
Piperine Improves Lipid Dysregulation by Modulating Circadian Genes Bmal1 and Clock in HepG2 Cells


Abstract
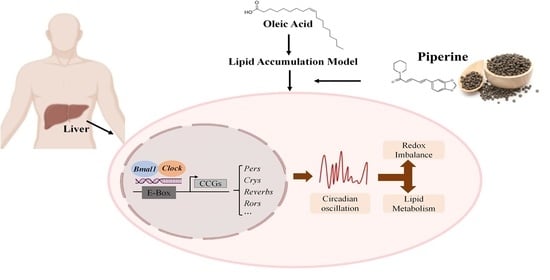
:
1. Introduction
2. Results and Discussion
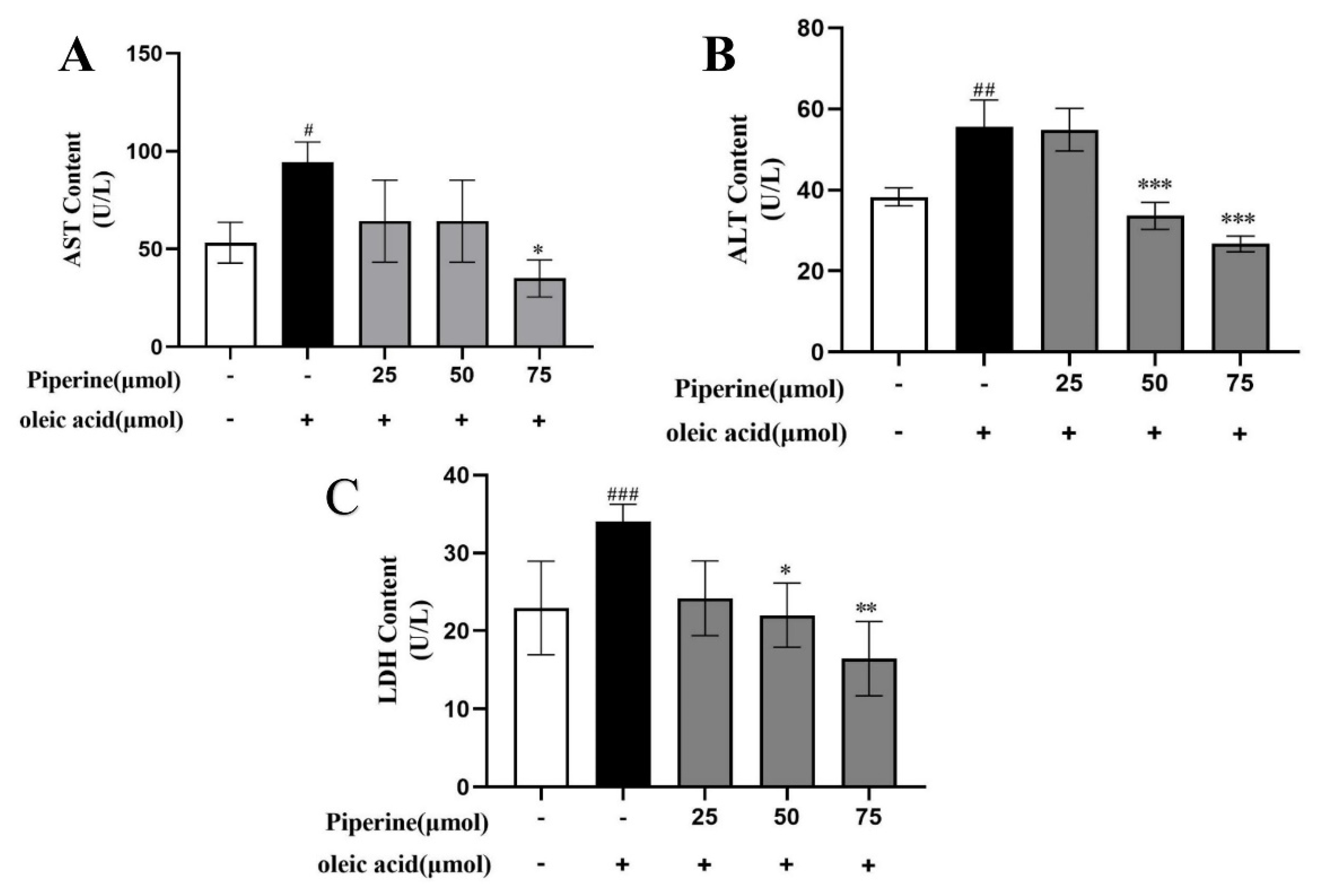
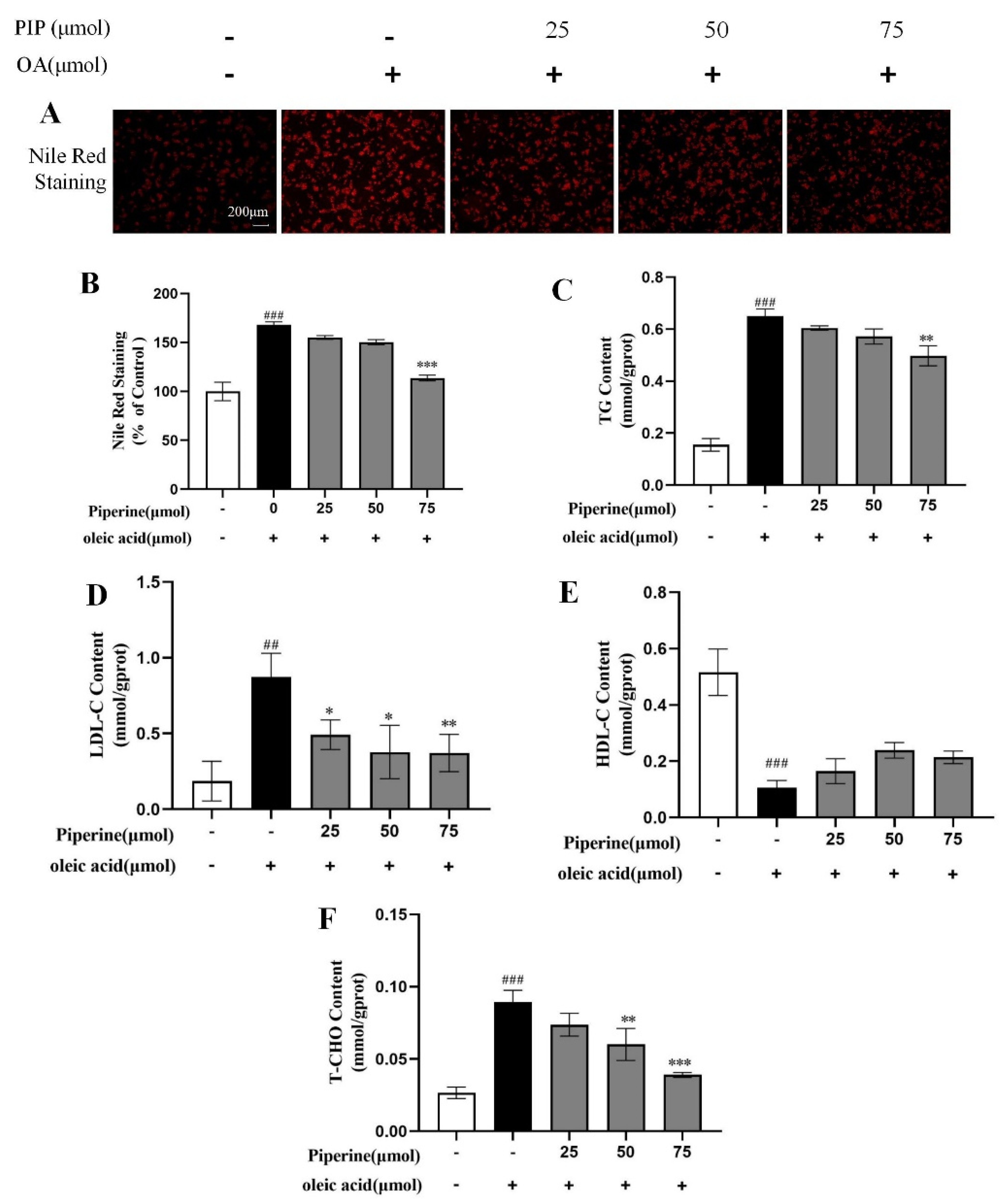
2.1. Preventive Effects of PIP on OA-Induced Lipid Accumulation in HepG2 Cells
2.2. Inhibitory Effects of PIP on OA-Induced Lipids Accumulation in HepG2 Cells
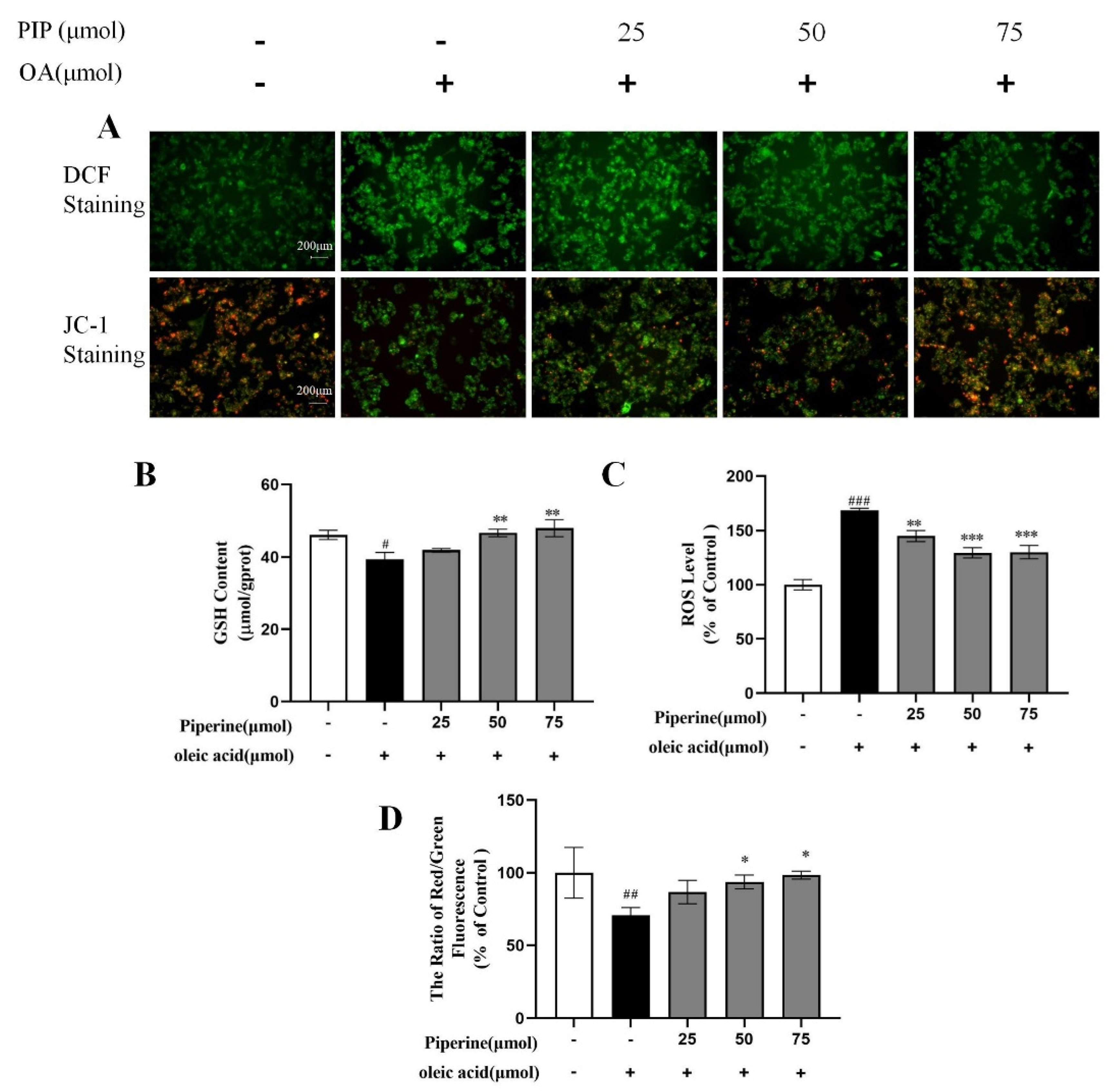
2.3. Effects of PIP on OA-Induced Mitochondrial Dysfunction and ROS Overproduction in HepG2 Cells
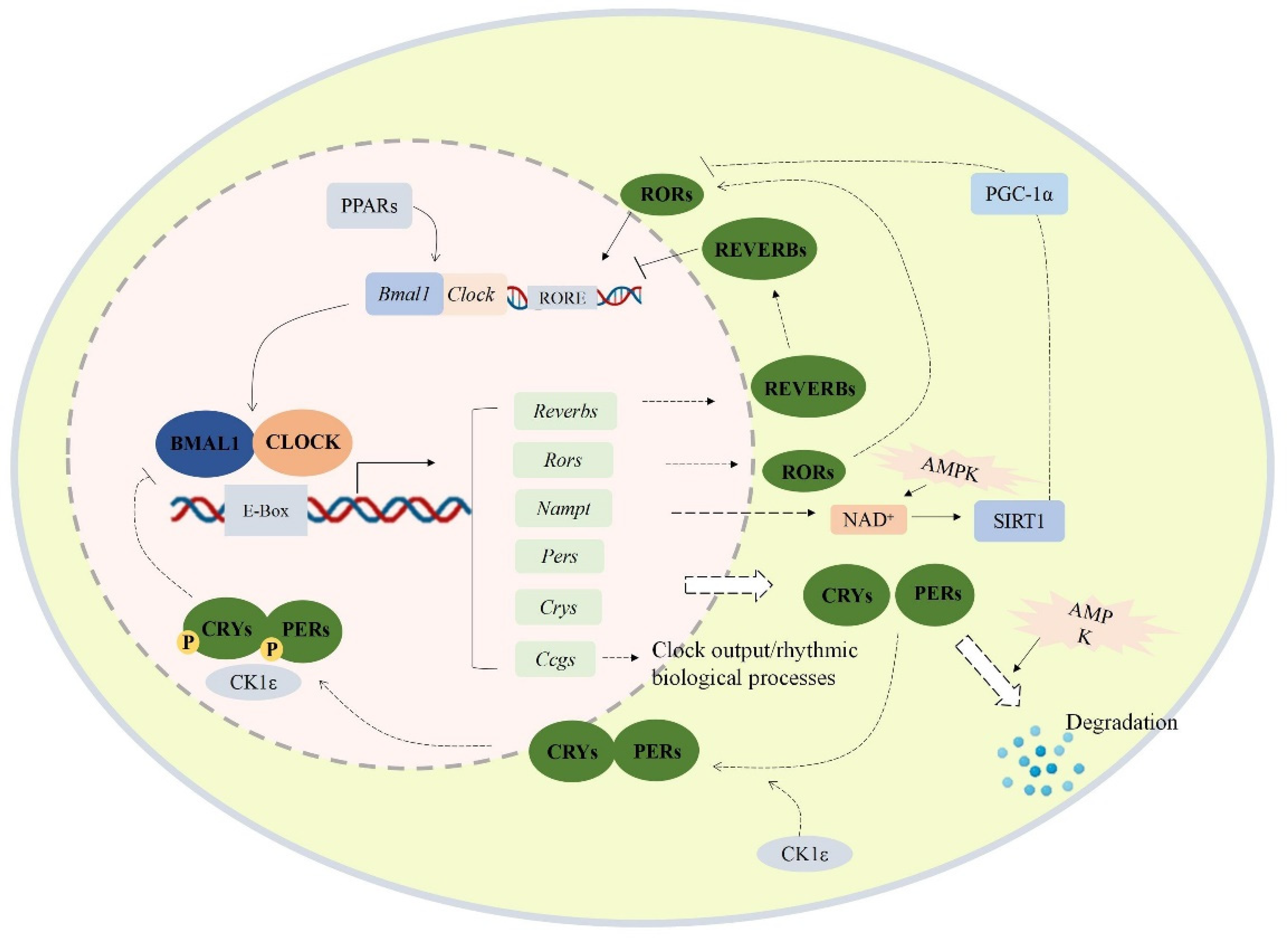
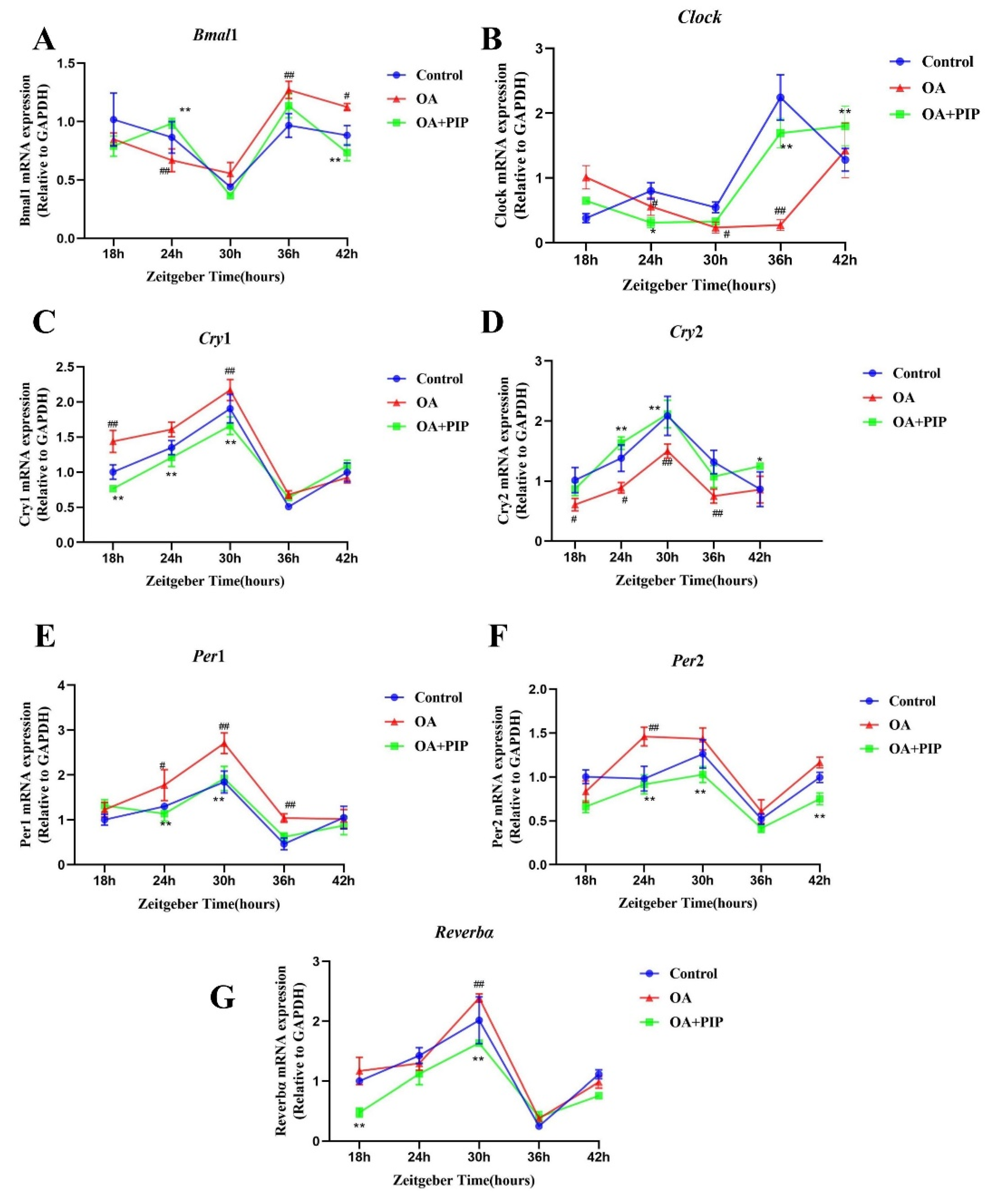
2.4. Intervention Effect of PIP on Circadian Misalignment
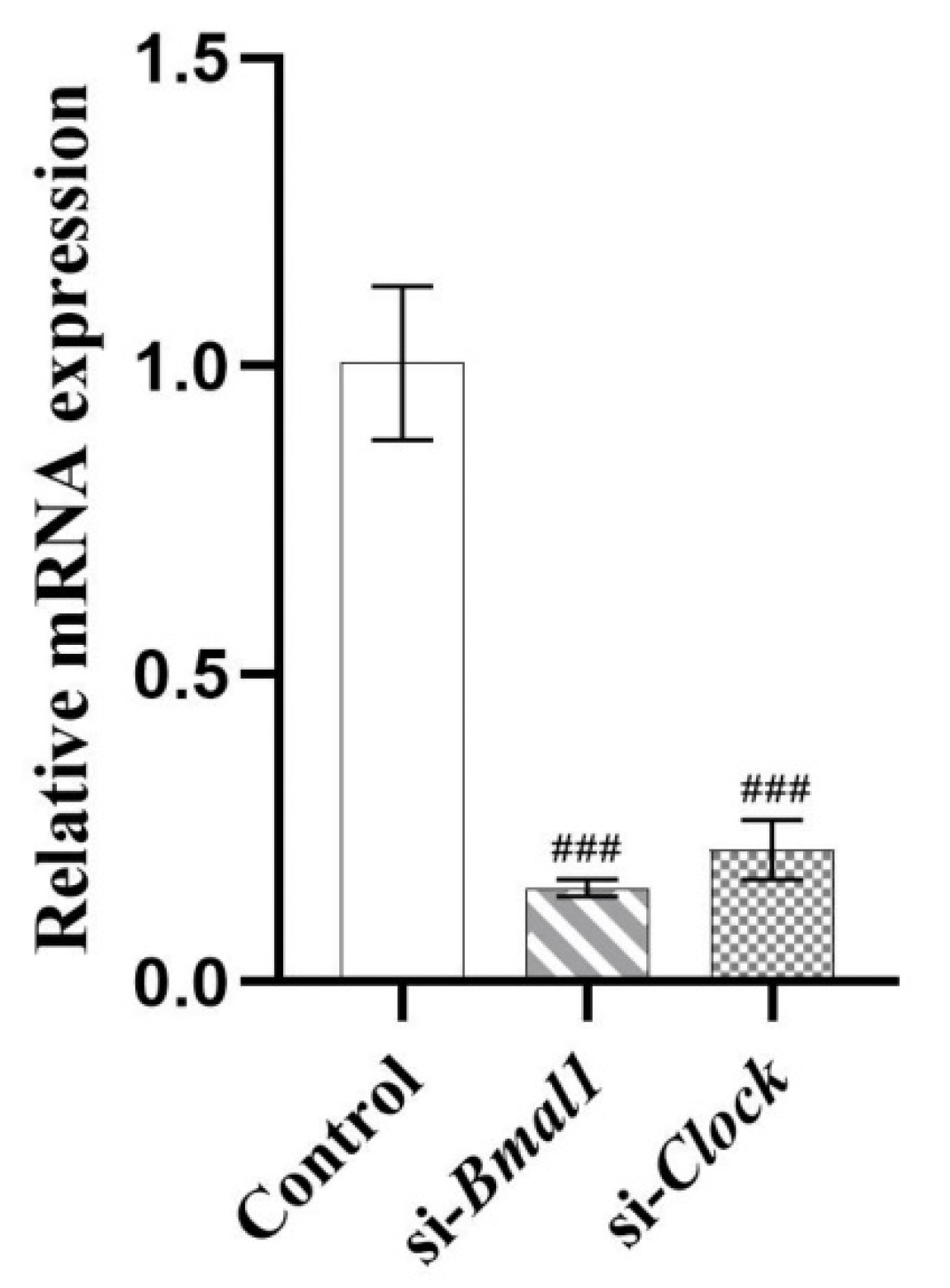
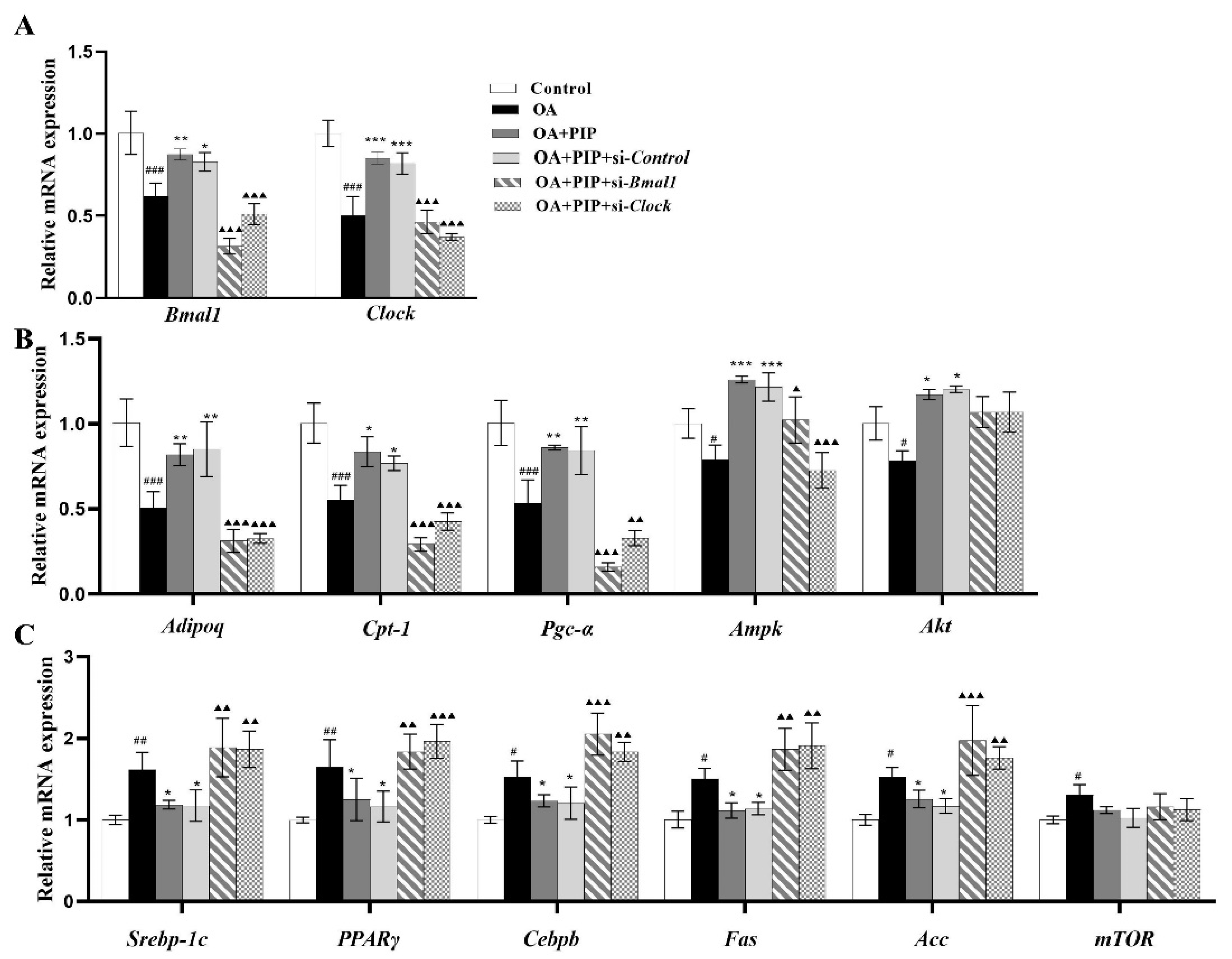
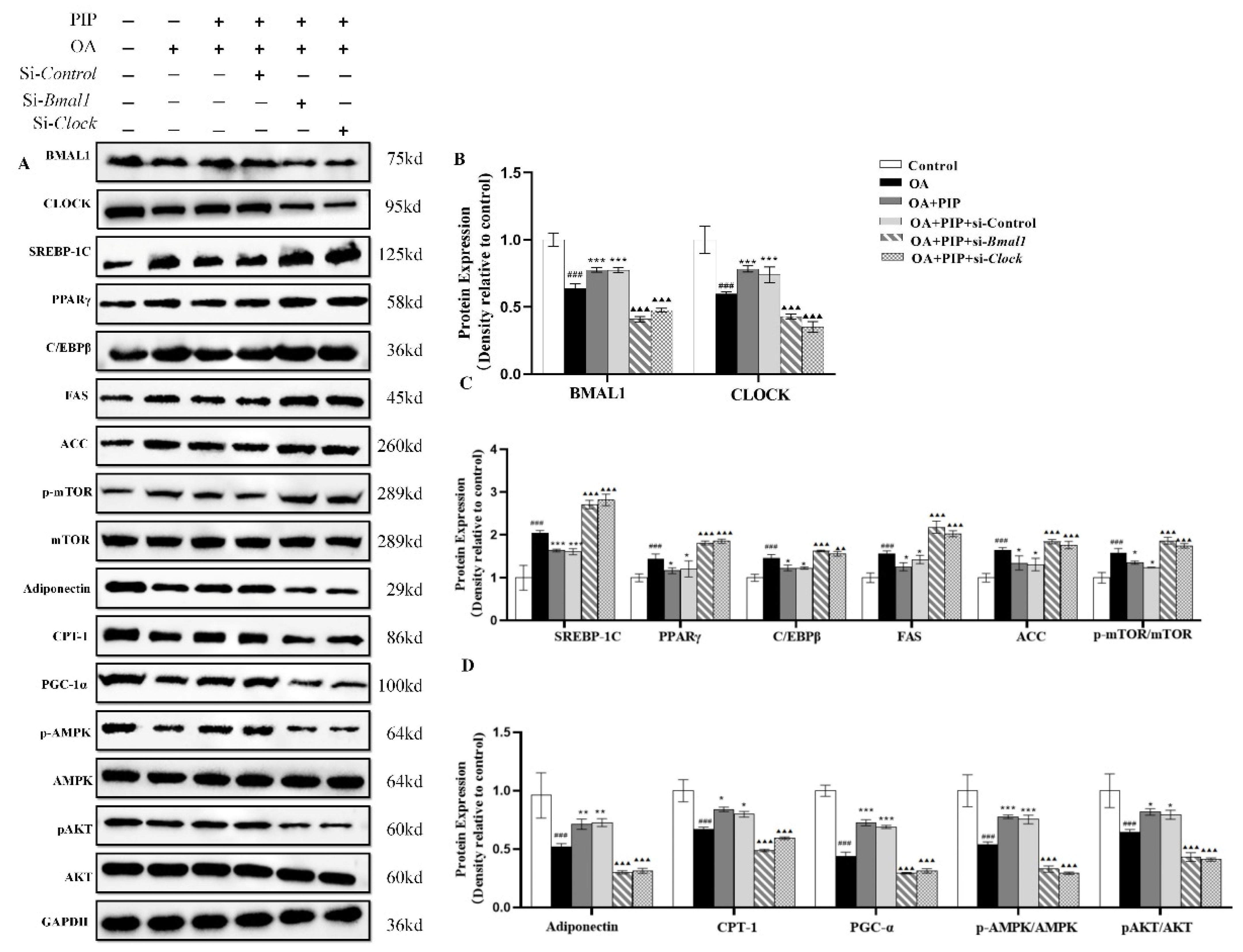
2.5. PIP Attenuated OA-Induced Redox Imbalance and Mitochondrial Dysfunction via Regulating Bmal1 and Clock in HepG2 Cells
2.6. Effects of PIP on OA-Induced Lipid Metabolism Disorder Imbalance in a Bmal1/Clock-Dependent Way
3. Materials and Methods
3.1. Materials
3.2. Cell Culture and Viability Assay
3.3. Measurement of Lipid Profiles and Enzyme Activity in HepG2 Cells
3.4. Transfection of HepG2 Cells
3.5. JC-1, DCFDA, and Nile Red Staining
3.6. RNA Isolation and Real-Time qPCR
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACC | acetyl coenzyme A carboxylase |
| ALT | alanine aminotransferase |
| AMPK | AMP-activated protein kinase |
| AST | aspartate aminotransferase |
| BMAL1 | aryl hydrocarbon receptor nuclear translocatorlike 1 proteins |
| Bmal1 | aryl hydrocarbon receptor nuclear translocatorlike 1 |
| C/EBPβ | adipogenesis-associated transcription factor CCAAT enhancer binding proteins β |
| Ccgs | other clock-controlled genes |
| CLOCK | circadian locomotor output cycles kaput proteins |
| Clock | circadian locomotor output cycles kaput |
| CPT-1 | carnitine palmitoyl transferase |
| CRYs | cryptochrome proteins |
| Crys | cryptochrome genes 1/2 |
| DCF | dichlorofluorescin |
| DCFH-DA | 2,7-dichlorofluorescin diacetate |
| FAS | fatty acid synthase |
| HDL-C | HDL cholesterol |
| LDH | lactate dehydrogenase |
| LDL-C | LDL cholesterol |
| MMP | mitochondrial membrane potential |
| mTOR | mammalian/mechanistic target of rapamycin |
| NAD+ | nicotinamide adenine dinucleotide |
| Nampt | nicotinamide phosphoribosyltransferase gene |
| OA | oleic acid |
| PERs | period proteins |
| Pers | period genes 1/2/3 |
| PGC-1α | peroxlsome proliferator-activated receptor-γ coactlvator-1α |
| PPARs | peroxisome proliferator-activated receptors |
| PPARγ | peroxisome proliferator activated receptor gamma |
| Reverbs | reverse erythroblastosis virus α/β |
| RORE | ROR response element |
| RORs | retinoic acid-related orphan receptors |
| ROS | reactive oxygen species |
| SCN | suprachiasmatic nucleus |
| SIRT1 | silent information regulator 1 |
| SREBP-1c | sterol-regulatory-element-binding protein 1c |
| TC | total cholesterol |
| TG | total triglyceride |
| TP | total protein |
| TTFL | transcriptional/translational feedback loop |
References
- Yan, R.; Yang, C.S.; Zhang, X. Maintain host health with time-restricted eating and phytochemicals: A review based on gut microbiome and circadian rhythm. Trends Food Sci. Technol. 2021, 108, 258–268. [Google Scholar] [CrossRef]
- Huang, J.Q.; Lu, M.; Ho, C.T. Health benefits of dietary chronobiotics: Beyond resynchronizing internal clocks. Food Funct. 2021, 12, 6136–6156. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.; Hsu, J.W.; Manka, P.P.; Syn, W.K. Role of the Circadian Clock in the Metabolic Syndrome and Nonalcoholic Fatty Liver Disease. Dig. Dis. Sci. 2018, 63, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front. Microbiol. 2018, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Zhao, B.; Tan, X.; Wang, L.; Liu, X. Role of Food Phytochemicals in the Modulation of Circadian Clocks. J. Agric. Food Chem. 2019, 67, 8735–8739. [Google Scholar] [CrossRef] [PubMed]
- Stenvers, D.J.; Scheer, F.; Schrauwen, P.; la Fleur, S.E.; Kalsbeek, A. Circadian clocks and insulin resistance. Nat. Rev. Endocrinol. 2019, 15, 75–89. [Google Scholar] [CrossRef]
- Lekkas, D.; Paschos, G.K. The circadian clock control of adipose tissue physiology and metabolism. Auton. Neurosci. 2019, 219, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Pan, S.; Xu, P.; Xue, T.; Wang, J.; Guo, Y.; Jia, L.; Qiao, X.; Li, L.; Zhai, Y. Melatonin Orchestrates Lipid Homeostasis through the Hepatointestinal Circadian Clock and Microbiota during Constant Light Exposure. Cells 2020, 9, 489. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Ho, C.T.; Zhang, X.; Wu, Z.; Cao, J. Modulatory effect of Cyclocarya paliurus flavonoids on the intestinal microbiota and liver clock genes of circadian rhythm disorder mice model. Food Res. Int. 2020, 138, 109769. [Google Scholar] [CrossRef]
- Shimizu, H.; Hanzawa, F.; Kim, D.; Sun, S.; Laurent, T.; Umeki, M.; Ikeda, S.; Mochizuki, S.; Oda, H. Delayed first active-phase meal, a breakfast-skipping model, led to increased body weight and shifted the circadian oscillation of the hepatic clock and lipid metabolism-related genes in rats fed a high-fat diet. PLoS ONE 2018, 13, e0206669. [Google Scholar]
- Guo, R.; Zhao, B.; Wang, Y.; Wu, D.; Wang, Y.; Yu, Y.; Yan, Y.; Zhang, W.; Liu, Z.; Liu, X. Cichoric Acid Prevents Free-Fatty-Acid-Induced Lipid Metabolism Disorders via Regulating Bmal1 in HepG2 Cells. J. Agric. Food Chem. 2018, 66, 9667–9678. [Google Scholar] [CrossRef]
- Lu, M.; Lan, Y.; Xiao, J.; Song, M.; Chen, C.; Liang, C.; Huang, Q.; Cao, Y.; Ho, C.T. Capsaicin Ameliorates the Redox Imbalance and Glucose Metabolism Disorder in an Insulin-Resistance Model via Circadian Clock-Related Mechanisms. J. Agric. Food Chem. 2019, 67, 10089–10096. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xiao, J.; Cao, Y.; Huang, Q.; Ho, C.T.; Lu, M. Capsaicin Attenuates Oleic Acid-Induced Lipid Accumulation via the Regulation of Circadian Clock Genes in HepG2 Cells. J. Agric. Food Chem. 2021, 70, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Li, L.; Wang, J.; Zhao, B.; Pan, J.; Wang, L.; Liu, X.; Liu, X.; Liu, Z. Resveratrol Prevents Acrylamide-Induced Mitochondrial Dysfunction and Inflammatory Responses via Targeting Circadian Regulator Bmal1 and Cry1 in Hepatocytes. J. Agric. Food Chem. 2019, 67, 8510–8519. [Google Scholar] [PubMed]
- Li, J.; Wei, L.; Zhao, C.; Li, J.; Liu, Z.; Zhang, M.; Wang, Y. Resveratrol Maintains Lipid Metabolism Homeostasis via One of the Mechanisms Associated with the Key Circadian Regulator Bmal1. Molecules 2019, 24, 2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, G.; Wu, W.; Mi, Y.; Shi, R.; Sun, K.; Li, R.; Liu, X.; Liu, X. Tea polyphenols direct Bmal1-driven ameliorating of the redox imbalance and mitochondrial dysfunction in hepatocytes. Food Chem. Toxicol. 2018, 122, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Y.; Song, Y.; Cheng, X.R.; Xia, S.; Rahman, M.R.; Shi, Y.; Le, G. Resveratrol restores the circadian rhythmic disorder of lipid metabolism induced by high-fat diet in mice. Biochem. Biophys. Res. Commun. 2015, 458, 86–91. [Google Scholar] [CrossRef]
- Zhang, W.; Zheng, Q.; Song, M.; Xiao, J.; Cao, Y.; Huang, Q.; Ho, C.T.; Lu, M. A review on the bioavailability, bio-efficacies and novel delivery systems for piperine. Food Funct. 2021, 12, 8867–8881. [Google Scholar] [CrossRef]
- Wu, J.G.; Kan, Y.J.; Wu, Y.B.; Yi, J.; Chen, T.Q.; Wu, J.Z. Hepatoprotective effect of ganoderma triterpenoids against oxidative damage induced by tert-butyl hydroperoxide in human hepatic HepG2 cells. Pharm. Biol. 2016, 54, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Escorcia, W.; Ruter, D.L.; Nhan, J.; Curran, S.P. Quantification of Lipid Abundance and Evaluation of Lipid Distribution in Caenorhabditis elegans by Nile Red and Oil Red O Staining. J. Vis. Exp. 2018, 133, e57352. [Google Scholar] [CrossRef]
- Wu, C.H.; Lin, M.C.; Wang, H.C.; Yang, M.Y.; Jou, M.J.; Wang, C.J. Rutin inhibits oleic acid induced lipid accumulation via reducing lipogenesis and oxidative stress in hepatocarcinoma cells. J. Food Sci. 2011, 76, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ontawong, A.; Pasachan, T.; Trisuwan, K.; Soodvilai, S.; Duangjai, A.; Pongchaidecha, A.; Amornlerdpison, D.; Srimaroeng, C. Coffea arabica pulp aqueous extract attenuates oxidative stress and hepatic lipid accumulation in HepG2 cells. J. Herb. Med. 2021, 29, 100465. [Google Scholar] [CrossRef]
- Srinivas, N. Prashanthkumar Goudappala and Kashinath R T, Significance of plasma total cholesterol-VLDL cholesterol ratio (TC/VLDLC) in Type-2 diabetes mellitus. Int. J. Biomed. Res. 2018, 9, 36–39. [Google Scholar]
- Hou, X.; Zhang, C.; Wang, L.; Wang, K. Natural Piperine Improves Lipid Metabolic Profile of High-Fat Diet-Fed Mice by Upregulating SR-B1 and ABCG8 Transporters. J. Nat. Prod. 2021, 84, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Dong, Y.; Hou, Q.; Lei, J.; Wolf, P.G.; Ayansola, H.; Zhang, B. Quercetin Alleviates Intestinal Oxidative Damage Induced by H2O2 via Modulation of GSH: In Vitro Screening and In Vivo Evaluation in a Colitis Model of Mice. ACS Omega 2020, 5, 8334–8346. [Google Scholar] [CrossRef] [Green Version]
- Kovacs-Nolan, J.; Rupa, P.; Matsui, T.; Tanaka, M.; Konishi, T.; Sauchi, Y.; Sato, K.; Ono, S.; Mine, Y. In vitro and ex vivo uptake of glutathione (GSH) across the intestinal epithelium and fate of oral GSH after in vivo supplementation. J. Agric. Food Chem. 2014, 62, 9499–9506. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Firdaus, F.; Zafeer, M.F.; Anis, E.; Ahmad, M.; Afzal, M. Ellagic acid attenuates arsenic induced neuro-inflammation and mitochondrial dysfunction associated apoptosis. Toxicol. Rep. 2018, 5, 411–417. [Google Scholar] [CrossRef]
- Malota, K.; Student, S.; Swiatek, P. Low mitochondrial activity within developing earthworm male germ-line cysts revealed by JC-1. Mitochondrion 2019, 44, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Feng, M.; Zhang, S.; Dong, Z.; Wang, Y.; Zhang, W.; Liu, C. Angptl8 mediates food-driven resetting of hepatic circadian clock in mice. Nat. Commun. 2019, 10, 3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Wang, Z.; Zhang, T.; Chen, X.; Xu, H.; Wang, F.; Guo, L.; Chen, M.; Liu, K.; Wu, B. Deficiency of intestinal Bmal1 prevents obesity induced by high-fat feeding. Nat. Commun. 2021, 12, 5323. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, I.; Leinweber, B.; Brandenburger, M.; Oster, H. Circadian clock network desynchrony promotes weight gain and alters glucose homeostasis in mice. Mol. Metab. 2019, 30, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, S.Y.; Wu, G.; Altman, B.J.; Growe, J.; Rhoades, S.D.; Coldren, F.; Venkataraman, A.; Olarerin-George, A.O.; Francey, L.J.; Mukherjee, S.; et al. Clock Regulation of Metabolites Reveals Coupling between Transcription and Metabolism. Cell Metab. 2017, 25, 961–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corella, D.; Asensio, E.M.; Coltell, O.; Sorli, J.V.; Estruch, R.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Castaner, O.; Aros, F.; Lapetra, J.; et al. CLOCK gene variation is associated with incidence of type-2 diabetes and cardiovascular diseases in type-2 diabetic subjects: Dietary modulation in the PREDIMED randomized trial. Cardiovasc. Diabetol. 2016, 15, 4. [Google Scholar] [CrossRef] [Green Version]
- Barnea, M.; Chapnik, N.; Genzer, Y.; Froy, O. The circadian clock machinery controls adiponectin expression. Mol. Cell. Endocrinol. 2015, 399, 284–287. [Google Scholar] [CrossRef]
- Onder, Y.; Green, C.B. Rhythms of metabolism in adipose tissue and mitochondria. Neurobiol. Sleep Circadian Rhythm. 2018, 4, 57–63. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W. Role of mTOR in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2018, 19, 2043. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Nam, M.; Kang, M.S.; Lee, J.O.; Lee, Y.W.; Hwang, G.S.; Kim, H.S. Piperine regulates UCP1 through the AMPK pathway by generating intracellular lactate production in muscle cells. Sci. Rep. 2017, 7, 41066. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Ramanathan, C.; Kathale, N.D.; Liu, D.; Lee, C.; Freeman, D.A.; Hogenesch, J.B.; Cao, R.; Liu, A.C. mTOR signaling regulates central and peripheral circadian clock function. PLoS Genet. 2018, 14, e1007369. [Google Scholar]
- Maeda, A.; Shirao, T.; Shirasaya, D.; Yoshioka, Y.; Yamashita, Y.; Akagawa, M.; Ashida, H. Piperine promotes glucose uptake through ROS-dependent activation of the CAMKK/AMPK signaling pathway in skeletal muscle. Mol. Nutr. Food Res. 2018, 62, e1800086. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Miyata, S.; Shimizu, M.; Inoue, J.; Sato, R. Piperine induces hepatic low-density lipoprotein receptor expression through proteolytic activation of sterol regulatory element-binding proteins. PLoS ONE 2015, 10, e0139799. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Bmal1 | ATGGGGCTGGATGAAGACAA | CTGTTGCCCTCTGGTCTACA |
| Clock | ACGACGAGAACTTGGCATTG | GGTGTTGAGGAAGGGTCTGA |
| Per1 | AAGTCCGTCTTCTGCCGTAT | TATCCGGGGAGCTTCGTAAC |
| Per2 | AGCCGGAGTTAGAGATGGTG | TCTGCTCCTCCTTCTGTGTG |
| Cry1 | GTCTACATCCTGGACCCCTG | CTGGGAAACACATCTGCTGG |
| Cry2 | GGGAGGAGAGACAGAAGCTC | AATAGGGAGAGGGGAGGTGT |
| Reverbα | CTGGGAGGATTTCTCCATGA | TTCACGTTGAACAACGAAGC |
| Srebp-1c | CTGAGGCAAAGCTGAATAAATCTGCTG | GTTCTCCTGCTTGAGTTTCTGGTTG |
| Pparγ | TGCTGTTATGGGTGAAACTCTG | CTGTGTCAACCATGGTAATTTCTT |
| Cebpb | ACGAGCGCGCCATCGACTTC | GAAGCCCGGCTCCGCCTTG |
| Adipoq | CCTGAACCCTACAAGCGATG | GGTTCCACTTCTTTGTCCTCG |
| Fas | TGCCCAAGTGACTGACATCA | CATCCCCATTGACTGTGCAG |
| Acc | CAACTTTGTGCCCACGGTTA | TTTGTCAGGAAGAGGCGGAT |
| Akt | CTGCCCTTCTACAACCAGGA | ATGATCTCCTTGGCGTCCTC |
| mTOR | CCTGCCTTTGTCATGCCTTT | CTGGGTTTGGATCAGGGTCT |
| Pgc-1α | AGCGCCGTGTGATTTATGTC | TGCGTCCACAAAAGTACAGC |
| Cpt-1 | CCTCCGTAGCTGACTCGGTA | GGAGTGACCGTGAACTGAAAG |
| Ampk | GACAAGCCCACCTGATTC | TTCCTTCGTACACGCAAA |
| GAPDH | TCAAGAAGGTGGTGAAGCAGG | TCAAAGGTGGAGGAGTGGGT |
| Primary Antibodies | Catalogue | Company | Dilution Rates |
|---|---|---|---|
| BMAL1 | 14268-1-AP | Proteintech | 1:600 |
| CLOCK | 18094-1-AP | Proteintech | 1:600 |
| SREBP-1C | 14088-1-AP | Proteintech | 1:1000 |
| PPARγ | 16643-1-AP | Proteintech | 1:1000 |
| C/EBPβ | 66649-1-IG | Proteintech | 1:2000 |
| FAS | 66369-1-Ig | Proteintech | 1:5000 |
| ACC | 67373-1-Ig | Proteintech | 1:5000 |
| p-mTOR | 67778-1-Ig | Proteintech | 1:1000 |
| mTOR | 10176-2-AP | Proteintech | 1:1000 |
| Adiponectin | 21613-1-AP | Proteintech | 1:1000 |
| CPT-1 | 15184-1-AP | Proteintech | 1:5000 |
| PGC-1α | 20658-1-AP | Proteintech | 1:1000 |
| p-AMPK | #2535 | CST | 1:1000 |
| AMPK | 66536-1-Ig | Proteintech | 1:1000 |
| p-AKT | 28731-1-AP | Proteintech | 1:1000 |
| AKT | 10176-1-AP | Proteintech | 1:1000 |
| GAPDH | ATPA00013Rb | Atagenix | 1:5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Ho, C.-T.; Lu, M. Piperine Improves Lipid Dysregulation by Modulating Circadian Genes Bmal1 and Clock in HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 5611. https://doi.org/10.3390/ijms23105611
Zhang W, Ho C-T, Lu M. Piperine Improves Lipid Dysregulation by Modulating Circadian Genes Bmal1 and Clock in HepG2 Cells. International Journal of Molecular Sciences. 2022; 23(10):5611. https://doi.org/10.3390/ijms23105611
Chicago/Turabian StyleZhang, Weiyun, Chi-Tang Ho, and Muwen Lu. 2022. "Piperine Improves Lipid Dysregulation by Modulating Circadian Genes Bmal1 and Clock in HepG2 Cells" International Journal of Molecular Sciences 23, no. 10: 5611. https://doi.org/10.3390/ijms23105611
APA StyleZhang, W., Ho, C.-T., & Lu, M. (2022). Piperine Improves Lipid Dysregulation by Modulating Circadian Genes Bmal1 and Clock in HepG2 Cells. International Journal of Molecular Sciences, 23(10), 5611. https://doi.org/10.3390/ijms23105611