
Actions and Roles of FSH in Germinative Cells

 , , , , , , , and
, , , , , , , and
Abstract
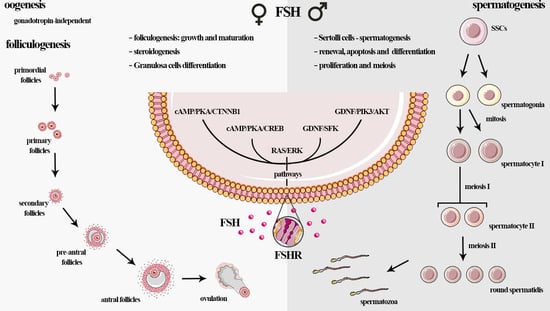
1. Introduction
2. The Role of FSH in Reproductive Development: Fetal and Neonatal Periods
3. FSH in the Adult Phase: Folliculogenesis and Spermatogenesis
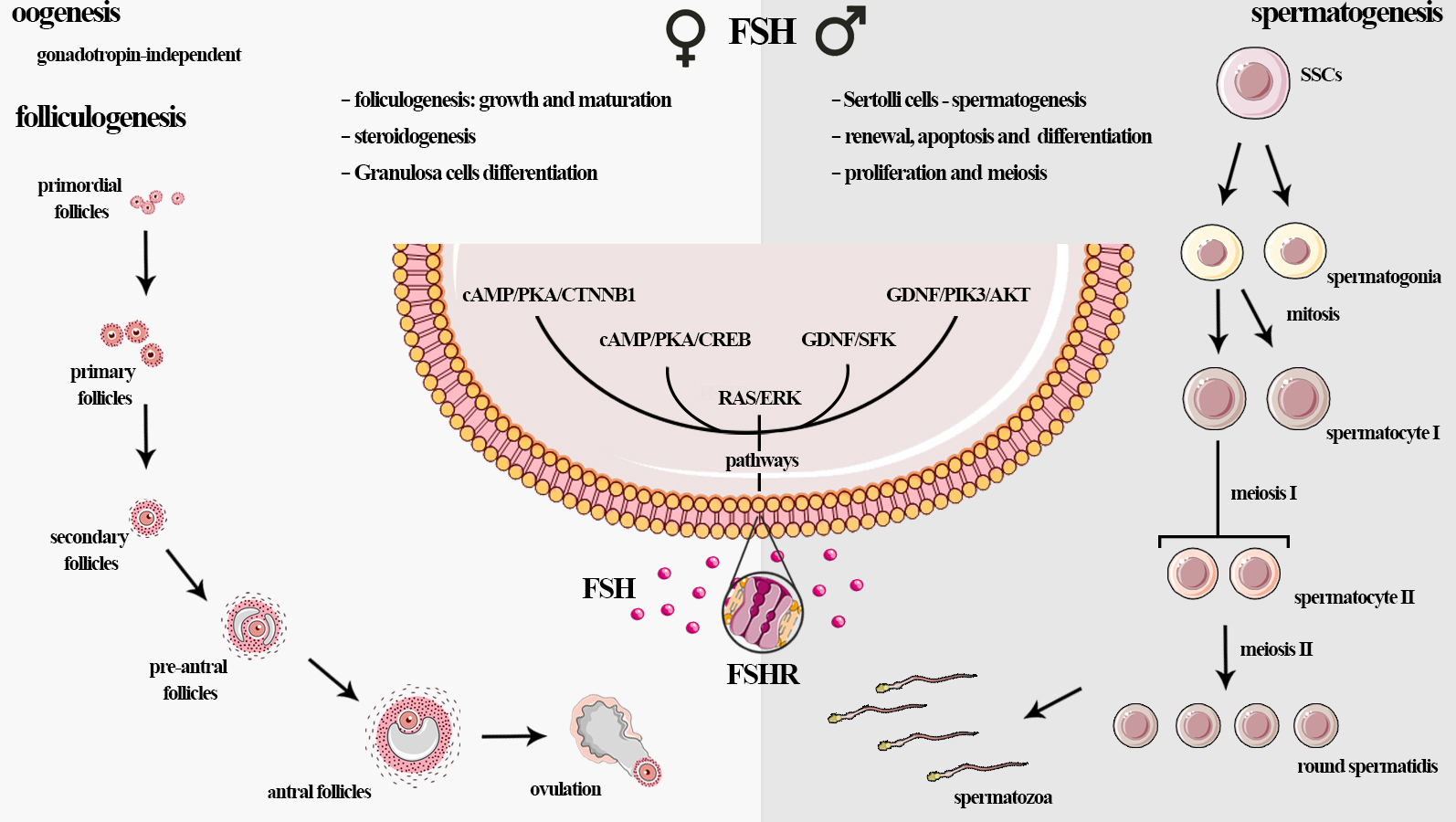
3.1. Females: FSH Influences Folliculogenesis
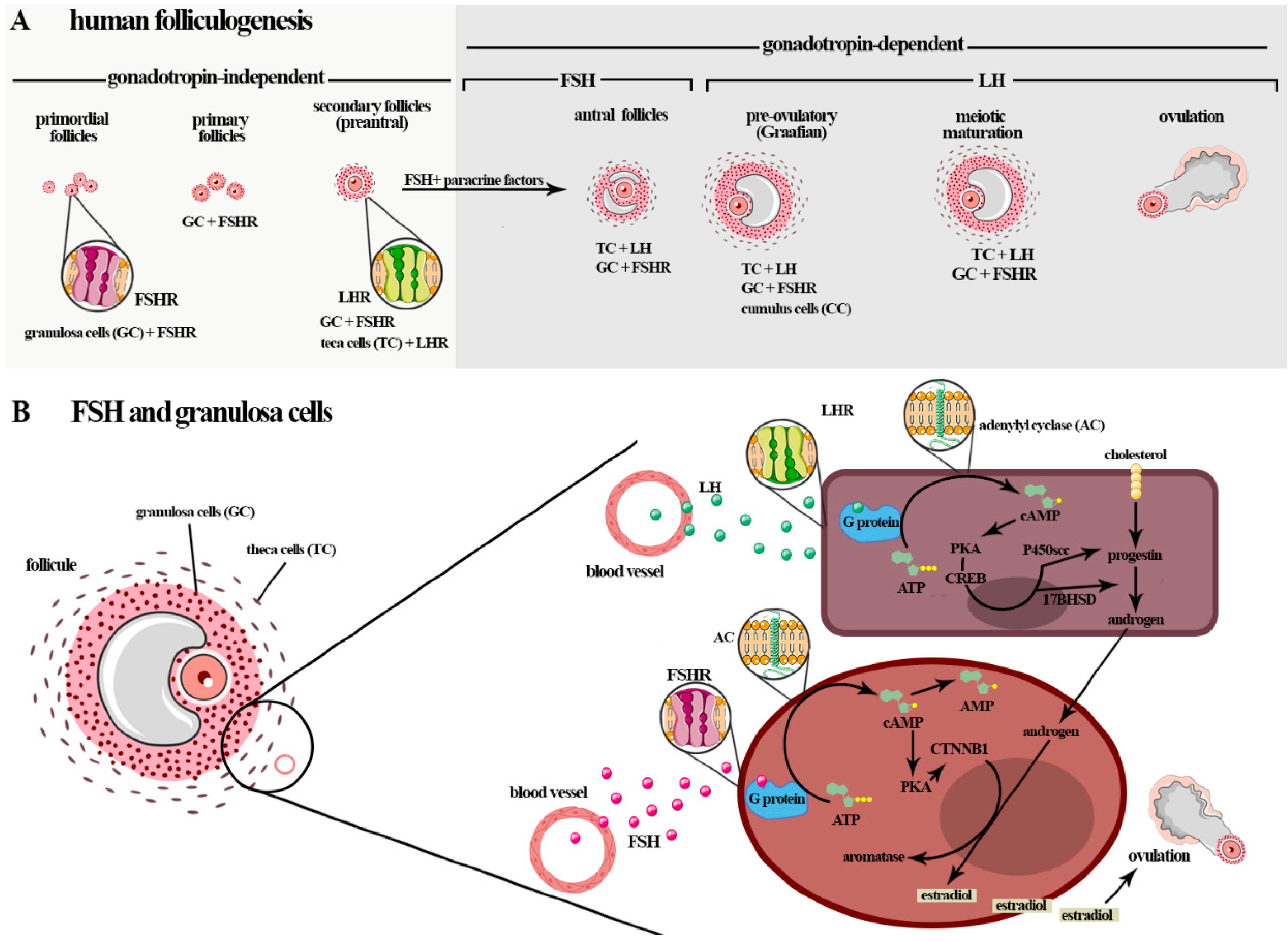
3.2. Males-Influence of FSH in Spermatogenesis
4. Influence of FSH on Germ Cells
4.1. Females: Granulosa Cells
4.2. Males: Sertoli Cells and Spermatogonial Stem Cells (SSCs)
5. Influence of FSH in Germ Cells In Vitro
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oduwole, O.O.; Peltoketo, H.; Huhtaniemi, I.T. Role of follicle-stimulating hormone in spermatogenesis. Front. Endocrinol. 2018, 9, 763. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Q.; Zhang, W.-D.; Yuan, B.; Zhang, J.-B. Advances in the Regulation of Mammalian Follicle-Stimulating Hormone Secretion. Animals 2021, 11, 1134. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, A.; Kim, B.; Yeh, J. Luteinizing Hormone Action in Human Oocyte Maturation and Quality: Signaling Pathways, Regulation, and Clinical Impact. Reprod. Sci. 2020, 27, 1223–1252. [Google Scholar] [CrossRef]
- Padmanabhan, V.; Cardoso, R.C. Neuroendocrine, autocrine, and paracrine control of follicle-stimulating hormone secretion. Mol. Cell. Endocrinol. 2020, 500, 110632. [Google Scholar] [CrossRef]
- Patel, H.; Bhartiya, D. Testicular Stem Cells Express Follicle-Stimulating Hormone Receptors and Are Directly Modulated by FSH. Reprod. Sci. 2016, 23, 1493–1508. [Google Scholar] [CrossRef]
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330. [Google Scholar] [CrossRef]
- Rossi, P.; Dolci, S. Paracrine mechanisms involved in the control of early stages of mammalian spermatogenesis. Front. Endocrinol. 2013, 4, 181. [Google Scholar] [CrossRef]
- Choi, S.G.; Jia, J.; Pfeffer, R.L.; Sealfon, S.C. G proteins and autocrine signaling differentially regulate gonadotropin subunit expression in pituitary gonadotrope. J. Biol. Chem. 2012, 287, 21550–21560. [Google Scholar] [CrossRef]
- Duan, H.; Xiao, L.; Ge, W.; Yang, S.; Jiang, Y.; Lv, J.; Hu, J.; Zhang, Y.; Zhao, X.; Hua, Y. Follicle-stimulating hormone and luteinizing hormone regulate the synthesis mechanism of dihydrotestosterone in sheep granulosa cells. Reprod. Domest. Anim. 2021, 56, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Casarini, L.; Lazzaretti, C.; Paradiso, E.; Limoncella, S.; Riccetti, L.; Sperduti, S.; Melli, B.; Marcozzi, S.; Anzivino, C.; Sayers, N.S.; et al. Membrane Estrogen Receptor (GPER) and Follicle-Stimulating Hormone Receptor (FSHR) Heteromeric Complexes Promote Human Ovarian Follicle Survival. iScience 2020, 23, 101812. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Kumar, T.R. Molecular regulation of follicle-stimulating hormone synthesis, secretion and action. J. Mol. Endocrinol. 2018, 60, R131–R155. [Google Scholar] [CrossRef]
- Siegel, E.T.; Kim, H.G.; Nishimoto, H.K.; Layman, L.C. The molecular basis of impaired follicle-stimulating hormone action: Evidence from human mutations and mouse models. Reprod. Sci. 2013, 20, 211–233. [Google Scholar] [CrossRef]
- Szymańska, K.; Kałafut, J.; Przybyszewska, A.; Paziewska, B.; Adamczuk, G.; Kiełbus, M.; Rivero-Müller, A. FSHR trans-activation and oligomerization. Front. Endocrinol. 2018, 9, 760. [Google Scholar] [CrossRef]
- Simoni, M.; Gromoll, J.; Nieschlag, E. The follicle-stimulating hormone receptor: Biochemistry, molecular biology, physiology, and pathophysiology. Endocr. Rev. 1997, 18, 739–773. [Google Scholar] [CrossRef] [PubMed]
- Jonas, K.C.; Rivero-Müller, A.; Huhtaniemi, I.T.; Hanyaloglu, A.C. G Protein-Coupled Receptor Transactivation. From Molecules to Mice. Methods Cell Biol. 2013, 117, 433–450. [Google Scholar]
- Rivero-Müller, A.; Chou, Y.Y.; Ji, I.; Lajic, S.; Hanyaloglu, A.C.; Jonas, K.; Rahman, N.; Ji, T.H.; Huhtaniemi, I. Rescue of defective G protein—Coupled receptor function in vivo by intermolecular cooperation. Proc. Natl. Acad. Sci. USA 2010, 107, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Bonfil, D.; Chuderland, D.; Kraus, S.; Shahbazian, D.; Friedberg, I.; Seger, R.; Naor, Z. Extracellular signal-regulated kinase, Jun N-terminal kinase, p38, and c-Src are involved in gonadotropin-releasing hormone-stimulated activity of the glycoprotein hormone follicle-stimulating hormone beta-subunit promoter. Endocrinology 2004, 145, 2228–2244. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ruiz, M.S.; Austin, D.A.; Webster, N.J. Constitutively active Gq impairs gonadotropin-releasing hormone-induced intracellular signaling and luteinizing hormone secretion in LbetaT2 cells. Mol. Endocrinol. 2005, 19, 2074–2085. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Choi, S.; Ruf-Zamojski, F.; Pincas, H.; Roysam, B.; Sealfon, S.C. Characterization of a MAPK scaffolding protein logic gate in gonadotropes. Mol. Endocrinol. 2011, 25, 1027–1039. [Google Scholar] [CrossRef]
- James, K.; Bhartiya, D.; Ganguly, R.; Kaushik, A.; Gala, K.; Singh, P.; Metkari, S.M. Gonadotropin and steroid hormones regulate pluripotent very small embryonic-like stem cells in adult mouse uterine endometrium. J. Ovarian Res. 2018, 11, 1–20. [Google Scholar] [CrossRef]
- Stilley, J.A.; Guan, R.; Duffy, D.M.; Segaloff, D.L. Signaling through FSH receptors on human umbilical vein endothelial cells promotes angiogenesis. J. Clin. Endocrinol. Metab. 2014, 99, E813–E820. [Google Scholar] [CrossRef]
- Chrusciel, M.; Ponikwicka-Tyszko, D.; Wolczynski, S.; Huhtaniemi, I.; Rahman, N.A. Extragonadal FSHR expression and function—Is it real? Front. Endocrinol. 2019, 10, 32. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Ratajczak, J.; Kucia, M. Very Small Embryonic-Like Stem Cells (VSELs): An Update and Future Directions. Circ. Res. 2019, 124, 208–210. [Google Scholar] [CrossRef]
- Bhartiya, D.; Shaikh, A.; Anand, S.; Patel, H.; Kapoor, S.; Sriraman, K.; Parte, S.; Unni, S. Endogenous, very small embryonic-like stem cells: Critical review, therapeutic potential and a look ahead. Hum. Reprod. Update 2016, 23, 1–36. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Zuba-surma, E.K.; Ratajczak, J.; Wysoczynski, M. Very Small Embryonic Like (VSEL) Stem Cells—Characterization, Developmental Origin and Biological Significance. Exp. Hematol. 2009, 36, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Bhartiya, D. Direct action of FSH on testicular stem cells. Stem Cell Res. Ther. 2019, 10, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Siraj, A.; Desestret, V.; Antoine, M.; Fromont, G.; Huerre, M.; Sanson, M.; Camparo, P.; Pichon, C.; Planeix, F.; Gonin, J.; et al. Expression of follicle-stimulating hormone receptor by the vascular endothelium in tumor metastases. BMC Cancer 2013, 13, 246. [Google Scholar] [CrossRef] [PubMed]
- Planeix, F.; Siraj, M.A.; Bidard, F.C.; Robin, B.; Pichon, C.; Sastre-Garau, X.; Antoine, M.; Ghinea, N. Endothelial follicle-stimulating hormone receptor expression in invasive breast cancer and vascular remodeling at tumor periphery. J. Exp. Clin. Cancer Res. 2015, 34, 12. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.J.; Tourkova, I.; Wang, Y.; Sharrow, A.C.; Landau, M.S.; Yaroslavskiy, B.B.; Sun, L.; Zaidi, M.; Blair, H.C. FSH-receptor isoforms and FSH-dependent gene transcription in human monocytes and osteoclasts. Biochem. Biophys. Res. Commun. 2010, 394, 12–17. [Google Scholar] [CrossRef]
- Casarini, L.; Simoni, M. Recent advances in understanding gonadotropin signaling. Fac. Rev. 2021, 10. [Google Scholar] [CrossRef]
- Stilley, J.A.W.; Christensen, D.E.; Dahlem, K.B.; Guan, R.; Santillan, D.A.; England, S.K.; Al-Hendy, A.; Kirby, P.A.; Segaloff, D.L. FSH receptor (FSHR) expression in human extragonadal reproductive tissues and the developing placenta, and the impact of its deletion on pregnancy in mice. Biol. Reprod. 2014, 91, 74. [Google Scholar] [CrossRef]
- Song, Y.; Wang, E.S.; Xing, L.L.; Shi, S.; Qu, F.; Zhang, D.; Li, J.Y.; Shu, J.; Meng, Y.; Sheng, J.Z.; et al. Follicle-stimulating hormone induces postmenopausal dyslipidemia through inhibiting hepatic cholesterol metabolism. J. Clin. Endocrinol. Metab. 2016, 101, 254–263. [Google Scholar] [CrossRef]
- Stelmaszewska, J.; Chrusciel, M.; Doroszko, M.; Akerfelt, M.; Ponikwicka-Tyszko, D.; Nees, M.; Frentsch, M.; Li, X.; Kero, J.; Huhtaniemi, I.; et al. Revisiting the expression and function of follicle-stimulation hormone receptor in human umbilical vein endothelial cells. Sci. Rep. 2016, 6, 37095. [Google Scholar] [CrossRef]
- Rougier, C.; Hieronimus, S.; Panaïa-Ferrari, P.; Lahlou, N.; Paris, F.; Fenichel, P. Isolated follicle-stimulating hormone (FSH) deficiency in two infertile men without FSH β gene mutation: Case report and literature review. Ann. Endocrinol. Paris 2019, 80, 234–239. [Google Scholar] [CrossRef]
- Hamilton-fairley, D.; Kiddy, D.; Watson, H.; Sagle, M.; Franks, S. Low-dose gonadotrophin therapy for induction of ovulation in 100 women with polycystic ovary syndrome. Hum. Reprod. 1991, 6, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Karakas, S.E.; Surampudi, P. New Biomarkers to Evaluate Hyperandrogenemic Women and Hypogonadal Men. Adv. Clin. Chem. 2018, 86, 71–125. [Google Scholar] [CrossRef] [PubMed]
- Fraietta, R.; Zylberstejn, D.S.; Esteves, S.C. Hypogonadotropic Hypogonadism Revisited. Clinics 2013, 68, 81–88. [Google Scholar] [CrossRef]
- Lanfranco, F.; Kamischke, A.; Zitzmann, M.; Nieschlag, E. Klinefelter’s syndrome. Lancet Lond. Engl. 2004, 364, 273–283. [Google Scholar] [CrossRef]
- Groth, K.A.; Skakkebaek, A.; Høst, C.; Gravholt, C.H.; Bojesen, A. Klinefelter Syndrome-A Clinical Update Diagnosis, Epidemiology, and Genetics. J. Clin. Endocrinol. Metab. 2013, 98, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Behre, H.M. Clinical Use of FSH in Male Infertility. Front. Endocrinol. 2019, 10, 322. [Google Scholar] [CrossRef]
- Bhasin, S.; Cunningham, G.R.; Hayes, F.J.; Matsumoto, A.M.; Snyder, P.J.; Swerdloff, R.S.; Montori, V.M. Testosterone therapy in men with androgen deficiency syndromes: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2010, 95, 2536–2559. [Google Scholar] [CrossRef]
- Mao, J.; Liu, X.; Nie, M.; Wang, X.; Xu, L.; Huang, B.; Zheng, J.; Min, L.; Kaiser, I.; Wu, X. Pulsatile gonadotropin-releasing hormone therapy is associated with earlier spermatogenesis compared to combined gonadotropin therapy in patients with congenital hypogonadotropic hypogonadism. Asian J. Androl. 2017, 19, 680–685. [Google Scholar] [CrossRef]
- Leener De, A.; Montanelli, L.; Van Durme, J.; Chae, H.; Smits, G.; Vassart, G.; Costagliola, S. Presence and absence of follicle-stimulating hormone receptor mutations provide some insights into spontaneous ovarian hyperstimulation syndrome physiopathology. J. Clin. Endocrinol. Metab. 2006, 91, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.A.; Joseph, S.; Mahale, S.D. From cell surface to signalling and back: The life of the mammalian FSH receptor. FEBS J. 2020, 288, 2673–2696. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.C. Efficacy, efficiency and effectiveness of gonadotropin therapy for infertility treatment. Med. Express 2015, 2. [Google Scholar] [CrossRef]
- Pang, S.C. Use of Follicle-Stimulating Hormone for the Treatment of Female Infertility—Current Concepts. Women’s Health 2017, 1, 87–95. [Google Scholar] [CrossRef]
- Rashidi, M.; Najmi, Z.; Mobasseri, A. Advantages of Recombinant Follicle-Stimulating Hormone over Human Menopausal Gonadotropin in Intrauterine Insemination: A Randomized Clinical Trial in Polycystic Ovary Syndrome-Associated Infertility. Gynecol. Obstet. Investig. 2016, 81, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Coelingh Bennink, H.J.T.; Fauser, B.C.J.M.; Out, H.J. Recombinant follicle-stimulating hormone (FSH.; Puregon) is more efficient than urinary FSH (Metrodin) in women with clomiphene citrate-resistant, normogonadotropic, chronic anovulation: A prospective, multicenter, assessor-blind, randomized, clinical tria. Fertil. Steril. 1998, 69, 19–25. [Google Scholar] [CrossRef]
- Hayden, C.J.; Rutherford, A.J.; Balen, A.H. Induction of ovulation with the use of a starting dose of 50 units of recombinant human follicle-stimulating hormone (Puregon∗). Fertil. Steril. 1999, 71, 106–108. [Google Scholar] [CrossRef]
- Huirne, J.A.F.; Lambalk, C.B.; Van Loenen, A.C.D.; Schats, R.; Hompes, P.G.A.; Fauser, B.C.J.M.; Macklon, N.S. Contemporary pharmacological manipulation in assisted reproduction. Drugs 2004, 64, 297–322. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Kim, I.H.; Izadyar, F.; Hyttel, P.; Bevers, M.M.; Dieleman, S.J.; Hendriksen, P.J.; Vos, P.L. Impaired Final Follicular Maturation in Heifers after Superovulation with Recombinant Human FSH. Reproduction 2001, 121, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Jones, A.L.; Moore, K.; Looney, C.R.; Bondioli, K.R. Superovulation of cattle with a recombinant-DNA bovine follicle stimulating hormone. Anim. Reprod. Sci. 1993, 33, 71–82. [Google Scholar] [CrossRef]
- Carvalho, P.D.; Hackbart, K.S.; Bender, R.W.; Baez, G.M.; Dresch, A.R.; Guenther, J.N.; Souza, A.H.; Fricke, P.M. Use of a single injection of long-acting recombinant bovine FSH to superovulate Holstein heifers: A preliminary study. Theriogenology 2014, 82, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Bó, G.A.; Mapletoft, R.J. Superstimulation of ovarian follicles in cattle: Gonadotropin treatment protocols and FSH profiles. Theriogenology 2020, 150, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Bó, G.A.; Mapletoft, R.J. Historical perspectives and recent research on superovulation in cattle. Theriogenology 2014, 81, 38–48. [Google Scholar] [CrossRef]
- Josefsson, I. Presence of Hypergonadotropic Hypogonadism in Dogs with Primary Adrenocortical Insufficiency; Second cycle, A2E; SLU, Dept. of Clinical Sciences: Uppsala, Sweden, 2021. [Google Scholar]
- De Tassigny, X.D.A.; Fagg, L.A.; Dixon, J.P.C.; Day, K.; Leitch, H.G.; Hendrick, A.G.; Zahn, D.; Franceschini, I.; Caraty, A.; Carlton, M.B.L.; et al. Hypogonadotropic hypogonadism in mice lacking a functional Kiss1 gene. Proc. Natl. Acad. Sci. USA 2007, 104, 10714–10719. [Google Scholar] [CrossRef] [PubMed]
- Lapatto, R.; Pallais, J.C.; Zhang, D.; Chan, Y.M.; Mahan, A.; Cerrato, F.; Le, W.W.; Hoffman, G.E.; Seminara, S.B. Kiss1-/- mice exhibit more variable hypogonadism than Gpr54 -/- mice. Endocrinology 2007, 148, 4927–4936. [Google Scholar] [CrossRef]
- Lents, C.A. Review: Kisspeptin and reproduction in the pig. Animal 2019, 13, 2986–2999. [Google Scholar] [CrossRef]
- Orisaka, M.; Miyazaki, Y.; Shirafuji, A.; Tamamura, C.; Tsuyoshi, H.; Tsang, B.K.; Yoshida, Y. The role of pituitary gonadotropins and intraovarian regulators in follicle development: A mini-review. Reprod. Med. Biol. 2021, 20, 169–175. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef]
- Collodel, G.; Cantara, S.; Di Cairano, G.; Moretti, E.; Taglianetti, S.; Pacini, F. Alterations of the FSH and LH receptor genes and evaluation of sperm ultrastructure in men with idiopathic hypergonadotropic hypogonadism. J. Assist. Reprod. Genet. 2013, 30, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Mao, J.; Cui, M.; Liu, Z.; Wang, X.; Xiong, S.; Nie, M.; Wu, X. Novel FSHβ mutation in a male patient with isolated FSH deficiency and infertility. Eur. J. Med. Genet. 2017, 60, 335–339. [Google Scholar] [CrossRef]
- La Marca, A.; Sighinolfi, G.; Argento, C.; Grisendi, V.; Casarini, L.; Volpe, A.; Simoni, M. Polymorphisms in gonadotropin and gonadotropin receptor genes as markers of ovarian reserve and response in in vitro fertilization. Fertil. Steril. 2013, 99, 970–978.e1. [Google Scholar] [CrossRef]
- Laven, J.S.E. Follicle Stimulating Hormone Receptor (FSHR) Polymorphisms and Polycystic Ovary Syndrome (PCOS). Front. Endocrinol. 2019, 10, 23. [Google Scholar] [CrossRef]
- Kim, S.; Pyun, J.; Cha, D.; Ko, J.; Kwack, K. Epistasis between FSHR and CYP19A1 polymorphisms is associated with premature ovarian failure. Fertil. Steril. 2011, 95, 2585–2588. [Google Scholar] [CrossRef]
- Haller-Kikkatalo, K.; Salumets, A.; Uibo, R. Review on autoimmune reactions in female infertility: Antibodies to follicle stimulating hormone. Clin. Dev. Immunol. 2012, 2012, 1–15. [Google Scholar] [CrossRef]
- Geva, E.; Amit, A.; Lerner-Geva, L.; Lessing, J.B. Autoimmunity and reproduction. Fertil. Steril. 1997, 67, 599–611. [Google Scholar] [CrossRef]
- Haller, K.; Salumets, A.; Grigorova, M.; Talja, I.; Salur, L.; Béné, M.; Laan, M.; Uibo, R. Putative predictors of antibodies against follicle-stimulating hormone in female infertility: A study based on in vitro fertilization patients. Am. J. Reprod. Immunol. 2007, 57, 193–200. [Google Scholar] [CrossRef]
- Jankowska, K. Premature ovarian failure. Prz. Menopauzalny Menopause Rev. 2017, 16, 51–56. [Google Scholar] [CrossRef]
- Giglio, T.; Imro, M.A.; Filaci, G.; Scudeletti, M.; Puppo, F.; De Cecco, L.; Indiveri, F.; Costantini, S. Immune cell circulating subsets are affected by gonadal function. Life Sci. 1994, 54, 1305–1312. [Google Scholar] [CrossRef]
- Alberio, R.; Kobayashi, T.; Surani, M.A. Conserved features of non-primate bilaminar disc embryos and the germline. Stem Cell Rep. 2021, 16, 1078–1092. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Yamaji, M. Primordial germ cells in mice. Cold Spring Harb. Perspect. Biol. 2012, 4, a008375. [Google Scholar] [CrossRef]
- Wrobel, K.; Süss, F. Identification and temporospatial distribution of bovine primordial germ cells prior to gonadal sexual differentiation. Anat. Embryol. 1998, 197, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Maj, S.; Hyldig, W.; Ostrup, O.; Vejlsted, M.; Thomsen, P.D. Changes of DNA Methylation Level and Spatial Arrangement of Primordial Germ Cells in Embryonic Day 15 to Embryonic Day 28 Pig Embryos 1. Biol. Reprod. 2011, 84, 1087–1093. [Google Scholar] [CrossRef]
- Rey, R.; Josso, N.; Racine, C. Sexual Differentiation. 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279001/ (accessed on 1 June 2021).
- Fechner, P. The role of SRY in mammalian sex determination. Acta Paediatr. Jpn. Overseas Ed. 1996, 38, 380–389. [Google Scholar] [CrossRef]
- Koopman, P.; Münsterberg, A.; Capel, B.; Vivian, N.; Lovell-Badge, R. Expression of a candidate sex-determining gene during mouse testis differentiation. Nature 1990, 348, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Hacker, A.; Capel, B.; Goodfellow, P.; Lovell-Badge, R. Expression of Sry, the mouse sex determining gene. Development 1995, 121, 1603–1614. [Google Scholar] [CrossRef]
- Jeske, Y.; Bowles, J.; Greenfield, A.; Koopman, P. Expression of a linear Sry transcript in the mouse genital ridge. Nat. Genet. 1995, 10, 480–482. [Google Scholar] [CrossRef]
- Morais Da Silva, S.; Hacker, A.; Harleym, V.; Goodfellow, P.; Swain, A.; Lovell-Badge, R. Sox9 expression during gonadal development implies a conserved role for the gene in testis differentiation in mammals and birds. Nat. Genet. 1996, 14, 62–68. [Google Scholar] [CrossRef]
- Meyers-Wallen, V.; Bowman, L.; Acland, G.; Palmer, V.; Schlafer, D.; Fajt, V. Sry-negative XX sex reversal in the German shorthaired pointer dog. J. Hered. 1995, 86, 369–374. [Google Scholar] [CrossRef]
- McElreavey, K.; Vilain, E.; Cotinot, C.; Payen, E.; Fellous, M. Control of sex determination in animals. Eur. J. Biochem. 1993, 218, 769–783. [Google Scholar] [CrossRef]
- Berkovitz, G.; Fechner, P.; Marcantonio, S.; Bland, G.; Stetten, G.; Goodfellow, P.; Smith, K.; Migeon, C. The role of the sex-determining region of the Y chromosome (SRY) in the etiology of 46,XX true hermaphroditism. Hum. Genet. 1992, 88, 411–416. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, J.; Li, H.; Lv, J.; Wei, R.; Cong, Y.; Liu, Z. bFGF signaling-mediated reprogramming of porcine primordial germ cells. Cell Tissue Res. 2016, 364, 429–441. [Google Scholar] [CrossRef]
- De Souza, A.F.; Godoy Pieri, N.C.; Roballo, K.C.S.; Bressan, F.F.F.; Casals, J.B.; Ambrósio, C.E.; Perecin, F.; Martins, D.S. Dynamics of male canine germ cell development. PLoS ONE 2018, 13, 1–22. [Google Scholar] [CrossRef]
- Endo, T.; Mikedis, M.M.; Nicholls, P.K.; Page, D.C.; de Rooij, D.G. Retinoic acid and germ cell development in the ovary and testis. Biomolecules 2019, 9, 775. [Google Scholar] [CrossRef]
- McLaren, A. Meiosis and differentiation of mouse germ cells. Symp. Soc. Exp. Biol. 1984, 38, 7–23. [Google Scholar]
- Dolci, S.; Campolo, F.; De Felici, M. Gonadal development and germ cell tumors in mouse and humans. Semin. Cell Dev. Biol. 2015, 45, 114–123. [Google Scholar] [CrossRef]
- Tang, W.W.C.; Kobayashi, T.; Irie, N.; Dietmann, S.; Surani, M.A. Specification and epigenetic programming of the human germ line. Nat. Rev. Genet. 2016, 17, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Lavoir, M.-C.; Basrur, P.K.; Betteridge, K.J. Isolation and identification of germ cells from fetal bovine ovaries. Mol. Reprod. Dev. 1994, 37, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Dunkel, L.; Tilly, J.L.; Shikone, T.; Nishimori, K.; Hsueh, A.J.W. Follicle-stimulating hormone receptor expression in the rat ovary: Increases during prepubertal development and regulation by the opposing actions of transforming growth factors β and α. Biol. Reprod. 1994, 50, 940–948. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goxe, B.; Prunier, A.; Remy, J.J.; Salesse, R. Ontogeny of gonadal luteinizing hormone and follicle-stimulating hormone receptors in the fetal pig and related changes in gonadotropin and testosterone secretion. Biol. Reprod. 1993, 49, 609–616. [Google Scholar] [CrossRef][Green Version]
- Bielańska-Osuchowska, Z. Oogenesis in pig ovaries during the prenatal period: Ultrastructure and morphometry. Reprod. Biol. 2006, 6, 161–193. [Google Scholar]
- Nakamura, M.; Nakamura, K.; Igarashi, S.; Tano, M.; Miyamoto, K.; Ibuki, Y.; Minegishi, T. Interaction between activin A and cAMP in the induction of FSH receptor in cultured rat granulosa cells. J. Endocrinol. 1995, 147, 103–110. [Google Scholar] [CrossRef]
- Romero, C.; Paredes, A.; Dissen, G.A.; Ojeda, S.R. Nerve growth factor induces the expression of functional FSH receptors in newly formed follicles of the rat ovary. Endocrinology 2002, 143, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Wandji, S.A.; Pelletier, G.; Sirard, M.A. Ontogeny and cellular localization of 125I-labeled insulin-like growth factor-I, 125I-labeled follicle-stimulating hormone, and 125I-labeled human chorionic gonadotropin binding sites in ovaries from bovine fetuses and neonatal calves. Biol. Reprod. 1992, 47. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Albee, L. Requirement for follicle-stimulating hormone action in the formation of primordial follicles during perinatal ovarian development in the hamster. Endocrinology 2000, 141, 4449–4456. [Google Scholar] [CrossRef] [PubMed]
- White, S.S.; Ojeda, S.R. Changes in ovarian luteinizing hormone and follicle-stimulating hormone receptor content and in gonadotropin-induced ornithine decarboxylase activity during prepubertal and pubertal development of the female rat. Endocrinology 1981, 109, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Sokka, T.; Huhtaniemi, I. Ontogeny of gonadotrophin receptors and gonadotrophin-stimulated cyclic AMP production in the neonatal rat ovary. J. Endocrinol. 1990, 127, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, S.A.; Kohen, F.; Ausher, J.; Zor, U.; Lindner, H.R. Hormonal stimulation of oestradiol 17β release from the rat ovary during early postnatal development. J. Endocrinol. 1976, 68, 343–344. [Google Scholar] [CrossRef]
- McLaren, A. Primordial germ cells in the mouse. Dev. Biol. 2003, 262, 1–15. [Google Scholar] [CrossRef]
- Rouiller-Fabre, V.; Muczynski, V.; Lambrot, R.; Lécureuil, C.; Coffigny, H.; Pairault, C.; Moison, D.; Angenard, G.; Bakalska, M.; Courtot, A.; et al. Ontogenesis of testicular function in humans. Folia Histochem. Cytobiol. 2009, 47, 19–24. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Monteiro, A.; Abel, M. Testicular development in mice lacking receptors for follicle stimulating hormone and androgen. PLoS ONE 2012, 7, e35136. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J. Hormonal control of germ cell development and spermatogenesis. Semin. Cell Dev. Biol. 2014, 29, 55–65. [Google Scholar] [CrossRef]
- Meachem, S.J.; Ruwanpura, S.M.; Ziolkowski, J.; Ague, J.M.; Skinner, M.K.; Loveland, K.L. Developmentally distinct in vivo effects of FSH on proliferation and apoptosis during testis maturation. J. Endocrinol. 2005, 186, 429–446. [Google Scholar] [CrossRef]
- Boulogne, B.; Olaso, R.; Levacher, C.; Durand, P.; Habert, R. Apoptosis and mitosis in gonocytes of the rat testis during foetal and neonatal development. Int. J. Androl. 1999, 22, 356–365. [Google Scholar] [CrossRef]
- Meehan, T.; Schlatt, S.; O’Bryan, M.K.; De Kretser, D.M.; Loveland, K.L. Regulation of germ cell and sertoli cell development by activin, follistatin, and FSH. Dev. Biol. 2000, 220, 225–237. [Google Scholar] [CrossRef]
- Huhtaniemi, I.T.; Yamamoto, M.; Ranta, T.; Jalkanen, J.; Jaffe, R.B. Follicle-stimulating hormone receptors appear earlier in the primate fetal testis than in the ovary. J. Clin. Endocrinol. Metab. 1987, 65, 1210–1214. [Google Scholar] [CrossRef]
- Plant, T.M.; Marshall, G.R. The functional significance of FSH in spermatogenesis and the control of its secretion in male primates. Endocr. Rev. 2001, 22, 764–786. [Google Scholar] [CrossRef]
- Warren, D.W.; Huhtaniemi, I.T.; Tapanainen, J.; Dufau, M.L.; Catt, K.J. Ontogeny of gonadotropin receptors in the fetal and neonatal rat testis. Endocrinology 1984, 114, 470–476. [Google Scholar] [CrossRef]
- Meroni, S.B.; Galardo, M.N.; Rindone, G.; Gorga, A.; Riera, M.F.; Cigorraga, S.B. Molecular mechanisms and signaling pathways involved in Sertoli cell proliferation. Front. Endocrinol. 2019, 10, 224. [Google Scholar] [CrossRef]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and functional maturation of Sertoli cells, and their relevance to disorders of testis function in adulthood. Reproduction 2003, 125, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Musnier, A.; Heitzler, D.; Boulo, T.; Tesseraud, S.; Durand, G.; Lécureuil, C.; Guillou, H.; Poupon, A.; Reiter, E.; Crépieux, P. Developmental regulation of p70 S6 kinase by a G protein-coupled receptor dynamically modelized in primary cells. Cell. Mol. Life Sci. 2009, 66, 3487–3503. [Google Scholar] [CrossRef] [PubMed]
- Riera, M.; Regueira, M.; Galardo, M.; Pellizzari, E.; Meroni, S.; Cigorraga, S. Signal transduction pathways in FSH regulation of rat Sertoli cell proliferation. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E914–E923. [Google Scholar] [CrossRef]
- Kumar, T.R.; Wang, Y.; Lu, N.; Matzuk, M.M. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat. Genet. 1997, 15, 201–204. [Google Scholar] [CrossRef] [PubMed]
- McGee, E.A.; Hsueh, A.J.W. Initial and Cyclic Recruitment of Ovarian Follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Palermo, R. Differential actions of FSH and LH during folliculogenesis. Reprod. Biomed. Online 2007, 15, 326–337. [Google Scholar] [CrossRef]
- Fortune, J.E.; Cushman, R.A.; Wahl, C.M.; Kito, S. The primordial to primary follicle transition. Mol. Cell. Endocrinol. 2000, 163, 53–60. [Google Scholar] [CrossRef]
- Skinner, M.K. Regulation of primordial follicle assembly and development. Hum. Reprod. Update 2005, 11, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Kulick, L.J.; Bergfelt, D.R.; Kot, K.; Ginther, O.J. Follicle selection in cattle: Follicle deviation and codominance within sequential waves. Biol. Reprod. 2001, 65, 839–846. [Google Scholar] [CrossRef]
- Morbeck, D.E.; Flowers, W.L.; Britt, J.H. Response of porcine granulosa cells isolated from primary and secondary follicles to FSH, 8-bromo-cAMP and epidermal growth factor in vitro. J. Reprod. Fertil. 1993, 99, 577–584. [Google Scholar] [CrossRef]
- Wandji, S.A.; Eppig, J.J.; Fortune, I.E. FSH and growth factors affect the growth and endocrine function in vitro of granulosa cells of bovine preantral follicles. Theriogenology 1996, 45, 817–832. [Google Scholar] [CrossRef]
- Wandji, S.A.; Fortier, M.A.; Sirard, M.A. Differential response to gonadotropins and prostaglandin E2 in ovarian tissue during prenatal and postnatal development in cattle. Biol. Reprod. 1992, 46, 1034–1041. [Google Scholar] [CrossRef][Green Version]
- Eppig, J.J.; O’Brien, M.J. Development in vitro of mouse oocytes from primordial follicles. Biol. Reprod. 1996, 54, 197–207. [Google Scholar] [CrossRef]
- Gougeon, A. Regulation of ovarian follicular development in primates: Facts and hypotheses. Endocr. Rev. 1996, 17, 121–155. [Google Scholar] [CrossRef]
- Durlej, M.; Knapczyk-Stwora, K.; Duda, M.; Galas, J.; Slomczynska, M. The expression of FSH receptor (FSHR) in the neonatal porcine ovary and its regulation by flutamide. Reprod. Domest. Anim. 2011, 46, 377–384. [Google Scholar] [CrossRef]
- Van den Hurk, R.; Bevers, M.M.; Beckers, J.F. In-vivo and in-vitro development of preantral follicles. Theriogenology 1997, 47, 73–82. [Google Scholar] [CrossRef]
- Yamoto, M.; Shima, K.; Nakano, R. Gonadotropin receptors in human ovarian follicles and corpora lutea throughout the menstrual cycle. Horm. Res. 1992, 37 (Suppl. 1), 5–11. [Google Scholar] [CrossRef] [PubMed]
- Candelaria, J.I.; Rabaglino, M.B.; Denicol, A.C. Ovarian preantral follicles are responsive to FSH as early as the primary stage of development. J. Endocrinol. 2020, 247, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Tisdall, D.J.; Watanabe, K.; Hudson, N.L.; Smith, P.; McNatty, K.P. FSH receptor gene expression during ovarian follicle development in sheep. J. Mol. Endocrinol. 1995, 15, 273–281. [Google Scholar] [CrossRef]
- Rajapaksha, W.; Robertson, L.; O’Shaughnessy, P. Expression of follicle-stimulating hormone-receptor mRNA alternate transcripts in bovine granulosa cells during luteinization in vivo and in vitro. Mol. Cell. Endocrinol. 1996, 120, 25–30. [Google Scholar] [CrossRef]
- Oktay, K.; Briggs, D.; Gosden, R.G. Ontogeny of Follicle-Stimulating Hormone Receptor Gene Expression in Isolated Human Ovarian Follicles 1. J. Clin. Endocrinol. Metab. 1997, 82, 3748–3751. [Google Scholar] [CrossRef]
- Lan, R.X.; Liu, F.; He, Z.B.; Chen, C.; Liu, S.J.; Shi, Y.; Liu, Y.L.; Yoshimura, Y.; Zhang, M. Immunolocalization of GnRHRI, gonadotropin receptors, PGR, and PGRMCI during follicular development in the rabbit ovary. Theriogenology 2014, 81, 1139–1147. [Google Scholar] [CrossRef]
- De Castro, F.C.; Cruz, M.H.C.; Leal, C. Role of Growth Differentiation Factor 9 and Bone Morphogenetic Protein 15 in Ovarian Function and Their Importance in Mammalian Female Fertility—A Review. Asian-Australas. J. Anim. Sci. 2015, 29, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Spicer, L.; Aad, P.; Allen, D.; Mazerbourg, S.; Hsueh, A.J. Growth differentiation factor-9 has divergent effects on proliferation and steroidogenesis of bovine granulosa cells. J. Endocrinol. 2006, 189, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, L.; Sun, Y.; Wu, D.; Li, J.; Zhu, L.; Jiang, S.; Pan, X. The optimized research of the in vitro culture of preantral follicles in mice. J. Clin. Lab. Anal. 2020, 34, e23498. [Google Scholar] [CrossRef]
- El-Hayek, S.; Demeestere, I.; Clarke, H.J. Follicle-stimulating hormone regulates expression and activity of epidermal growth factor receptor in the murine ovarian follicle. Proc. Natl. Acad. Sci. USA 2014, 111, 16778–16783. [Google Scholar] [CrossRef]
- Balla, A.; Danilovich, N.; Yang, Y.; Sairam, M.R. Dynamics of ovarian development in the FORKO immature mouse: Structural and functional implications for ovarian reserve. Biol. Reprod. 2003, 69, 1281–1293. [Google Scholar] [CrossRef]
- Pakarainen, T.; Zhang, F.P.; Nurmi, L.; Poutanen, M.; Huhtaniemi, I. Knockout of luteinizing hormone receptor abolishes the effects of follicle-stimulating hormone on preovulatory maturation and ovulation of mouse graafian follicles. Mol. Endocrinol. 2005, 19, 2591–2602. [Google Scholar] [CrossRef]
- Nagashima, J.B.; Wildt, D.E.; Travis, A.J.; Songsasen, N. Activin promotes growth and antral cavity expansion in the dog ovarian follicle. Theriogenology 2019, 129, 168–177. [Google Scholar] [CrossRef]
- Dewailly, D.; Robin, G.; Peigne, M.; Decanter, C.; Pigny, P.; Catteau-Jonard, S. Interactions between androgens, FSH, anti-Mullerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum. Reprod. Update 2016, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Aittomäki, K.; Dieguez Lucena, J.; Pakarinen, P.; Sistonen, P.; Tapanainen, J.; Gromoll, J.; Kaskikari, R.; Sankila, E.M.; Lehväslaiho, H.; Reyes Engel, A.; et al. Mutation in the follicle-stimulating hormone receptor gene causes hereditary hypergonadotropic ovarian failure. Cell 1995, 82, 959–968. [Google Scholar] [CrossRef]
- Touraine, P.; Beau, I.; Gougeon, A.; Meduri, G.; Desroches, A.; Pichard, C.; Detoeuf, M.; Paniel, B.; Prieur, M.; Zorn, J.R.; et al. New natural inactivating mutations of the follicle-stimulating hormone receptor: Correlations between receptor function and phenotype. Mol. Endocrinol. 1999, 13, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Abel, M.H.; Wootton, A.N.; Wilkins, V.; Huhtaniemi, I.; Knight, P.G.; Charlton, H.M. The effect of a null mutation in the follicle-stimulating hormone receptor gene on mouse reproduction. Endocrinology 2000, 141, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.K.; Telfer, E.E.; Webb, R.; Baird, D.T. Evidence of a role for follicle-stimulating hormone in controlling the rate of preantral follicle development in sheep. Endocrinology 2004, 145, 1870–1879. [Google Scholar] [CrossRef][Green Version]
- Patsoula, E.; Loutradis, D.; Drakakis, P.; Michalas, L.; Bletsa, R.; Michalas, S. Messenger RNA expression for the follicle-stimulating hormone receptor and luteinizing hormone receptor in human oocytes and preimplantation-stage embryos. Fertil. Steril. 2003, 79, 1187–1193. [Google Scholar] [CrossRef]
- Méduri, G.; Charnaux, N.; Driancourt, M.; Combettes, L.; Granet, P.; Vannier, B.; Loosfelt, H.; Milgrom, E. Follicle-stimulating hormone receptors in oocytes? J. Clin. Endocrinol. Metab. 2002, 87, 2266–2276. [Google Scholar] [CrossRef] [PubMed]
- Ruwanpura, S.M.; McLachlan, R.I.; Meachem, S.J. Hormonal regulation of male germ cell development. J. Endocrinol. 2010, 205, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, C.; Stévant, I.; Borel, C.; Conne, B.; Pitetti, J.L.; Calvel, P.; Kaessmann, H.; Jégou, B.; Chalmel, F.; Nef, S. Research resource: The dynamic transcriptional profile of sertoli cells during the progression of spermatogenesis. Mol. Endocrinol. 2015, 29. [Google Scholar] [CrossRef] [PubMed]
- Regueira, M.; Riera, M.F.; Galardo, M.N.; del Camberos, M.; Pellizzari, E.H.; Cigorraga, S.B.; Meroni, S.B. FSH and bFGF regulate the expression of genes involved in Sertoli cell energetic metabolism. Gen. Comp. Endocrinol. 2015, 222, 124–133. [Google Scholar] [CrossRef]
- Gromoll, J.; Simoni, M.; Nieschlag, E. An activating mutation of the follicle-stimulating hormone receptor autonomously sustains spermatogenesis in a hypophysectomized man. J. Clin. Endocrinol. Metab. 1996, 81, 1367–1370. [Google Scholar] [CrossRef]
- McLachlan, R.I.; Wreford, N.G.; Meachem, S.J.; De Kretser, D.M.; Robertson, D.M. Effects of Testosterone on Spermatogenic Cell Populations in the Adult Rat1. Biol. Reprod. 1994, 51, 945–955. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McLachlan, R.I.; Wreford, N.G.; Tsonis, C.; De Kretser, D.M.; Robertson, D.M. Testosterone Effects on Spermatogenesis in the Gonadotropin-Releasing Hormone-Immunized Rat1. Biol. Reprod. 1994, 50, 271–280. [Google Scholar] [CrossRef][Green Version]
- McLean, D.; Friel, P.; Pouchnik, D.; Griswold, M. Oligonucleotide microarray analysis of gene expression in follicle-stimulating hormone-treated rat Sertoli cells. Mol. Endocrinol. 2002, 16, 2780–2792. [Google Scholar] [CrossRef] [PubMed]
- Sadate-Ngatchou, P.; Pouchnik, D.; Griswold, M. Follicle-stimulating hormone induced changes in gene expression of murine testis. Mol. Endocrinol. 2004, 18, 2805–2816. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Walker, W.H.; Cheng, J. FSH and testosterone signaling in Sertoli cells. Reproduction 2005, 130, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Godmann, M.; Katz, J.P.; Guillou, F.; Simoni, M.; Kaestner, K.H.; Behr, R. Krüppel-like factor 4 is involved in functional differentiation of testicular Sertoli cells. Dev. Biol. 2008, 315, 552–566. [Google Scholar] [CrossRef]
- Sluka, P.; O’Donnell, L.; Bartles, J.R.; Stanton, P.G. FSH regulates the formation of adherens junctions and ectoplasmic specialisations between rat Sertoli cells in vitro and in vivo. J. Endocrinol. 2006, 189, 381–395. [Google Scholar] [CrossRef]
- Soffientini, U.; Rebourcet, D.; Abel, M.H.; Lee, S.; Hamilton, G.; Fowler, P.A.; Smith, L.B.; O’Shaughnessy, P.J. Identification of Sertoli cell-specific transcripts in the mouse testis and the role of FSH and androgen in the control of Sertoli cell activity. BMC Genomics 2017, 18, 972. [Google Scholar] [CrossRef]
- Simorangkir, D.R.; Ramaswamy, S.; Marshall, G.R.; Pohl, C.R.; Plant, T.M. A selective monotropic elevation of FSH, but not that of LH, amplifies the proliferation and differentiation of spermatogonia in the adult rhesus monkey (Macaca mulatta). Hum. Reprod. 2009, 24, 1584–1595. [Google Scholar] [CrossRef]
- Pieri, N.C.G.; Mançanares, A.C.F.; De Souza, A.F.; Fernandes, H.; Diaza, A.M.G.; Bressan, F.F.; Roballo, K.C.S.; Casals, J.B.; Binelli, M.; Ambrósio, C.E.; et al. Xenotransplantation of canine spermatogonial stem cells (cSSCs) regulated by FSH promotes spermatogenesis in infertile mice. Stem Cell Res. Ther. 2019, 10, 135. [Google Scholar] [CrossRef]
- Baumgarten, S.C.; Stocco, C. Granulosa cells. In Encyclopedia of Reproduction; Elsevier: Amsterdam, The Netherlands, 2018; pp. 8–13. ISBN 9780128151457.1. [Google Scholar]
- Palma, G.A.; Argañaraz, M.E.; Barrera, A.D.; Rodler, D.; Mutto, A.; Sinowatz, F. Biology and Biotechnology of Follicle Development. Sci. World J. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Hillier, S.G. Gonadotropic control of ovarian follicular growth and development. Mol. Cell. Endocrinol. 2001, 179, 39–46. [Google Scholar] [CrossRef]
- Silva, J.M.; Price, C.A. Effect of follicle-stimulating hormone on steroid secretion and messenger ribonucleic acids encoding cytochromes P450 aromatase and cholesterol side-chain cleavage in bovine granulosa cells in vitro. Biol. Reprod. 2000, 62, 186–191. [Google Scholar] [CrossRef][Green Version]
- Piquette, G.N.; Lapolt, P.S.; Oikawa, M.; Hsueh, A.J.W.; Hsueh, A.J.W. Regulation of luteinizing hormone receptor messenger ribonucleic acid levels by gonadotropins, growth factors, and gonadotropin-releasing hormone in cultured rat granulosa cells. Endocrinology 1991, 128, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Hunzicker-Dunn, M.; Maizels, E.T. FSH signaling pathways in immature granulosa cells that regulate target gene expression: Branching out from protein kinase A. Cell. Signal. 2006, 18, 1351. [Google Scholar] [CrossRef]
- Sasson, R.; Dantes, A.; Tajima, K.; Amsterdam, A. Novel genes modulated by FSH in normal and immortalized FSH-responsive cells: New insights into the mechanism of FSH action. FASEB J. 2003, 17, 1256–1266. [Google Scholar] [CrossRef]
- Bonnet, A.; Frappart, P.O.; Dehais, P.; Tosser-Klopp, G.; Hatey, F. Identification of differential gene expression in in vitro FSH treated pig granulosa cells using suppression subtractive hybridization. Reprod. Biol. Endocrinol. 2006, 4, 35. [Google Scholar] [CrossRef]
- Verbraak, E.; Veld, E.V.; Koerkamp, M.G.; Roelen, B.; van Haeften, T.; Stoorvogel, W.; Zijlstra, C. Identification of genes targeted by FSH and oocytes in porcine granulosa cells. Theriogenology 2011, 75, 362–376. [Google Scholar] [CrossRef]
- Heng, D.; Wang, Q.; Ma, X.; Tian, Y.; Xu, K.; Weng, X.; Hu, X.; Liu, W.; Zhang, C. Role of OCT4 in the Regulation of FSH-Induced Granulosa Cells Growth in Female Mice. Front. Endocrinol. 2020, 10, 915. [Google Scholar] [CrossRef]
- Adashi, E.Y. Endocrinology of the ovary. Hum. Reprod. 1994, 9, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Hunzicker-Dunn, M.; Mayo, K. Gonadotropin Signaling in the Ovary. In Knobil and Neill’s Physiology of Reproduction: Two-Volume Set; 2015; Volume 1, pp. 1–51. [Google Scholar] [CrossRef]
- Goodman, H.M. Hormonal Control of Reproduction in the Female. In Basic Medical Endocrinology; Elsevier Inc.: Amsterdam, The Netherlands, 2003; pp. 423–456. [Google Scholar] [CrossRef]
- Durlinger, A.L.L.; Gruijters, M.J.G.; Kramer, P.; Karels, B.; Kumar, T.R.; Matzuk, M.M.; Rose, U.M.; De Jong, F.H.; Uilenbroek, J.T.J.; Grootegoed, J.A.; et al. Anti-Müllerian hormone attenuates the effects of FSH on follicle development in the mouse ovary. Endocrinology 2001, 142, 4891–4899. [Google Scholar] [CrossRef] [PubMed]
- Pellatt, L.; Rice, S.; Dilaver, N.; Heshri, A.; Galea, R.; Brincat, M.; Brown, K.; Simpson, E.R.; Mason, H.D. Anti-Müllerian hormone reduces follicle sensitivity to follicle-stimulating hormone in human granulosa cells. Fertil. Steril. 2011, 96, 1246–1251.e1. [Google Scholar] [CrossRef]
- Skinner, M.K. Encyclopedia of Reproduction, 2nd ed.; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar] [CrossRef]
- Jozkowiak, M.; Hutchings, G.; Jankowski, M.; Kulcenty, K.; Mozdziak, P.; Kempisty, B.; Spaczynski, R.Z.; Piotrowska-Kempisty, H. The Stemness of Human Ovarian Granulosa Cells and the Role of Resveratrol in the Differentiation of MSCs-A Review Based on Cellular and Molecular Knowledge. Cells 2020, 9, 1418. [Google Scholar] [CrossRef]
- Prasasya, R.D.; Mayo, K.E. Notch signaling regulates differentiation and steroidogenesis in female mouse ovarian granulosa cells. Endocrinology 2018, 159, 184–198. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Cheung, M.Y.; Sia, Z.K.; Agapiou, D.; Corley, S.M.; Wilkins, M.R.; Richani, D.; Harrison, C.A.; Gilchrist, R.B. Non-canonical cyclic AMP SMAD1/5/8 signalling in human granulosa cells. Mol. Cell. Endocrinol. 2019, 490, 37–46. [Google Scholar] [CrossRef]
- Fortune, J.E.; Rivera, G.M.; Evans, A.C.O.; Turzillo, A.M. Differentiation of Dominant Versus Subordinate Follicles in Cattle. Biol. Reprod. 2001, 65, 648–654. [Google Scholar] [CrossRef]
- Fortune, J.E. Ovarian Follicular Growth and Development in Mammals1. Biol. Reprod. 1994, 50, 225–232. [Google Scholar] [CrossRef]
- Garcia-Velasco, J.A.; Rodríguez, S.; Agudo, D.; Pacheco, A.; Schneider, J.; Pellicer, A. FSH receptor in vitro modulation by testosterone and hCG in human luteinized granulosa cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 165, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, H.; Pope, W.F. Androgen receptor and follicle-stimulating hormone receptor in the pig ovary during the follicular phase of the estrous cycle. Mol. Reprod. Dev. 2002, 62, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, R.; Kalab, P.; Nagyova, E. Epidermal growth factor-receptor tyrosine kinase activity regulates expansion of porcine oocyte-cumulus cell complexes in vitro. Biol. Reprod. 2003, 68, 797–803. [Google Scholar] [CrossRef]
- Sugimura, S.; Richani, D.; Gilchrist, R.B. Follicular guidance for oocyte developmental competence. Anim. Reprod. 2018, 15, 721–726. [Google Scholar] [CrossRef]
- Bhartiya, D.; Singh, J. FSH-FSHR3-stem cells in ovary surface epithelium: Basis for adult ovarian biology, failure, aging, and cancer. Reproduction 2015, 149, R35–R48. [Google Scholar] [CrossRef]
- Parte, S.; Bhartiya, D.; Manjramkar, D.D.; Chauhan, A.; Joshi, A. Stimulation of ovarian stem cells by follicle stimulating hormone and basic fibroblast growth factor during cortical tissue culture. J. Ovarian Res. 2013, 6, 20. [Google Scholar] [CrossRef]
- Bhartiya, D.; Unni, S.; Parte, S.; Anand, S. Very small embryonic-like stem cells: Implications in reproductive biology. Biomed. Res. Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Bhartiya, D.; Sriraman, K.; Parte, S.; Patel, H. Ovarian stem cells: Absence of evidence is not evidence of absence. J. Ovarian Res. 2013, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Parte, S.; Bhartiya, D.; Telang, J.; Daithankar, V.; Salvi, V.; Zaveri, K.; Hinduja, I. Detection, characterization, and spontaneous differentiation in vitro of very small embryonic-like putative stem cells in adult mammalian ovary. Stem Cells Dev. 2011, 20, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Parte, S.; Bhartiya, D.; Patel, H.; Daithankar, V.; Chauhan, A.; Zaveri, K.; Hinduja, I. Dynamics associated with spontaneous differentiation of ovarian stem cells in vitro. J. Ovarian Res. 2014, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Skutella, T.; Hren, M.; Gruden, K.; Cvjeticanin, B.; Vogler, A.; Sinkovec, J. Isolation of small ssea-4-positive putative stem cells from the ovarian surface epithelium of adult human ovaries by two different methods. Biomed. Res. Int. 2013, 2013, 1–15. [Google Scholar] [CrossRef]
- Patel, H.; Bhartiya, D.; Parte, S.; Gunjal, P.; Yedurkar, S.; Bhatt, M. Follicle stimulating hormone modulates ovarian stem cells through alternately spliced receptor variant FSH-R3. J. Ovarian Res. 2013, 6, 52. [Google Scholar] [CrossRef]
- Sairam, M.R.; Babu, P.S. The tale of follitropin receptor diversity: A recipe for fine tuning gonadal responses? Mol. Cell. Endocrinol. 2007, 260, 163–171. [Google Scholar] [CrossRef]
- Mertens-Walker, I.; Baxter, R.C.; Marsh, D.J. Gonadotropin signalling in epithelial ovarian cancer. Cancer Lett. 2012, 324, 152–159. [Google Scholar] [CrossRef]
- Wreford, N.G.; Kumar, T.R.; Matzuk, M.M.; De Kretser, D.M. Analysis of the testicular phenotype of the follicle-stimulating hormone β-subunit knockout and the activin type II receptor knockout mice by stereological analysis. Endocrinology 2001, 142, 2916–2920. [Google Scholar] [CrossRef] [PubMed]
- Abel, M.H.; Baker, P.J.; Charlton, H.M.; Monteiro, A.; Verhoeven, G.; De Gendt, K.; Guillou, F.; O’Shaughnessy, P.J. Spermatogenesis and sertoli cell activity in mice lacking sertoli cell receptors for follicle-stimulating hormone and androgen. Endocrinology 2008, 149, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, R.; O’Donnell, L.; Meachem, S.; Stanton, P.; de David, M.; Pratis, K.; Robertson, D.M. Hormonal regulation of spermatogenesis in primates and man: Insights for development of the male hormonal contraceptive. J. Androl. 2002, 23, 149–162. [Google Scholar] [PubMed]
- Meachem, S.; Mclachlan, R.; Stanton, P.; Robertson, D.; Wreford, N.G. FSH immunoneutralization acutely impairs spermatogonial development in normal adult rats-PubMed. J. Androl. 1999, 20, 756–762. [Google Scholar] [PubMed]
- Ruwanpura, S.M.; McLachlan, R.I.; Stanton, P.G.; Meachem, S.J. Follicle-stimulating hormone affects spermatogonial survival by regulating the intrinsic apoptotic pathway in adult rats. Biol. Reprod. 2008, 78, 705–713. [Google Scholar] [CrossRef]
- Clermont, Y.; Antar, M. Duration of the cycle of the seminiferous epithelium and the spermatogonial renewal in the monkey Macaca arctoides. Am. J. Anat. 1973, 136, 153–165. [Google Scholar] [CrossRef]
- Clermont, Y.; Leblond, C.P. Differentiation and renewal of spermatogonia in the monkey, Macacus rhesus. Am. J. Anat. 1959, 104, 237–273. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Hansel, M.C.; Orwig, K.E. Spermatogonial stem cells in higher primates: Are there differences to those in rodents? Reproduction 2010, 139, 479. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial stem cells and spermatogenesis in mice, monkeys and men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef]
- Dym, M.; Kokkinaki, M.; He, Z. Spermatogonial stem cells: Mouse and human comparisons. Birth Defects Res. C Embryo Today 2009, 87, 27–34. [Google Scholar] [CrossRef]
- Meachem, S.; von Schönfeldt, V.; Schlatt, S. Spermatogonia: Stem cells with a great perspective. Reproduction 2001, 121, 825–834. [Google Scholar] [CrossRef]
- Simorangkir, D.R.; Marshall, G.R.; Ehmcke, J.; Schlatt, S.; Plant, T.M. Prepubertal expansion of dark and pale type A spermatogonia in the rhesus monkey (Macaca mulatta) results from proliferation during infantile and juvenile development in a relatively gonadotropin independent manner. Biol. Reprod. 2005, 73, 1109–1115. [Google Scholar] [CrossRef][Green Version]
- McLachlan, R.I.; O’Donnell, L.; Stanton, P.G.; Balourdos, G.; Frydenberg, M.; De Kretser, D.M.; Robertson, D.M. Effects of testosterone plus medroxyprogesterone acetate on semen quality, reproductive hormones, and germ cell populations in normal young men. J. Clin. Endocrinol. Metab. 2002, 87, 546–556. [Google Scholar] [CrossRef]
- Eto, K.; Shiotsuki, M.; Sakai, T.; Abe, S. Nociceptin is upregulated by FSH signaling in Sertoli cells in murine testes. Biochem. Biophys. Res. Commun. 2012, 421, 678–683. [Google Scholar] [CrossRef]
- Eto, K.; Shiotsuki, M.; Abe, S.-I. Nociceptin induces Rec8 phosphorylation and meiosis in postnatal murine testes. Endocrinology 2013, 154, 2891–2899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Eto, K.; Honmyou, A.; Nakao, K.; Kiyonari, H.; Abé, S. Neuregulins are essential for spermatogonial proliferation and meiotic initiation in neonatal mouse testis. Development 2011, 138, 3159–3168. [Google Scholar] [CrossRef] [PubMed]
- Matthiesson, K.L.; McLachlan, R.I.; O’Donnell, L.; Frydenberg, M.; Robertson, D.M.; Stanton, P.G.; Meachem, S.J. The relative roles of follicle-stimulating hormone and luteinizing hormone in maintaining spermatogonial maturation and spermiation in normal men. J. Clin. Endocrinol. Metab. 2006, 91, 3962–3969. [Google Scholar] [CrossRef] [PubMed]
- Khanehzad, M.; Abbaszadeh, R.; Holakuyee, M.; Modarressi, M.; Nourashrafeddin, S. FSH regulates RA signaling to commit spermatogonia into differentiation pathway and meiosis. Reprod. Biol. Endocrinol. 2021, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Lindahl, M.; Hyvönen, M.E.; Parvinen, M.; De Rooij, D.G.; Hess, M.W.; Raatikainen-Ahokas, A.; Sainio, K.; Rauvala, H.; Lakso, M.; et al. Regulation of cell fate decision of undifferentiated spermatogonia by GDNF. Science 2000, 287, 1489–1493. [Google Scholar] [CrossRef]
- de Rooij, D. Regulation of spermatogonial stem cell behavior in vivo and in vitro 1. Anim. Reprod. 2006, 31, 130–134. [Google Scholar]
- Naughton, C.; Jain, S.; Strickland, A.; Gupta, A.; Milbrandt, J. Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol. Reprod. 2006, 74, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Wen, D.; Yu, Y.; Holst, P.; Luo, Y.; Fang, M.; Tamir, R.; Antonio, L.; Hu, Z.; Cupples, R.; et al. GDNF-induced activation of the ret protein tyrosine kinase is mediated by GDNFR-alpha, a novel receptor for GDNF. Cell 1996, 85, 1113–1124. [Google Scholar] [CrossRef]
- Tadokoro, Y.; Yomogida, K.; Ohta, H.; Tohda, A.; Nishimune, Y. Homeostatic regulation of germinal stem cell proliferation by the GDNF/FSH pathway. Mech. Dev. 2002, 113, 29–39. [Google Scholar] [CrossRef]
- Jijiwa, M.; Kawai, K.; Fukihara, J.; Nakamura, A.; Hasegawa, M.; Suzuki, C.; Sato, T.; Enomoto, A.; Asai, N.; Murakumo, Y.; et al. GDNF-mediated signaling via RET tyrosine 1062 is essential for maintenance of spermatogonial stem cells. Genes Cells 2008, 13, 365–374. [Google Scholar] [CrossRef]
- Kostereva, N.; Hofmann, M.C. Regulation of the Spermatogonial Stem Cell Niche. Reprod. Domest. Anim. 2008, 43, 386–392. [Google Scholar] [CrossRef]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. ROS are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef]
- Parekh, P.A.; Garcia, T.X.; Hofmann, M.C. Regulation of GDNF expression in Sertoli cells. Reproduction 2019, 157, R95–R107. [Google Scholar] [CrossRef]
- Chen, S.-R.R.; Liu, Y.-X.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by Sertoli cell signaling. Reproduction 2015, 149, R159–R167. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Avarbock, M.R.; Brinster, R.L. Glial cell line-derived neurotrophic factor regulation of genes essential for self-renewal of mouse spermatogonial stem cells is dependent on Src family kinase signaling. J. Biol. Chem. 2007, 282, 25842–25851. [Google Scholar] [CrossRef]
- Lee, J.; Kanatsu-Shinohara, M.; Inoue, K.; Ogunuki, N.; Miki, H.; Toyokuni, S.; Kimura, T.; Nakano, T.; Ogura, A.; Shinohara, T. Akt mediates self-renewal division of mouse spermatogonial stem cells. Development 2007, 134, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kokkinaki, M.; Dym, M. Signaling molecules and pathways regulating the fate of spermatogonial stem cells. Microsc. Res. Tech. 2009, 72, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.; Avarbock, M.; Telaranta, A.; Fearon, D.; Brinster, R. Identifying genes important for spermatogonial stem cell self-renewal and survival. Proc. Natl. Acad. Sci. USA 2006, 103, 9524–9529. [Google Scholar] [CrossRef]
- Wu, X.; Schmidt, J.; Avarbock, M.; Tobias, J.; Carlson, C.; Kolon, T.; Ginsberg, J.; Brinster, R. Prepubertal human spermatogonia and mouse gonocytes share conserved gene expression of germline stem cell regulatory molecules. Proc. Natl. Acad. Sci. USA 2009, 106, 21672–21677. [Google Scholar] [CrossRef]
- Chuykin, I.; Stauske, M.A.; Guan, K. Spermatogonial stem cells. In Regenerative Medicine; Springer: Dordrecht, The Netherlands, 2013; Volume 99, pp. 219–249. ISBN 9789400756908. [Google Scholar]
- He, Z.; Jiang, J.; Kokkinaki, M.; Golestaneh, N.; Hofmann, M.; Dym, M. Gdnf upregulates c-Fos transcription via the Ras/Erk1/2 pathway to promote mouse spermatogonial stem cell proliferation. Stem Cells 2008, 26, 266–278. [Google Scholar] [CrossRef]
- Lee, J.; Kanatsu-Shinohara, M.; Morimoto, H.; Kazuki, Y.; Takashima, S.; Oshimura, M.; Toyokuni, S.; Shinohara, T. Genetic reconstruction of mouse spermatogonial stem cell self-renewal in vitro by Ras-cyclin D2 activation. Cell Stem Cell 2009, 5, 76–86. [Google Scholar] [CrossRef]
- Yang, Y.; Han, C. GDNF stimulates the proliferation of cultured mouse immature Sertoli cells via its receptor subunit NCAM and ERK1/2 signaling pathway. BMC Cell Biol. 2010, 11, 78. [Google Scholar] [CrossRef]
- Sharma, M.; Braun, R.E. Cyclical expression of GDNF is required for spermatogonial stem cell homeostasis. Development 2018, 145, dev151555. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Jabarpour, M.; Tajik, P. Evaluation of the effect of follicular stimulating hormone on the in vitro bovine spermatogonial stem cells self-renewal: An experimental study. Int. J. Reprod. Biomed. 2017, 15, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M. GFRα-1 is a reliable marker of bovine gonocytes/undifferentiated spermatogonia: A mini-review. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2021, 50, 13–14. [Google Scholar] [CrossRef]
- Azizi, H.; Niazi Tabar, A.; Skutella, T.; Govahi, M. In vitro and in vivo determinations of the anti-GDNF family receptor alpha 1 antibody in mice by immunochemistry and RT-PCR. Int. J. Fertil. Steril. 2020, 14, 228–233. [Google Scholar] [CrossRef]
- Filatov, M.; Khramova, Y.; Parshina, E.; Bagaeva, T.; Semenova, M. Influence of gonadotropins on ovarian follicle growth and development in vivo and in vitro. Zygote 2017, 25, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.A.; Cadenas, J.; Sá, N.A.R.; Correia, H.H.V.; Guerreiro, D.D.; Lobo, C.H.; Alves, B.G.; Maside, C.; Gastal, E.L.; Rodrigues, A.P.R.; et al. In vitro culture of isolated preantral and antral follicles of goats using human recombinant FSH: Concentration-dependent and stage-specific effect. Anim. Reprod. Sci. 2018, 196, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhu, L.; Jin, L. Human Follicle in vitro Culture Including Activation, Growth, and Maturation: A Review of Research Progress. Front. Endocrinol. 2020, 11, 548. [Google Scholar] [CrossRef]
- Xu, J.; Bernuci, M.P.; Lawson, M.S.; Yeoman, R.R.; Fisher, T.E.; Zelinski, M.B.; Stouffer, R.L. Survival, growth, and maturation of secondary follicles from prepubertal, young, and older adult rhesus monkeys during encapsulated three-dimensional culture: Effects of gonadotropins and insulin. Reproduction 2010, 140, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ting, A.Y.; Tkachenko, O.; Xu, J.; Stouffer, R.L. Direct actions of androgen, estrogen and anti-Müllerian hormone on primate secondary follicle development in the absence of FSH in vitro. Hum. Reprod. 2017, 32, 2456–2464. [Google Scholar] [CrossRef]
- Caixeta, E.S.; Machado, M.F.; Ripamonte, P.; Price, C.; Buratini, J. Effects of FSH on the expression of receptors for oocyte-secreted factors and members of the EGF-like family during in vitro maturation in cattle. Reprod. Fertil. Dev. 2013, 25, 890–899. [Google Scholar] [CrossRef]
- Widayati, D.T.; Pangestu, M. Effect of follicle-stimulating hormone on Bligon goat oocyte maturation and embryonic development post in vitro fertilization. Vet. World 2020, 13, 2443–2446. [Google Scholar] [CrossRef]
- Silva, G.M.; Brito, I.R.; Sales, A.D.; Aguiar, F.L.N.; Duarte, A.B.G.; Araújo, V.R.; Vieira, L.A.; Magalhães-Padilha, D.M.; Lima, L.F.; Alves, B.G.; et al. In vitro growth and maturation of isolated caprine preantral follicles: Influence of insulin and FSH concentration, culture dish, coculture, and oocyte size on meiotic resumption. Theriogenology 2017, 90, 32–41. [Google Scholar] [CrossRef]
- Lee, H.-S.; Seo, Y.-I.; Yin, X.-J.; Cho, S.-G.; Lee, S.-S.; Kim, N.-H.; Cho, S.-K.; Kong, I.-K. Effect of Follicle Stimulation Hormone and Luteinizing Hormone on Cumulus Cell Expansion and In Vitro Nuclear Maturation of Canine Oocytes. Reprod. Domest. Anim. 2007, 42, 561–565. [Google Scholar] [CrossRef]
- Hu, M.; Du, Z.; Zhou, Z.; Long, H.; Ni, Q. Effects of serum and follicular fluid on the in vitro maturation of canine oocytes. Theriogenology 2020, 143, 10–17. [Google Scholar] [CrossRef]
- Wood, T.C.; Byers, A.P.; Jennette, B.E.; Wildt, D.E. Influence of protein and hormone supplementation on in vitro maturation and fertilization of domestic cat eggs. Reproduction 1995, 104, 315–323. [Google Scholar] [CrossRef]
- Wolny, H.L. Hormonal regulation of spermatogenesis. Wiad. Lek. 1978, 31, 185–188. [Google Scholar]
- Tesarik, J.; Greco, E.; Mendoza, C. Assisted reproduction with in-vitro-cultured testicular spermatozoa in cases of severe germ cell apoptosis: A pilot study. Hum. Reprod. 2001, 16, 2640–2645. [Google Scholar] [CrossRef][Green Version]
- Tesarik, J.; Guido, M.; Mendoza, C.; Greco, E. Human spermatogenesis in vitro: Respective effects of follicle- stimulating hormone and testosterone on meiosis, spermiogenesis, and sertoli cell apoptosis. J. Clin. Endocrinol. Metab. 1998, 83, 4467–4473. [Google Scholar] [CrossRef]
- Tesarik, J.; Greco, E.; Rienzi, L.; Ubaldi, F.; Guido, M.; Cohen-Bacrie, P.; Mendoza, C. Differentiation of spermatogenic cells during in-vitro culture of testicular biopsy samples from patients with obstructive azoospermia: Effect of recombinant follicle stimulating hormone. Hum. Reprod. 1998, 13, 2772–2781. [Google Scholar] [CrossRef]
- Sousa, M.; Cremades, N.; Alves, C.; Silva, J.; Barros, A. Developmental potential of human spermatogenic cells co-cultured with Sertoli cells. Hum. Reprod. 2002, 17, 161–172. [Google Scholar] [CrossRef][Green Version]
- Allan, C.M.; Garcia, A.; Spaliviero, J.; Zhang, F.-P.; Jimenez, M.; Huhtaniemi, I.; Handelsman, D.J. Complete Sertoli Cell Proliferation Induced by Follicle-Stimulating Hormone (FSH) Independently of Luteinizing Hormone Activity: Evidence from Genetic Models of Isolated FSH Action. Endocrinology 2004, 145, 1587–1593. [Google Scholar] [CrossRef]
- Vigier, M.; Weiss, M.; Perrard, M.H.; Godet, M.; Durand, P. The effects of FSH and of testosterone on the completion of meiosis and the very early steps of spermiogenesis of the rat: An in vitro study. J. Mol. Endocrinol. 2004, 33, 729–742. [Google Scholar] [CrossRef]
- Orth, J.M.; McGuinness, M.P.; Qiu, J.; Jester, W.F.; Li, L.H. Use of in vitro systems to study male germ cell development in neonatal rats. Theriogenology 1998, 49, 431–439. [Google Scholar] [CrossRef]
- Hikabe, O.; Hamazaki, N.; Nagamatsu, G.; Obata, Y.; Hirao, Y.; Hamada, N.; Shimamoto, S.; Imamura, T.; Nakashima, K.; Saitou, M.; et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature 2016, 539, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the Mouse Germ Cell Specification Pathway in Culture by Pluripotent Stem Cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science 2012, 338, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, M.; Tian, R.; Wang, Y.; Liu, L.; Zhu, Z.; Yang, S.; Yuan, Q.; Niu, M.; Yao, C.; et al. Derivation and propagation of spermatogonial stem cells from human pluripotent cells. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ye, S.; Liang, D.; Wang, P.; Fu, J.; Ma, Q.; Kong, R.; Shi, L.; Gong, X.; Chen, W.; et al. In Vitro Modeling of Human Germ Cell Development Using Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 509–523. [Google Scholar] [CrossRef]
- Sosa, E.; Chen, D.; Rojas, E.J.; Hennebold, J.D.; Peters, K.A.; Wu, Z.; Lam, T.N.; Mitchell, J.M.; Sukhwani, M.; Tailor, R.C.; et al. Differentiation of primate primordial germ cell-like cells following transplantation into the adult gonadal niche. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Pieri, N.C.; de Souza, A.F.; Botigelli, R.C.; de Figueiredo Pessôa, L.V.; Recchia, K.; Machado, L.S.; Glória, M.H.; de Castro, R.V.; Leal, D.F.; Neto, P.F.; et al. Porcine Primordial Germ Cell-Like Cells Generated from Induced Pluripotent Stem Cells under Different Culture Conditions. Stem Cell Rev. Rep. 2021, 1–18. [Google Scholar] [CrossRef]
- Raxita Singhal, D.K.S. Generation of Germ Cell-Like Cells and Oocyte-Like Cells from Goat Induced Pluripotent Stem Cells. J. Stem Cell Res. Ther. 2015, 5. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, M.; Yuan, Y.; Wang, X.; Fu, R.; Wan, H.; Xie, M.; Liu, M.; Guo, X.; Zheng, Y.; et al. Complete Meiosis from Embryonic Stem Cell-Derived Germ Cells in Vitro. Cell Stem Cell 2016, 18, 330–340. [Google Scholar] [CrossRef]
- Hayashi, K.; Hikabe, O.; Obata, Y.; Hirao, Y. Reconstitution of mouse oogenesis in a dish from pluripotent stem cells. Nat. Protoc. 2017, 12, 1733–1744. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cell Type | FSH Supplementation | Outcome | Author |
|---|---|---|---|---|
| Mouse | ESCs and iPSCs | FSH on in vitro growth media and in vitro maturation media | Mature oocytes/viable offspring | [259] |
| Human | ESCs and iPSCs | Recombinant human FSH in SSCLCs generation media | Human spermatogonial stem cells, further differentiation into spermatocytes and haploid cells | [262] |
| Mouse | ESCs | Spermatogenesis induction media contains FSH, testosterone, and bovine pituitary extract | Haploid spermatid-like cells/viable offspring | [267] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Recchia, K.; Jorge, A.S.; Pessôa, L.V.d.F.; Botigelli, R.C.; Zugaib, V.C.; de Souza, A.F.; Martins, D.d.S.; Ambrósio, C.E.; Bressan, F.F.; Pieri, N.C.G. Actions and Roles of FSH in Germinative Cells. Int. J. Mol. Sci. 2021, 22, 10110. https://doi.org/10.3390/ijms221810110
Recchia K, Jorge AS, Pessôa LVdF, Botigelli RC, Zugaib VC, de Souza AF, Martins DdS, Ambrósio CE, Bressan FF, Pieri NCG. Actions and Roles of FSH in Germinative Cells. International Journal of Molecular Sciences. 2021; 22(18):10110. https://doi.org/10.3390/ijms221810110
Chicago/Turabian StyleRecchia, Kaiana, Amanda Soares Jorge, Laís Vicari de Figueiredo Pessôa, Ramon Cesar Botigelli, Vanessa Cristiane Zugaib, Aline Fernanda de Souza, Daniele dos Santos Martins, Carlos Eduardo Ambrósio, Fabiana Fernandes Bressan, and Naira Caroline Godoy Pieri. 2021. "Actions and Roles of FSH in Germinative Cells" International Journal of Molecular Sciences 22, no. 18: 10110. https://doi.org/10.3390/ijms221810110
APA StyleRecchia, K., Jorge, A. S., Pessôa, L. V. d. F., Botigelli, R. C., Zugaib, V. C., de Souza, A. F., Martins, D. d. S., Ambrósio, C. E., Bressan, F. F., & Pieri, N. C. G. (2021). Actions and Roles of FSH in Germinative Cells. International Journal of Molecular Sciences, 22(18), 10110. https://doi.org/10.3390/ijms221810110