
Overexpression of Sugarcane ScDIR Genes Enhances Drought Tolerance in Nicotiana benthamiana

 and
and 
Abstract
:1. Introduction
2. Results
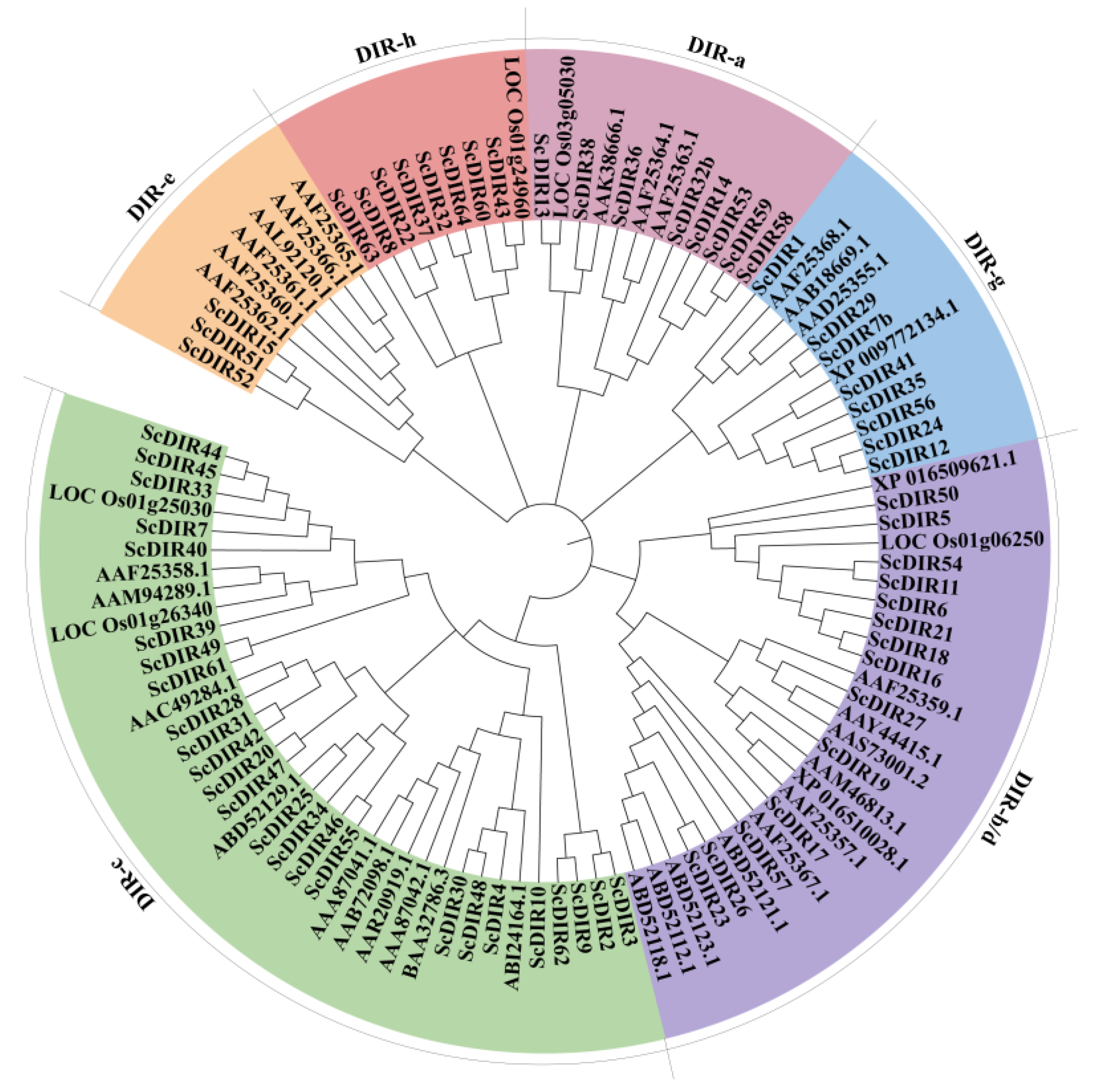
2.1. Characterization of Sugarcane ScDIR Proteins
2.2. Expression Pattern of Sugarcane ScDIR Genes under Drought Stress
2.3. Genetic Structures, Sequence Alignments, Motifs, and Tertiary Structures of the ScDIR5, ScDIR7, ScDIR11, and ScDIR40 Genes
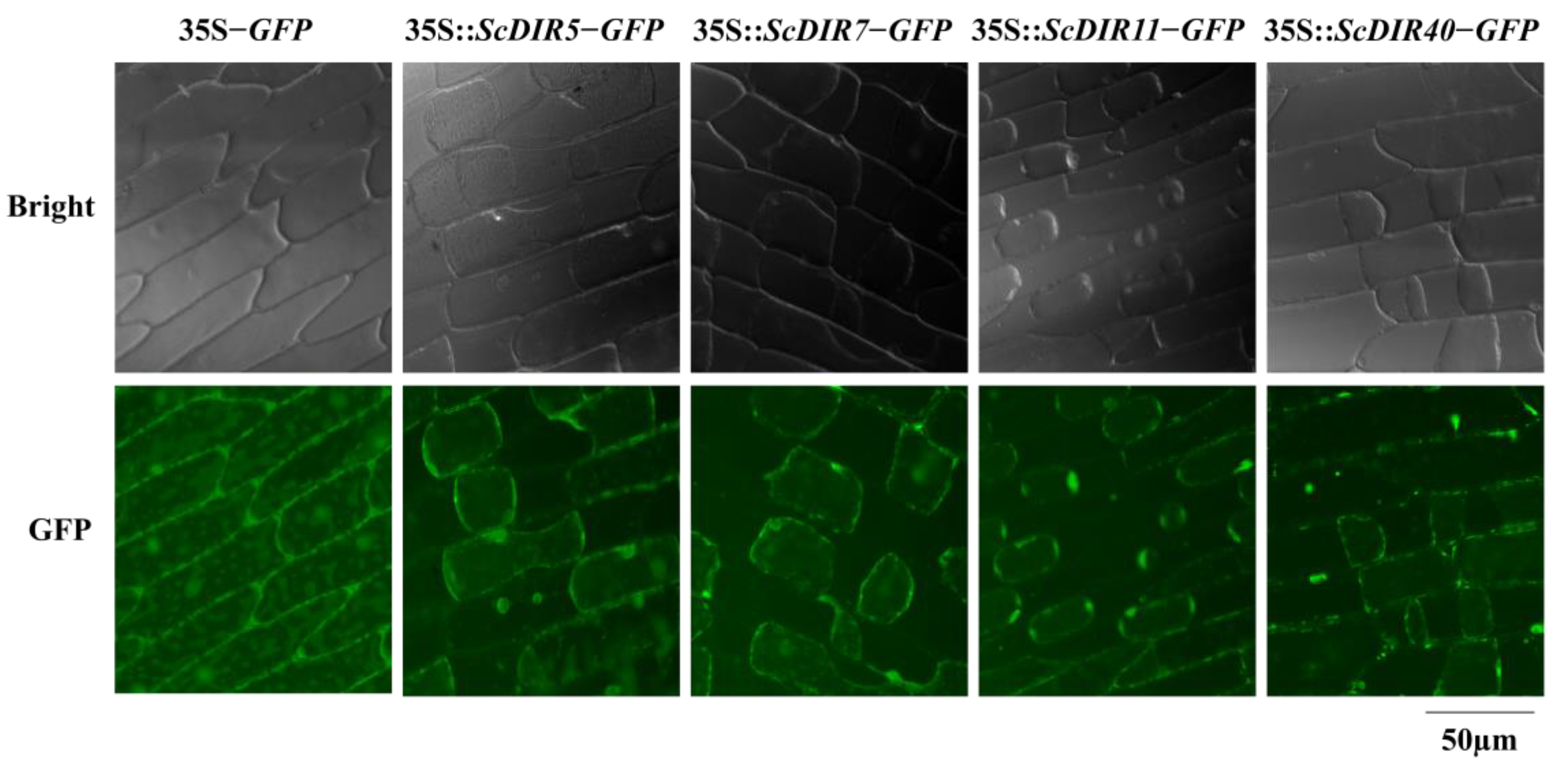
2.4. Subcellular Localization of ScDIRs
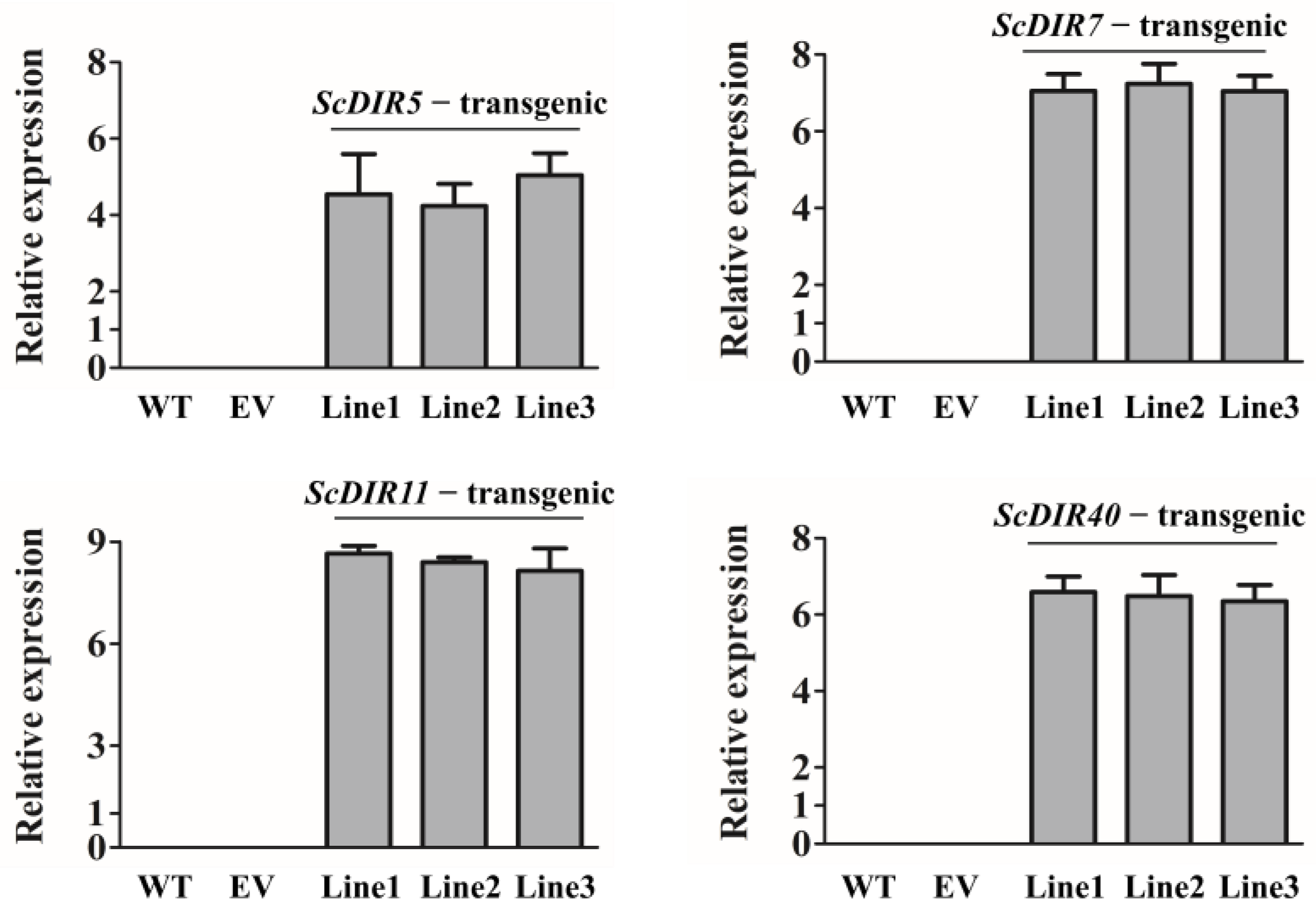
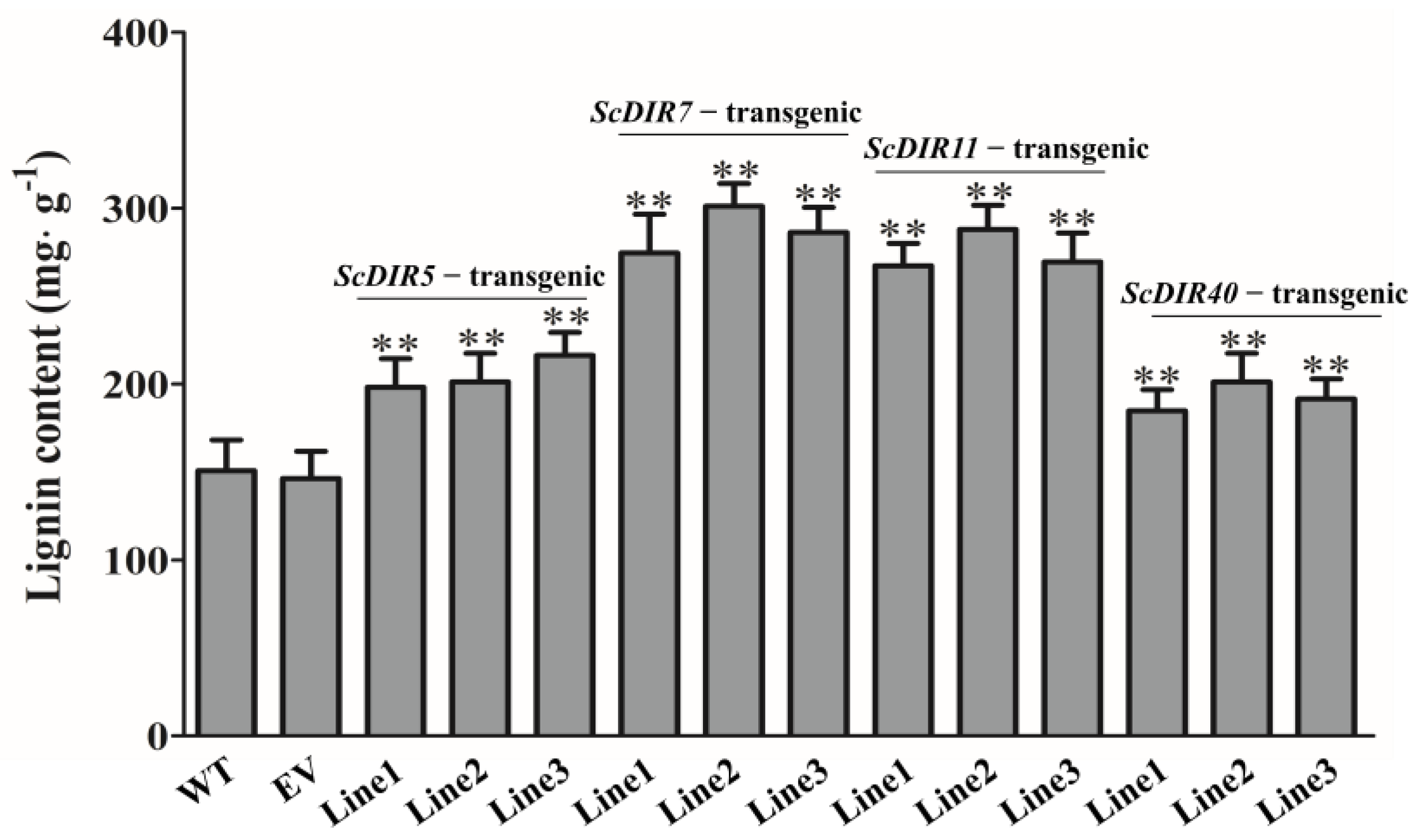
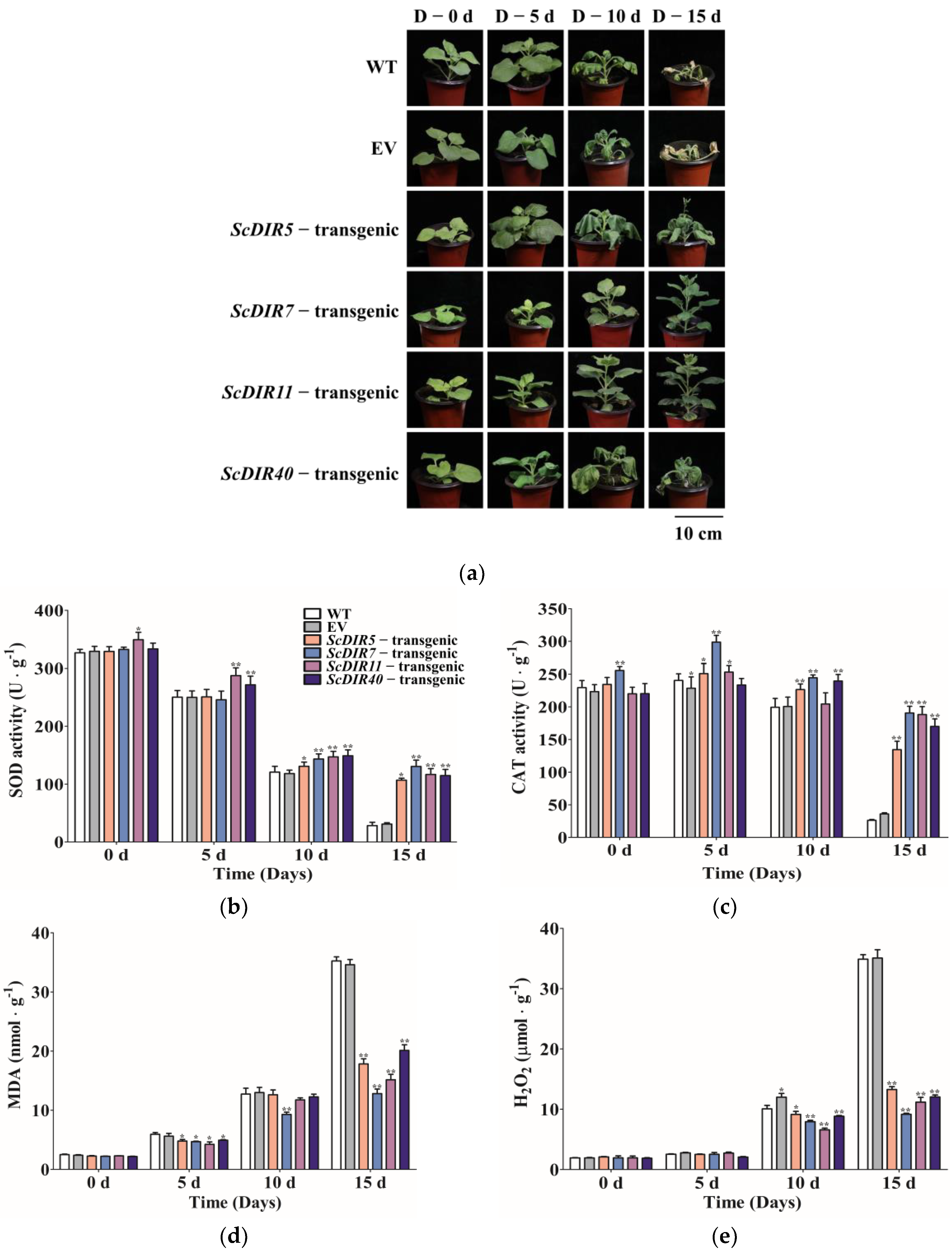
2.5. Overexpression of ScDIR Genes Enhances Tolerance to Drought Stress in Transgenic Tobacco
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Drought Stress Conditions
4.2. Bioinformatics Analysis of Sugarcane ScDIR Proteins
4.3. Gene Expression Quantification
4.4. Subcellular Localization of Proteins
4.5. Tobacco Transformation and Regeneration
4.6. Measurement of Physiological and Biochemical Indices
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marcos, F.C.C.; Silveira, N.M.; Mokochinski, J.B.; Sawaya, A.C.H.F.; Marchiori, P.E.R.; Machado, E.C.; Souza, G.M.; Landell, M.G.A.; Ribeiro, R.V. Drought Tolerance of Sugarcane Is Improved by Previous Exposure to Water Deficit. J. Plant Physiol. 2018, 223, 9–18. [Google Scholar] [CrossRef]
- Machado, D.F.S.P.; Lagôa, A.M.M.A.; Ribeiro, R.V.; Marchiori, P.E.R.; Machado, R.S.; Machado, E.C. Baixa Temperatura Noturna e Deficiência Hídrica Na Fotossíntese de Cana−de−Açúcar. Pesqui. Agropecu. Bras. 2013, 48, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, M.K.; Zingsheim, O.; El-Shourbagy, M.N.; Moore, P.H.; Komor, E. Growth and Sugar Storage in Sugarcane Grown at Temperatures below and above Optimum. J. Plant Physiol. 1998, 153, 593–602. [Google Scholar] [CrossRef]
- Davin, L.B.; Wang, H.-B.; Crowell, A.L.; Bedgar, D.L.; Martin, D.M.; Sarkanen, S.; Lewis, N.G. Stereoselective Bimolecular Phenoxy Radical Coupling by an Auxiliary (Dirigent) Protein without an Active Center. Science 1997, 275, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-S.; Yu, G.-L.; Zou, J.-N.; Wang, J.; Qiu, H.-M.; Zhu, R.-S.; Chang, H.-L.; Jiang, H.-W.; Hu, Z.-B.; Li, C.-Y.; et al. GmDRR1, a Dirigent Protein Resistant to Phytophthora Sojae in Glycine Max (L.) Merr. J. Integr. Agric. 2018, 17, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Ralph, S.; Park, J.-Y.; Bohlmann, J.; Mansfield, S.D. Dirigent Proteins in Conifer Defense: Gene Discovery, Phylogeny, and Differential Wound− and Insect−Induced Expression of a Family of DIR and DIR−like Genes in Spruce (Picea Spp.). Plant Mol. Biol. 2006, 60, 21–40. [Google Scholar] [CrossRef]
- Ralph, S.G.; Jancsik, S.; Bohlmann, J. Dirigent Proteins in Conifer Defense II: Extended Gene Discovery, Phylogeny, and Constitutive and Stress−Induced Gene Expression in Spruce (Picea Spp.). Phytochemistry 2007, 68, 1975–1991. [Google Scholar] [CrossRef]
- Liao, Y.; Liu, S.; Jiang, Y.; Hu, C.; Zhang, X.; Cao, X.; Xu, Z.; Gao, X.; Li, L.; Zhu, J.; et al. Genome−Wide Analysis and Environmental Response Profiling of Dirigent Family Genes in Rice (Oryza Sativa). Genes Genom. 2017, 39, 47–62. [Google Scholar] [CrossRef]
- Nobile, P.M.; Bottcher, A.; Mayer, J.L.S.; Brito, M.S.; dos Anjos, I.A.; Landell, M.G.A.; Vicentini, R.; Creste, S.; Riaño-Pachón, D.M.; Mazzafera, P. Identification, Classification and Transcriptional Profiles of Dirigent Domain−Containing Proteins in Sugarcane. Mol. Genet. Genom. 2017, 292, 1323–1340. [Google Scholar] [CrossRef]
- Cheng, X.; Su, X.; Muhammad, A.; Li, M.; Zhang, J.; Sun, Y.; Li, G.; Jin, Q.; Cai, Y.; Lin, Y. Molecular Characterization, Evolution, and Expression Profiling of the Dirigent (DIR) Family Genes in Chinese White Pear (Pyrus Bretschneideri). Front. Genet. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Gasper, R.; Effenberger, I.; Kolesinski, P.; Terlecka, B.; Hofmann, E.; Schaller, A. Dirigent Protein Mode of Action Revealed by the Crystal Structure of AtDIR6. Plant Physiol. 2016, 172, 2165–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Wang, L.; Wang, Z.; Shang, H.; Liu, X.; Zhu, Y.; Qi, D.; Deng, X. Cloning and Expression Analysis of a Dirigent Protein Gene from the Resurrection Plant Boea Hygrometrica. Prog. Nat. Sci. 2009, 19, 347–352. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a Tool to Investigate Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, X.; Yang, R.; Gao, B.; Yao, J.; Oliver, M.J.; Zhang, D. BaDBL1, a Unique DREB Gene from Desiccation Tolerant Moss Bryum Argenteum, Confers Osmotic and Salt Stress Tolerances in Transgenic Arabidopsis. Plant Sci. 2021, 313, 111047. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, M.Q.; Lin, Y.Q.; Zhang, H.; Chen, R.K. Studies on the Relationship of Chlorophyll FLuorescence Characters and Drought Tolerance in Seedling of Sugarcane under Water Stress. Sci. Agric. Sin. 2004, 37, 1718–1721. [Google Scholar]
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; et al. TaASR1, a Transcription Factor Gene in Wheat, Confers Drought Stress Tolerance in Transgenic Tobacco. Plant. Cell Environ. 2013, 36, 1449–1464. [Google Scholar] [CrossRef]
- Shi, H.; Liu, Z.; Zhu, L.; Zhang, C.; Chen, Y.; Zhou, Y.; Li, F.; Li, X. Overexpression of Cotton (Gossypium Hirsutum) Dirigent1 Gene Enhances Lignification That Blocks the Spread of Verticillium Dahliae. Acta Biochim. Biophys. Sin. 2012, 44, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.-L.; Xu, L.-P.; Fang, J.-P.; Su, Y.-C.; Fu, H.-Y.; Que, Y.-X.; Xu, J.-S. A Novel Dirigent Protein Gene with Highly Stem−Specific Expression from Sugarcane, Response to Drought, Salt and Oxidative Stresses. Plant Cell Rep. 2012, 31, 1801–1812. [Google Scholar] [CrossRef]
- Zhang, F.-J.; Zhang, K.-K.; Du, C.-Z.; Li, J.; Xing, Y.-X.; Yang, L.-T.; Li, Y.-R. Effect of Drought Stress on Anatomical Structure and Chloroplast Ultrastructure in Leaves of Sugarcane. Sugar Tech. 2015, 17, 41–48. [Google Scholar] [CrossRef]
- Casu, R.E.; Dimmock, C.M.; Chapman, S.C.; Grof, C.P.L.; McIntyre, C.L.; Bonnett, G.D.; Manners, J.M. Identification of Differentially Expressed Transcripts from Maturing Stem of Sugarcane by in Silico Analysis of Stem Expressed Sequence Tags and Gene Expression Profiling. Plant Mol. Biol. 2004, 54, 503–517. [Google Scholar] [CrossRef]
- Damaj, M.B.; Kumpatla, S.P.; Emani, C.; Beremand, P.D.; Reddy, A.S.; Rathore, K.S.; Buenrostro-Nava, M.T.; Curtis, I.S.; Thomas, T.L.; Mirkov, E.E. Sugarcane DIRIGENT and O−METHYLTRANSFERASE Promoters Confer Stem−Regulated Gene Expression in Diverse Monocots. Planta 2010, 231, 1439–1458. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Peng, X. Genome−Wide Identification and Characterization of DIR Genes in Medicago Truncatula. Biochem. Genet. 2019, 57, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhong, S.; Zhang, Q.; Ren, Y.; Sun, C.; Chen, F. A Loss−of−Function of the Dirigent Gene TaDIR−B1 Improves Resistance to Fusarium Crown Rot in Wheat. Plant Biotechnol. J. 2021, 19, 866–868. [Google Scholar] [CrossRef]
- Li, S.S.; Chang, Y.; Li, B.; Shao, S.L.; Zhen, Z.-Z. Functional Analysis of 4−Coumarate: Coa Ligase from Dryopteris Fragrans in Transgenic Tobacco Enhances Lignin and Flavonoids. Genet. Mol. Biol. 2020, 43, e20180355. [Google Scholar] [CrossRef]
- Li, Z.; Tang, M.; Hassan, M.J.; Zhang, Y.; Han, L.; Peng, Y. Adaptability to High Temperature and Stay−Green Genotypes Associated With Variations in Antioxidant, Chlorophyll Metabolism, and γ−Aminobutyric Acid Accumulation in Creeping Bentgrass Species. Front. Plant Sci. 2021, 12, 750728. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Jin, Y.; Li, H.; Amombo, E.; Fu, J. Stress Memory Induced Transcriptional and Metabolic Changes of Perennial Ryegrass (Lolium Perenne) in Response to Salt Stress. Physiol. Plant. 2015, 156, 54–69. [Google Scholar] [CrossRef]
- Silva, K.I.; Sales, C.R.G.; Marchiori, P.E.R.; Silveira, N.M.; Machado, E.C.; Ribeiro, R.V. Short−Term Physiological Changes in Roots and Leaves of Sugarcane Varieties Exposed to H2O2 in Root Medium. J. Plant Physiol. 2015, 177, 93–99. [Google Scholar] [CrossRef]
- Ahmad, P.; Alam, P.; Balawi, T.H.; Altalayan, F.H.; Ahanger, M.A.; Ashraf, M. Sodium Nitroprusside (SNP) Improves Tolerance to Arsenic (As) Toxicity in Vicia Faba through the Modifications of Biochemical Attributes, Antioxidants, Ascorbate−Glutathione Cycle and Glyoxalase Cycle. Chemosphere 2020, 244, 125480. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real−Time Quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef]
- Sun, W.; Cao, Z.; Li, Y.; Zhao, Y.; Zhang, H. A Simple and Effective Method for Protein Subcellular Localization Using Agrobacterium−mediated Transformation of Onion Epidermal Cells. Biologia 2007, 62, 529–532. [Google Scholar] [CrossRef]
- Lin, J.J.; Assad−Garcia, N.; Kuo, J. Plant Hormone Effect of Antibiotics on the Transformation Efficiency of Plant Tissues by Agrobacterium Tumefaciens Cells. Plant Sci. 1995, 109, 171–177. [Google Scholar] [CrossRef]
- Su, Y.; Xu, L.; Wang, Z.; Peng, Q.; Yang, Y.; Chen, Y.; Que, Y. Comparative Proteomics Reveals That Central Metabolism Changes Are Associated with Resistance against Sporisorium Scitamineum in Sugarcane. BMC Genom. 2016, 17, 800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.Q.; Yang, L.T.; Li, Y.R. Physiological and Biochemical Characteristics Related to Cold Resistance in Sugarcane. Sugar Tech. 2015, 17, 49–58. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Subordinate Function Values * | Integrated Assessment | Order of Resistance | |
|---|---|---|---|---|
| U1 | U2 | Value D | ||
| Wild type | 0.932 | 0.194 | 0.868 | 5 |
| EV | 1.024 | 0.191 | 0.834 | 6 |
| ScDIR5−transgenic | 1.258 | 0.228 | 1.178 | 4 |
| ScDIR7−transgenic | 1.689 | 0.277 | 1.587 | 1 |
| ScDIR11−transgenic | 1.521 | 0.261 | 1.428 | 2 |
| ScDIR40−transgenic | 1.272 | 0.198 | 1.202 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liu, Z.; Zhao, H.; Deng, X.; Su, Y.; Li, R.; Chen, B. Overexpression of Sugarcane ScDIR Genes Enhances Drought Tolerance in Nicotiana benthamiana. Int. J. Mol. Sci. 2022, 23, 5340. https://doi.org/10.3390/ijms23105340
Li X, Liu Z, Zhao H, Deng X, Su Y, Li R, Chen B. Overexpression of Sugarcane ScDIR Genes Enhances Drought Tolerance in Nicotiana benthamiana. International Journal of Molecular Sciences. 2022; 23(10):5340. https://doi.org/10.3390/ijms23105340
Chicago/Turabian StyleLi, Xiufang, Zongling Liu, Haiyun Zhao, Xingli Deng, Yizu Su, Ru Li, and Baoshan Chen. 2022. "Overexpression of Sugarcane ScDIR Genes Enhances Drought Tolerance in Nicotiana benthamiana" International Journal of Molecular Sciences 23, no. 10: 5340. https://doi.org/10.3390/ijms23105340
APA StyleLi, X., Liu, Z., Zhao, H., Deng, X., Su, Y., Li, R., & Chen, B. (2022). Overexpression of Sugarcane ScDIR Genes Enhances Drought Tolerance in Nicotiana benthamiana. International Journal of Molecular Sciences, 23(10), 5340. https://doi.org/10.3390/ijms23105340