCl2·6H2O, and 17 μg/L of CuCl2·2H2O, with pH adjusted to 7.0 by HCl or NaOH [26]. R2A medium contained 0.5/L g of tryptone, 0.5 g/L of yeast extract, 0.5 g/L of casamino acid, 0.5 g/L of glucose, 0.5 g/L of soluble starch, 0.3 g/L of K2HPO4, 0.024 g/L of MgSO4, and 0.3 g/L of sodium pyruvate, with pH adjusted to 7.0 by HCl or NaOH [48]. TY medium contained 5 g/L of tryptone, 3 g/L of yeast extract, and 1.3 g/L of CaCl2·6H2O, with pH adjusted to 7.0 by HCl or NaOH [49]. Then, 200 μL of each diluted culture was loaded into separate wells of a 100-well honeycomb plate in 3 duplicates. A cover was used on the plate to prevent evaporation. Growth experiments were performed in the Bioscreen C microbiology reader (Oy Growthcurves Ab Ltd., Helsinki, Finland) at 28 °C for 48 h, and the optical densities at 600nm (OD600nm) were automatically measured at 4 h intervals.
4.3.4. Nanopore Sequencing, Annotation, and Assembly
High-quality genomic DNA with OD260/280 values higher than 1.80 from strains Au-Bre29 and Au-Bre30 were extracted following the instructions of the TIANamp Bacteria DNA Kit (Tiangen Biotech, Beijing, China). DNA fragments with size > 20 Kb were collected and sent to Biomarker Biotech Co., Ltd. (Beijing, China) for sequencing by using the PromethION platform (R9.4.1, FLO-PRO002; Oxford Nanopore Technologies (ONT), Oxford, UK). The SQK-LSK109 kit was used to construct the DNA library under the guidance of the standard protocol of ONT. The NEBNext® Ultra™ DNA Library Prep Kit (New England Biolabs, Beijing, China) was used for DNA sequencing via the Illumina Novaseq 6000 platform, with an insert size of 300 bp (paired end). The sequence was assembled by using Canu (v1.5), and annotated by the NCBI Prokaryotic Genome Annotation Pipeline (PGAP) with GenBank accession numbers CP080034-CP080035 (Au-Bre29) and CP080036 (Au-Bre30) [50].
4.4. Identification of Putative ars Genes
4.4.1. Plasmids, Bacterial Strains, and Growth Conditions
Competent cell of E. coli DH5α, purchased from TransGen Biotech (Beijing, China), was used for plasmid DNA replication. The As(III)-hypersensitive strain, E. coli AW3110 (ΔarsRBC::cam) [51], was used for the following complementation studies. The PCR products of putative arsenic resistance-related genes were cloned into plasmid pTOPO by using the Zero Background pTOPO-Blunt Simple Cloning Kit under the guidance of the manufacturer (Aidlab, Beijing, China). The strains bearing the indicated plasmid were grown aerobically in LB medium containing 100 μg/mL of ampicillin (Amp). The absorbance at OD600nm was used to measure the growth of the strains. The three putative arsenic resistance-related genes were designated as folE, acr3, and arsH from B. nasdae Au-Bre30. The FolE, Acr3, and ArsH had GenBank accession numbers QYC15516, QYC15513, and QYC15515, respectively, and were cloned into the pTOPO plasmid. Furthermore, the pTOPO plasmid bearing indicated genes was first transferred into DH5α to replicate, and then transferred into AW3110 to complement.
4.4.2. Metal(loid)s Resistance Analysis
A single colony of E. coli strain AW3110 harboring the indicated plasmid was grown in LB medium containing 100 μg/mL of Amp at 37 °C, 220 rpm. After 12 h of incubation, the OD600nm of strain culture was adjusted to 0.1 with LB added, and then the adjusted strain culture was transferred into new LB medium supplied with 100 μg/mL of Amp at 1% and cultured under the same conditions for around 12 h. The absorbance at OD600nm of the overnight cultured strain was adjusted to 1.0. The adjusted strain culture was diluted 100-fold in LB medium containing 100 μg/mL of Amp and different concentrations of metal(loid)s and shaken for 24 h. The growth of strain E. coli AW3110 bearing different plasmids was measured by the value of OD600nm.
5. Conclusions
We characterized two highly arsenite-resistant B. nasdae strains, Au-Bre29 and Au-Bre30, and identified the novel arsenic resistance determinant folE in B. nasdae Au-Bre30, encoding GTP-Cyclohydrolase I (FolE). Heterologous expression of folE in the arsenic-hypersensitive E. coli strain AW3110 resulted in increased resistance to As(III), As(V), Rox(III), Rox(V), Sb(III), and Sb(V). Heterologous expression of arsH and acr3 in AW3110 resulted in increased resistance to As(III), Rox(III), Sb(III), and Sb(V). arsH also conferred resistance to As(V). These results indicated that expression of arsH conferred resistance to ROS generated due to As or Sb exposure.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23105619/s1.
Author Contributions
Conceptualization, W.X. and C.R.; formal analysis, investigation, X.Y. and Y.L.; validation, C.R.; writing—original draft preparation, X.Y. and Y.L.; writing—review and editing, critical revision, C.R., W.X., R.F., J.C. and H.A.A.; funding acquisition, C.R., W.X. and H.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by NSFC (grant number 31770123). We also acknowledge Researchers Supporting Project number RSP-2022/205, King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brammer, H.; Ravenscroft, P. Arsenic in groundwater: A threat to sustainable agriculture in South and South-east Asia. Environ. Int. 2009, 35, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Tseng, C.P.; Chiou, H.Y.; Hsueh, Y.M.; Chong, C.K.; Chen, C.-J. Epidemiologic evidence of diabetogenic effect of arsenic. Toxicol. Lett. 2002, 133, 69–76. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, C.; Wu, M.; Kuo, T. Cancer potential in liver, lung, bladder and kidney due to ingested inorganic arsenic in drinking water. Br. J. Cancer 1992, 66, 888–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.G.; Yoshinaga, M.; Zhao, F.J.; Rosen, B.P. Earth Abides Arsenic Biotransformations. Annu. Rev. Earth Planet. Sci. 2014, 42, 443–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolz, J.F.; Basu, P.; Santini, J.M.; Oremland, R.S. Arsenic and selenium in microbial metabolism. Annu. Rev. Microbiol. 2006, 60, 107–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef]
- Chen, B. Antimony in the environment: A review focused on natural waters: I. Occurrence. Earth Sci. Rev. 2002, 57, 125–176. [Google Scholar]
- Chen, J.; Rosen, B.P. The Arsenic Methylation Cycle: How Microbial Communities Adapted Methylarsenicals for Use as Weapons in the Continuing War for Dominance. Front. Environ. Sci. 2020, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, Q.; Oremland, R.S.; Kulp, T.R.; Rensing, C.; Wang, G. Microbial Antimony Biogeochemistry: Enzymes, Regulation, and Related Metabolic Pathways. Appl. Environ. Microbiol. 2016, 82, 5482–5495. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.C.; Fu, H.L.; Lin, Y.F.; Rosen, B.P. Pathways of arsenic uptake and efflux. Curr. Top. Membr. 2012, 69, 325–358. [Google Scholar]
- Giovannoni, S.J.; Halsey, K.H.; Saw, J.; Muslin, O.; Suffridge, C.P.; Sun, J.; Lee, C.P.; Moore, E.R.; Temperton, B.; Noell, S.E. A Parasitic Arsenic Cycle That Shuttles Energy from Phytoplankton to Heterotrophic Bacterioplankton. Mbio 2019, 10, e00246-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, J.; Rosen, B.P. Role of ArsEFG in Roxarsone and Nitarsone Detoxification and Resistance. Environ. Sci. Technol. 2019, 53, 6182–6191. [Google Scholar] [CrossRef] [PubMed]
- Galván, A.E.; Paul, N.P.; Chen, J.; Yoshinaga-Sakurai, K.; Utturkar, S.M.; Rosen, B.P.; Yoshinaga, M. Identification of the Biosynthetic Gene Cluster for the Organoarsenical Antibiotic Arsinothricin. Microbiol. Spectr. 2021, 9, e00502-21. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, J.; Wu, Y.F.; Zhao, F.J.; Rosen, B.P. ArsV and ArsW provide synergistic resistance to the antibiotic methylarsenite. Environ. Microbiol. 2021, 23, 7550–7562. [Google Scholar] [CrossRef]
- Qin, J.; Rosen, B.P.; Zhang, Y.; Wang, G.; Franke, S.; Rensing, C. Arsenic detoxification and evolution of trimethylarsine gas by a microbial arsenite S-adenosylmethionine methyltransferase. Proc. Natl. Acad. Sci. USA 2006, 103, 2075–2080. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, N.S.; Ranjan, R.; Purohit, H.J.; Kalia, V.C.; Sharma, R. Identification of genes conferring arsenic resistance to Escherichia coli from an effluent treatment plant sludge metagenomic library. FEMS Microbiol. Ecol. 2009, 67, 130–139. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, J.; Rosen, B.P. Organoarsenical tolerance in Sphingobacterium wenxiniae, a bacterium isolated from activated sludge. Environ. Microbiol. 2022, 24, 762–771. [Google Scholar] [CrossRef]
- Chen, J.; Bhattacharjee, H.; Rosen, B.P. ArsH is an organoarsenical oxidase that confers resistance to trivalent forms of the herbicide monosodium methylarsenate and the poultry growth promoter roxarsone. Mol. Microbiol. 2015, 96, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yoshinaga, M.; Garbinski, L.D.; Rosen, B.P. Synergistic interaction of glyceraldehydes-3-phosphate dehydrogenase and ArsJ, a novel organoarsenical efflux permease, confers arsenate resistance. Mol. Microbiol. 2016, 100, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Li, C.; Rensing, C.; Dai, X.; Fan, X.; Wang, G. Efflux Transporter ArsK Is Responsible for Bacterial Resistance to Arsenite, Antimonite, Trivalent Roxarsone, and Methylarsenite. Appl. Environ. Microbiol. 2018, 84, e01842-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Madegowda, M.; Bhattacharjee, H.; Rosen, B.P. ArsP: A methylarsenite efflux permease. Mol. Microbiol. 2015, 98, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, R.; Rosen, B.P. Arsenate reductases in prokaryotes and eukaryotes. Environ. Health Perspect. 2002, 110, 745–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Maury, L.; Sanchez-Riego, A.M.; Reyes, J.C.; Florencio, F.J. The glutathione/glutaredoxin system is essential for arsenate reduction in Synechocystis sp. strain PCC 6803. J. Bacteriol. 2009, 191, 3534–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Chen, S.; Xiao, X.; Huang, X.; You, D.; Zhou, X.; Deng, Z. arsRBOCT arsenic resistance system encoded by linear plasmid pHZ227 in Streptomyces sp. strain FR-008. Appl. Environ. Microbiol. 2006, 72, 3738–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Yang, S.M.; Kim, D.; Kim, H.Y. Complete Genome Sequencing and Comparative Genomics of Three Potential Probiotic Strains, Lacticaseibacillus casei FBL6, Lacticaseibacillus chiayiensis FBL7, and Lacticaseibacillus zeae FBL8. Front. Microbiol. 2022, 12, 794315. [Google Scholar] [CrossRef] [PubMed]
- Su, J. Isolation, Identification and Arsenic Resistant Mechanism Analysis of an Arsenic Tolerant Bacteria Paenibacillus taichungensis NC1 (In Chinese). Master Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2020. [Google Scholar]
- Moe, B.; Peng, H.; Lu, X.; Chen, B.; Chen, L.W.L.; Gabos, S.; Li, X.F.; Le, X.C. Comparative cytotoxicity of fourteen trivalent and pentavalent arsenic species determined using real-time cell sensing. J. Environ. Sci. 2016, 49, 113–124. [Google Scholar] [CrossRef]
- Wilson, S.C.; Lockwood, P.V.; Ashley, P.M.; Tighe, M. The chemistry and behaviour of antimony in the soil environment with comparisons to arsenic: A critical review. Environ. Pollut. 2010, 158, 1169–1181. [Google Scholar] [CrossRef]
- Paez-Espino, A.D.; Nikel, P.I.; Chavarria, M.; de Lorenzo, V. ArsH protects Pseudomonas putida from oxidative damage caused by exposure to arsenic. Environ. Microbiol. 2020, 22, 2230–2242. [Google Scholar] [CrossRef]
- Gu, J.; Yao, J.; Duran, R.; Sunahara, G. Comprehensive genomic and proteomic profiling reveal Acinetobacter johnsonii JH7 responses to Sb(III) toxicity. Sci. Total Environ. 2020, 748, 141174. [Google Scholar] [CrossRef]
- Rong, Q.; Ling, C.; Lu, D.; Zhang, C.; Zhao, H.; Zhong, K.; Nong, X.; Qin, X. Sb(III) resistance mechanism and oxidation characteristics of Klebsiella aerogenes X. Chemosphere 2022, 293, 133453. [Google Scholar] [CrossRef]
- Maier, J.; Witter, K.; Gutlich, M.; Ziegler, I.; Werner, T.; Ninnemann, H. Homology cloning of GTP-cyclohydrolase I from various unrelated eukaryotes by reverse-transcription polymerase chain reaction using a general set of degenerate primers. Biochem. Biophys. Res. Commun. 1995, 212, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Maita, N.; Hatakeyama, K.; Okada, K.; Hakoshima, T. Structural basis of biopterin-induced inhibition of GTP cyclohydrolase I by GFRP, its feedback regulatory protein. J. Biol. Chem. 2004, 279, 51534–51540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerbach, G.; Herrmann, A.; Bracher, A.; Bader, G.; Gutlich, M.; Fischer, M.; Neukamm, M.; Garrido-Franco, M.; Richardson, J.; Nar, H. Zinc plays a key role in human and bacterial GTP cyclohydrolase I. Proc. Natl. Acad. Sci. USA 2000, 97, 13567–13572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basset, G.; Quinlivan, E.P.; Gregory, J.F.; Hanson, A.D. Folate synthesis and metabolism in plants and prospects for biofortification. Crop Sci. 2005, 45, 449–453. [Google Scholar] [CrossRef] [Green Version]
- Kile, M.L.; Ronnenberg, A.G. Can folate intake reduce arsenic toxicity? Nutr. Rev. 2008, 66, 349–353. [Google Scholar] [CrossRef]
- Bozack, A.K.; Hall, M.N.; Liu, X.; Ilievski, V.; Lomax-Luu, A.M.; Parvez, F.; Siddique, A.B.; Shahriar, H.; Uddin, M.N.; Islam, T. Folic acid supplementation enhances arsenic methylation: Results from a folic acid and creatine supplementation randomized controlled trial in Bangladesh. Am. J. Clin. Nutr. 2019, 109, 380–391. [Google Scholar] [CrossRef]
- Hossain, T.; Rosenberg, I.; Selhub, J.; Kishore, G.; Beachy, R.; Schubert, K. Enhancement of folates in plants through metabolic engineering. Proc. Natl. Acad. Sci. USA 2004, 101, 5158–5163. [Google Scholar] [CrossRef] [Green Version]
- Sybesma, W.; Starrenburg, M.; Kleerebezem, M.; Mierau, I.; de Vos, W.M.; Hugenholtz, J. Increased production of folate by metabolic engineering of Lactococcus lactis. Appl. Environ. Microbiol. 2003, 69, 3069–3076. [Google Scholar] [CrossRef] [Green Version]
- Mansour, N.M.; Sawhney, M.; Tamang, D.G.; Vogl, C.; Saier, M.H., Jr. The bile/arsenite/riboflavin transporter (BART) superfamily. FEBS J. 2007, 274, 612–629. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.L.; Meng, Y.; Ordonez, E.; Villadangos, A.F.; Bhattacharjee, H.; Gil, J.A.; Mateos, L.M.; Rosen, B.P. Properties of arsenite efflux permeases (Acr3) from Alkaliphilus metalliredigens and Corynebacterium glutamicum. J. Biol. Chem. 2009, 284, 19887–19895. [Google Scholar] [CrossRef] [Green Version]
- Wysocki, R.; Bobrowicz, P.; Ulaszewski, S. The Saccharomyces cerevisiae ACR3 gene encodes a putative membrane protein involved in arsenite transport. J. Biol. Chem. 1997, 272, 30061–30066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbinski, L.D.; Rosen, B.P.; Chen, J. Pathways of arsenic uptake and efflux. Environ. Int. 2019, 126, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Lopéz, Y.P.; da Fonseca Breda, F.A.; Lima, E.S.A.; da Costa Barros de Souza, C.; González, J.M.F.; do Amaral Sobrinho, N.M.B. Variability factors of heavy metals in soils and transfer to pasture plants of Mayabeque in Cuba. Environ. Monit. Assess. 2021, 193, 245. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhou, X.F.; Yang, S.J.; Liu, W.H.; Hu, X.F. Design and application of specific 16S rDNA-targeted primers for assessing endophytic diversity in Dendrobium officinale using nested PCR-DGGE. Appl. Microbiol. Biotechnol. 2013, 97, 9825–9836. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Ruan, J.; Li, Y.; Qiu, R. A new model for simulating microbial cyanide production and optimizing the medium parameters for recovering precious metals from waste printed circuit boards. J. Hazard. Mater. 2018, 353, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, Y.; Wu, Y.; Qiu, Z.; Ding, Z.; Wang, X.; Chen, W.; Wang, R.; Fu, F.; Rensing, C.; et al. Improved grain yield and lowered arsenic accumulation in rice plants by inoculation with arsenite-oxidizing Achromobacter xylosoxidans GD03. Ecotoxicol. Environ. Saf. 2020, 206, 111229. [Google Scholar] [CrossRef]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Carlin, A.; Shi, W.; Dey, S.; Rosen, B.P. The ars operon of Escherichia coli confers arsenical and antimonial resistance. J. Bacteriol. 1995, 177, 981–986. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).