
Quest for the Molecular Basis of Improved Selective Toxicity of All-Trans Isomers of Aromatic Heptaene Macrolide Antifungal Antibiotics

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
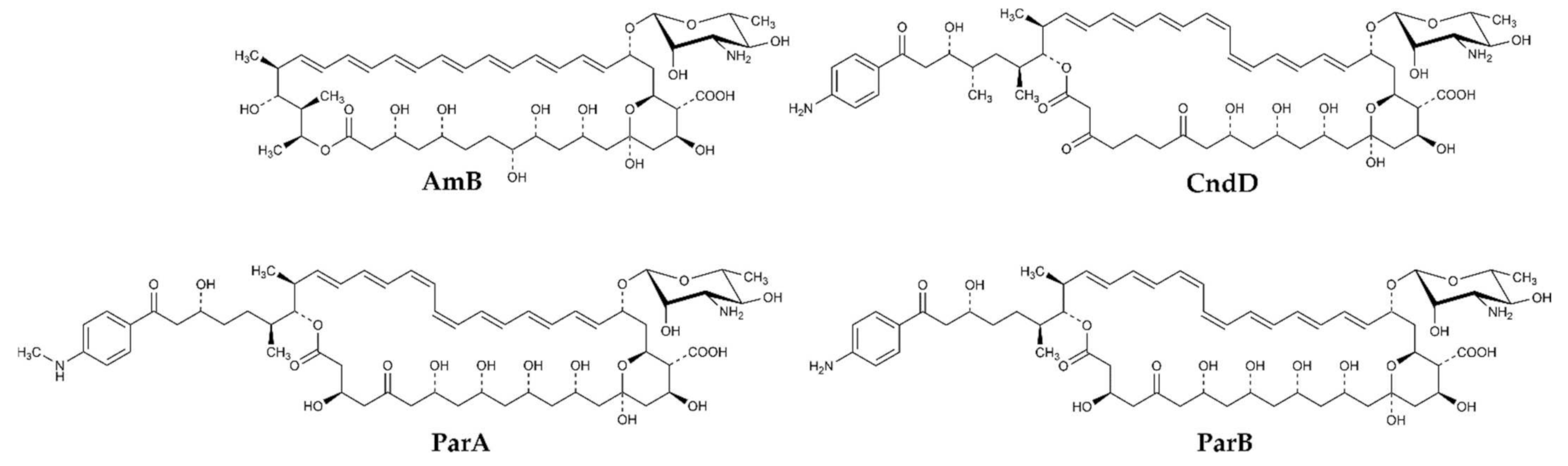
2.1. Preparation and Confirmation of Identity of Candicidin D, Partricin A, Partricin B and Their All-Trans Isomers
2.2. Antifungal In Vitro Activity, Hemolytic Properties and Selective Toxicity of CndD, ParA, ParB, isoCndD, isoParA and isoParB
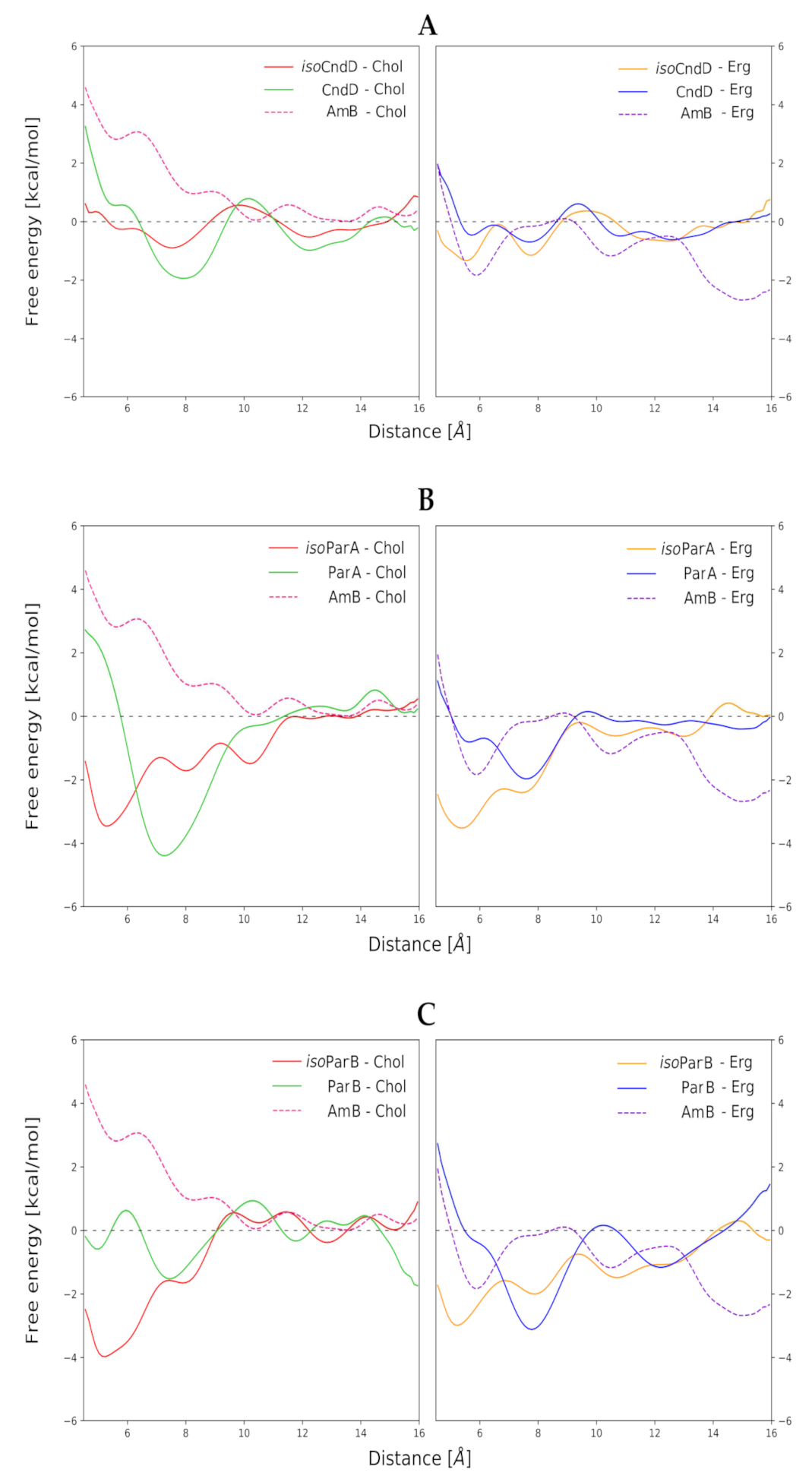
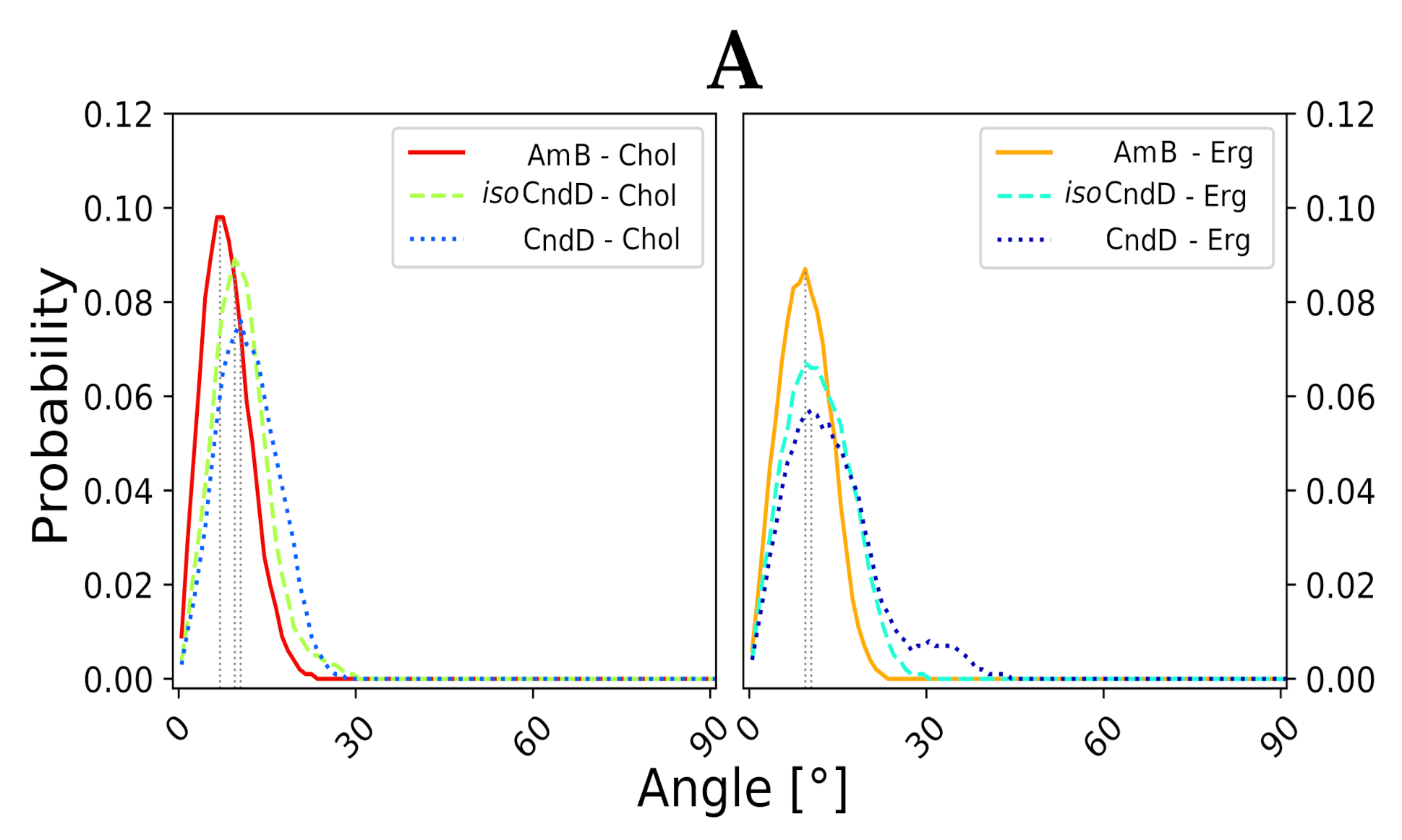
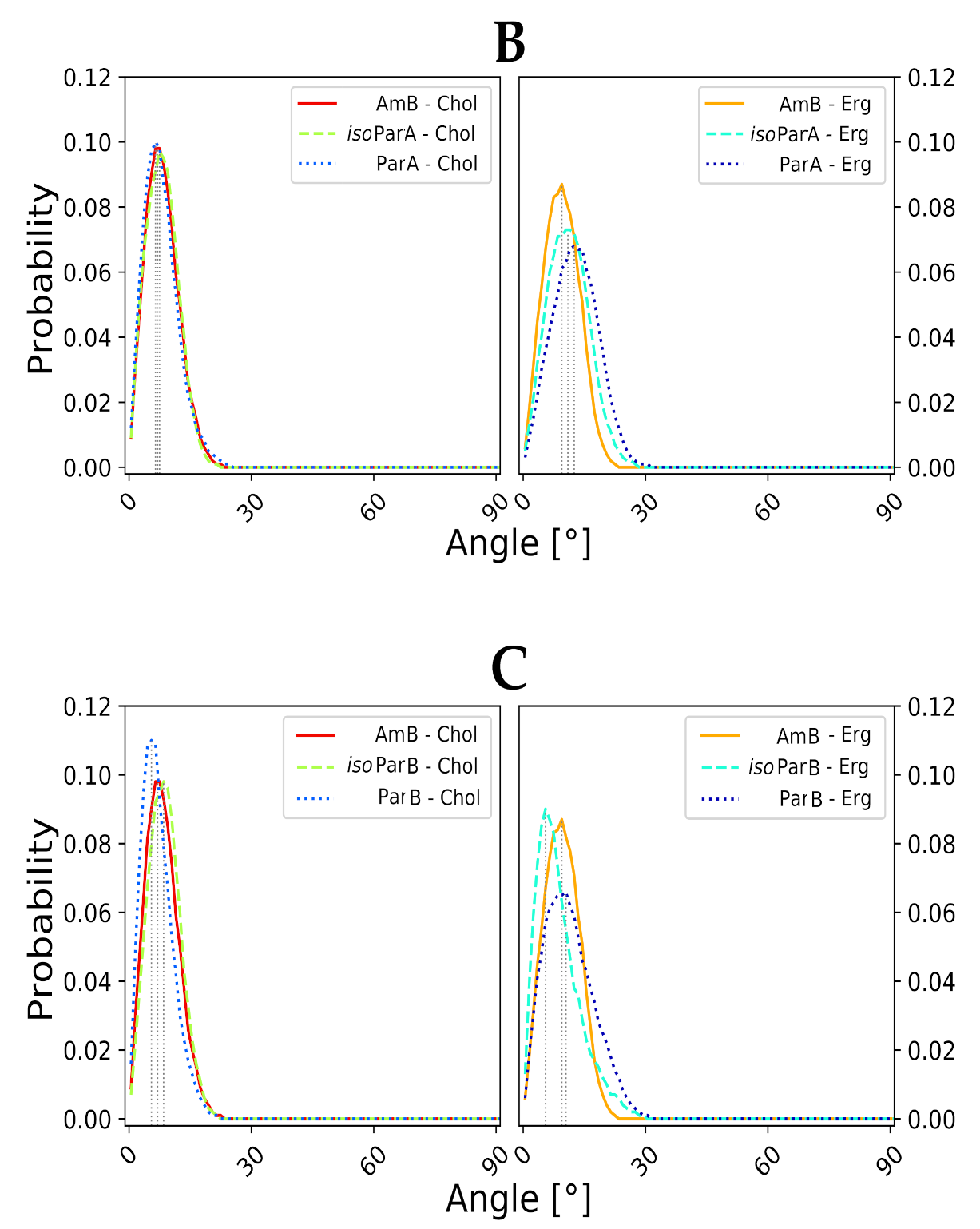
2.3. Molecular Modeling of AH:Sterol Interactions
3. Discussion
4. Materials and Methods
4.1. Preparation and Isolation of Compounds
4.1.1. Isolation of ParA, ParB and CndD from Antibiotic Complexes
4.1.2. Photochemical Cis-Trans → All-Trans Isomerisation of Components of Aureofacin and Candicidin Complexes and Isolation of All-Trans Isomers of ParA, ParB and CndD
4.1.3. Structural Analysis
NMR Experimental
HPLC-MS Analysis
4.2. Microbial Strains and Culture Conditions
4.3. Antifungal In Vitro Activity Determination
4.4. Hemolytic Activity Determination
4.5. Molecular Modelling
4.5.1. Parametrization
4.5.2. System Preparation
4.5.3. Molecular Dynamics Simulations
4.5.4. Umbrella Sampling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hamilton-Miller, J.M. Chemistry and biology of polyene macrolide antibiotics. Bacteriol. Rev. 1973, 37, 166–196. [Google Scholar] [CrossRef]
- Cavassin, S.B.; Baứ-Carneiro, J.L.; Vilas-Boas, R.R.; Queiroz-Telles, F. Sixty years of Amphotericin B: An overview of the main antifungal agent used to treat invasive fungal infections. Infect. Dis. Ther. 2021, 10, 115–147. [Google Scholar] [CrossRef]
- Oroshnik, W.; Mebane, A.D. The Polyene Antifungal Antibiotics. In Progress in the Chemistry of Organic Natural Products/Progrès Dans La Chimie Des Substances Organiques Naturelles; Springer: Berlin/Heidelberg, Germany, 1963; pp. 17–79. [Google Scholar] [CrossRef]
- Bagiński, M.; Czub, J. Amphotericin B and its new derivatives-mode of action. Curr. Drug Metab. 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.-T.; Zong, C.; Dagher, Z.; Hui, J.; Li, J.; Zhan, Y.; Zhang, M.; Mansour, M.K.; Cheng, J.-X. Polarization-sensitive stimulated Raman scattering imaging resolves amphotericin B orientation in Candida membrane. Sci. Adv. 2021, 7, eabd5230. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.M.; Clay, M.C.; Cioffi, A.G.; Diaz, K.A.; Hisao, G.S.; Tuttle, M.D.; Nieuwkoop, A.; Comellas, G.; Maryum, N.; Wang, S.; et al. Amphotericin forms an extramembranous and fungicidal sterol sponge. Nat. Chem. Biol. 2014, 10, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, J.; Li, X.; Xiao, E.; Lange, J.D.; Rienstra, C.M.; Burke, M.D.; Mitchell, D.A. Sterol Sponge Mechanism Is Conserved for Glycosylated Polyene Macrolides. ACS Central Sci. 2021, 7, 781–791. [Google Scholar] [CrossRef]
- Neumann, A.; Bagiński, M.; Czub, J. How Do Sterols Determine the Antifungal Activity of Amphotericin B? Free Energy of Binding between the Drug and Its Membrane Targets. J. Am. Chem. Soc. 2010, 132, 18266–18272. [Google Scholar] [CrossRef]
- Legrand, P.; Romero, E.A.; Cohen, B.E.; Bolard, J. Effects of aggregation and solvent on the toxicity of amphotericin B to human erythrocytes. Antimicrob. Agents Chemother. 1992, 36, 2518–2522. [Google Scholar] [CrossRef]
- Paquet, V.; Volmer, A.A.; Carreira, E.M. Synthesis and in vitro biological properties of novel cationic derivatives of amphotericin B. Chemistry 2008, 14, 2465–2481. [Google Scholar] [CrossRef]
- Wilcock, B.C.; Endo, M.M.; Uno, B.E.; Burke, M.D. C2′-OH of amphotericin B plays an important role in binding the primary sterol of human cells but not yeast cells. J. Am. Chem. Soc. 2013, 135, 8488–8491. [Google Scholar] [CrossRef]
- Antillón, A.; de Vries, A.H.; Espinosa-Caballero, M.; Falcón-González, J.M.; Flores Romero, D.; González-Damián, J.; Jiménez-Montejo, F.E.; León-Buitimea, A.; López-Ortiz, M.; Magaña, R.; et al. An Amphotericin B derivative equally potent to Amphotericin B and with increased safety. PLoS ONE 2016, 11, e0162171. [Google Scholar]
- Borowski, E.; Salewska, N.; Boros-Majewska, J.; Serocki, M.; Chabowska, I.; Milewska, M.J.; Ziętkowski, D.; Milewski, S. The substantial improvement of Amphotericin B selective toxicity upon modification of mycosamine with bulky substituents. Med. Chem. 2020, 16, 128–139. [Google Scholar] [CrossRef]
- Tevyashova, A.N.; Bychkova, E.N.; Solovieva, S.E.; Zatonsky, G.V.; Grammatikova, N.E.; Isakova, E.B.; Mirchink, E.P.; Treshchalin, I.D.; Pereverzeva, E.R.; Bykov, E.E.; et al. Discovery of amphamide, a drug candidate for the second generation of polyene antibiotics. ACS Infect. Dis. 2020, 14, 2029–2044. [Google Scholar] [CrossRef]
- Cybulska, B.; Zimiński, T.; Borowski, E.; Gary-Bobo, C.M. The influence of electric charge of aromatic heptaene macrolide antibiotics on their activity on biological and lipidic model membranes. Mol. Pharmacol. 1983, 24, 270–276. [Google Scholar] [PubMed]
- Szczeblewski, P.; Laskowski, T.; Bałka, A.; Borowski, E.; Milewski, S. Light-Induced Transformation of the Aromatic Heptaene Antifungal Antibiotic Candicidin D into Its All-Trans Isomer. J. Nat. Prod. 2018, 81, 1540–1545. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; CLSI standard M27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Sowiński, P.; Pawlak, J.; Borowski, E.; Gariboldi, P. ChemInform Abstract: Stereostructure of Gedamycin (I). ChemInform 1995, 26. [Google Scholar] [CrossRef]
- Sowiński, P.; Gariboldi, P.; Czerwiński, A.; Borowski, E. The structure of vacidin A, an aromatic heptaene macrolide antibiotic. I. Complete assignment of the 1H NMR spectrum and geometry of the polyene chromophore. J. Antibiot. 1989, 42, 1631–1638. [Google Scholar] [CrossRef]
- Sowiński, P.; Gariboldi, P.; Pawlak, J.K.; Borowski, E. The structure of vacidin A, an aromatic heptaene macrolide antibiotic. II. Stereochemistry of the antibiotic. J. Antibiot. 1989, 42, 1639–1642. [Google Scholar] [CrossRef]
- Szwarc, K.; Szczeblewski, P.; Sowiński, P.; Borowski, E.; Pawlak, J. The stereostructure of candicidin D. J. Antibiot. 2015, 68, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Ermishkin, L.N.; Kasumov, K.M.; Potzeluyev, V.M. Single ionic channels induced in lipid bilayers by polyene antibiotics amphotericin B and nystatine. Nature 1976, 262, 698–699. [Google Scholar] [CrossRef]
- Grudziński, W.; Sagan, J.; Welc, R.; Luchowski, R.; Gruszecki, W.I. Molecular organization, localization and orientation of antifungal antibiotic amphotericin B in a single lipid bilayer. Sci. Rep. 2016, 6, 32780. [Google Scholar] [CrossRef] [PubMed]
- Ślisz, M.; Cybulska, B.; Mazerski, J.; Grzybowska, J.; Borowski, E. Studies of the Effects of Antifungal Cationic Derivatives of Amphotericin B on Human Erythrocytes. J. Antibiot. 2004, 57, 669–678. [Google Scholar] [CrossRef][Green Version]
- Klauda, J.B.; Venable, R.M.; Freites, J.A.; O’Connor, J.W.; Tobias, D.J.; Mondragon-Ramirez, C.; Vorobyov, I.; MacKerell, A.; Pastor, R.W. Update of the CHARMM All-Atom Additive Force Field for Lipids: Validation on Six Lipid Types. J. Phys. Chem. B 2010, 114, 7830–7843. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 09, Revision D.01; Gaussian Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Mayne, C.G.; Saam, J.; Schulten, K.; Tajkhorshid, E.; Gumbart, J.C. Rapid parameterization of small molecules using the force field toolkit. J. Comput. Chem. 2013, 34, 2757–2770. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Dávila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M.; et al. CHARMM-GUI membrane builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Tribello, G.A.; Bonomi, M.; Branduardi, D.; Camilloni, C.; Bussi, G. PLUMED 2: New feathers for an old bird. Comput. Phys. Commun. 2014, 185, 604–613. [Google Scholar] [CrossRef]
- Laio, A.; Parrinello, M. Escaping free-energy minima. Proc. Natl. Acad. Sci. USA 2002, 99, 12562–12566. [Google Scholar] [CrossRef] [PubMed]
- Darden, T.A.; York, D.M.; Pedersen, L.G. Particle mesh Ewald: AnN⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Kästner, J. Umbrella Sampling. WIREs Comput. Mol. Sci. 2011, 1, 932–942. [Google Scholar] [CrossRef]
- Kumar, S.; Rosenberg, J.M.; Bouzida, D.; Swendsen, R.; Kollman, P.A. The weighted histogram analysis method for free-energy calculations on biomolecules. I. The method. J. Comput. Chem. 1992, 13, 1011–1021. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CndD | isoCndD | ParA | isoParA | ParB | isoParB | AmB |
|---|---|---|---|---|---|---|---|
| Microorganism | MIC (µg mL−1) | ||||||
| C. albicans | 0.25 | 0.5 | 0.25 | 1.0 | 0.25 | 0.5 | 0.25 |
| C. glabrata | 0.5 | 0.5 | 0.125 | 0.5 | 0.125 | 0.25 | 1.0 |
| C. krusei | 0.25 | 0.25 | 0.125 | 0.5 | 0.5 | 0.5 | 0.5 |
| C. parapsilosis | 0.25 | 0.06 | 0.06 | 0.125 | 0.25 | 0.125 | 0.5 |
| C. dubliniensis | 0.25 | 0.125 | 0.125 | 0.5 | 0.25 | 0.25 | 0.25 |
| C. guilliermondii | 1.0 | 0.5 | 0.5 | 0.5 | 1.0 | 1.0 | 0.25 |
| C. famata | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| C. rugosa | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 2.0 |
| S. cerevisiae | 0.5 | 0.25 | 0.125 | 0.5 | 0.125 | 0.25 | 0.5 |
| Compound | CndD | isoCndD | ParA | isoParA | ParB | isoParB | AmB |
|---|---|---|---|---|---|---|---|
| Microorganism | MIC (µg mL−1) | ||||||
| C. albicans | 0.003 | 0.003 | 0.002 | 0.006 | 0.002 | 0.003 | 0.06 |
| C. glabrata | 0.006 | 0.003 | 0.002 | 0.006 | 0.002 | 0.006 | 0.25 |
| C. krusei | 0.006 | 0.003 | 0.002 | 0.006 | 0.003 | 0.003 | 0.25 |
| C. parapsilosis | 0.01 | 0.006 | 0.003 | 0.006 | 0.006 | 0.006 | 0.125 |
| C. dubliniensis | 0.002 | 0.002 | 0.001 | 0.006 | 0.002 | 0.002 | 0.06 |
| C. guilliermondii | 0.03 | 0.03 | 0.006 | 0.03 | 0.01 | 0.01 | 0.125 |
| C. famata | 0.01 | 0.03 | 0.01 | 0.05 | 0.01 | 0.05 | 0.25 |
| C. rugosa | 0.03 | 0.03 | 0.03 | 0.05 | 0.03 | 0.03 | 0.125 |
| S. cerevisiae | 0.006 | 0.006 | 0.002 | 0.001 | 0.003 | 0.003 | 0.06 |
| Compound | CndD | isoCndD | ParA | isoParA | ParB | isoParB | AmB |
|---|---|---|---|---|---|---|---|
| EH50 (µg mL−1) | 7.53 ± 0.82 | 16.67 ± 1.40 | 1.28 ± 0.68 | >20 | 0.66 ± 0.09 | 9.64 ± 1.79 | 3.46 ± 0.15 |
| Compound | CndD | isoCndD | ParA | isoParA | ParB | isoParB | AmB |
|---|---|---|---|---|---|---|---|
| STI | 30.12 | 33.34 | 5.12 | >20 | 2.64 | 19.28 | 13.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borzyszkowska-Bukowska, J.; Górska, J.; Szczeblewski, P.; Laskowski, T.; Gabriel, I.; Jurasz, J.; Kozłowska-Tylingo, K.; Szweda, P.; Milewski, S. Quest for the Molecular Basis of Improved Selective Toxicity of All-Trans Isomers of Aromatic Heptaene Macrolide Antifungal Antibiotics. Int. J. Mol. Sci. 2021, 22, 10108. https://doi.org/10.3390/ijms221810108
Borzyszkowska-Bukowska J, Górska J, Szczeblewski P, Laskowski T, Gabriel I, Jurasz J, Kozłowska-Tylingo K, Szweda P, Milewski S. Quest for the Molecular Basis of Improved Selective Toxicity of All-Trans Isomers of Aromatic Heptaene Macrolide Antifungal Antibiotics. International Journal of Molecular Sciences. 2021; 22(18):10108. https://doi.org/10.3390/ijms221810108
Chicago/Turabian StyleBorzyszkowska-Bukowska, Julia, Justyna Górska, Paweł Szczeblewski, Tomasz Laskowski, Iwona Gabriel, Jakub Jurasz, Katarzyna Kozłowska-Tylingo, Piotr Szweda, and Sławomir Milewski. 2021. "Quest for the Molecular Basis of Improved Selective Toxicity of All-Trans Isomers of Aromatic Heptaene Macrolide Antifungal Antibiotics" International Journal of Molecular Sciences 22, no. 18: 10108. https://doi.org/10.3390/ijms221810108
APA StyleBorzyszkowska-Bukowska, J., Górska, J., Szczeblewski, P., Laskowski, T., Gabriel, I., Jurasz, J., Kozłowska-Tylingo, K., Szweda, P., & Milewski, S. (2021). Quest for the Molecular Basis of Improved Selective Toxicity of All-Trans Isomers of Aromatic Heptaene Macrolide Antifungal Antibiotics. International Journal of Molecular Sciences, 22(18), 10108. https://doi.org/10.3390/ijms221810108