
Insights Into the Micelle-Induced β-Hairpin-to-α-Helix Transition of a LytA-Derived Peptide by Photo-CIDNP Spectroscopy

 ,
, 

Abstract
1. Introduction
2. Results
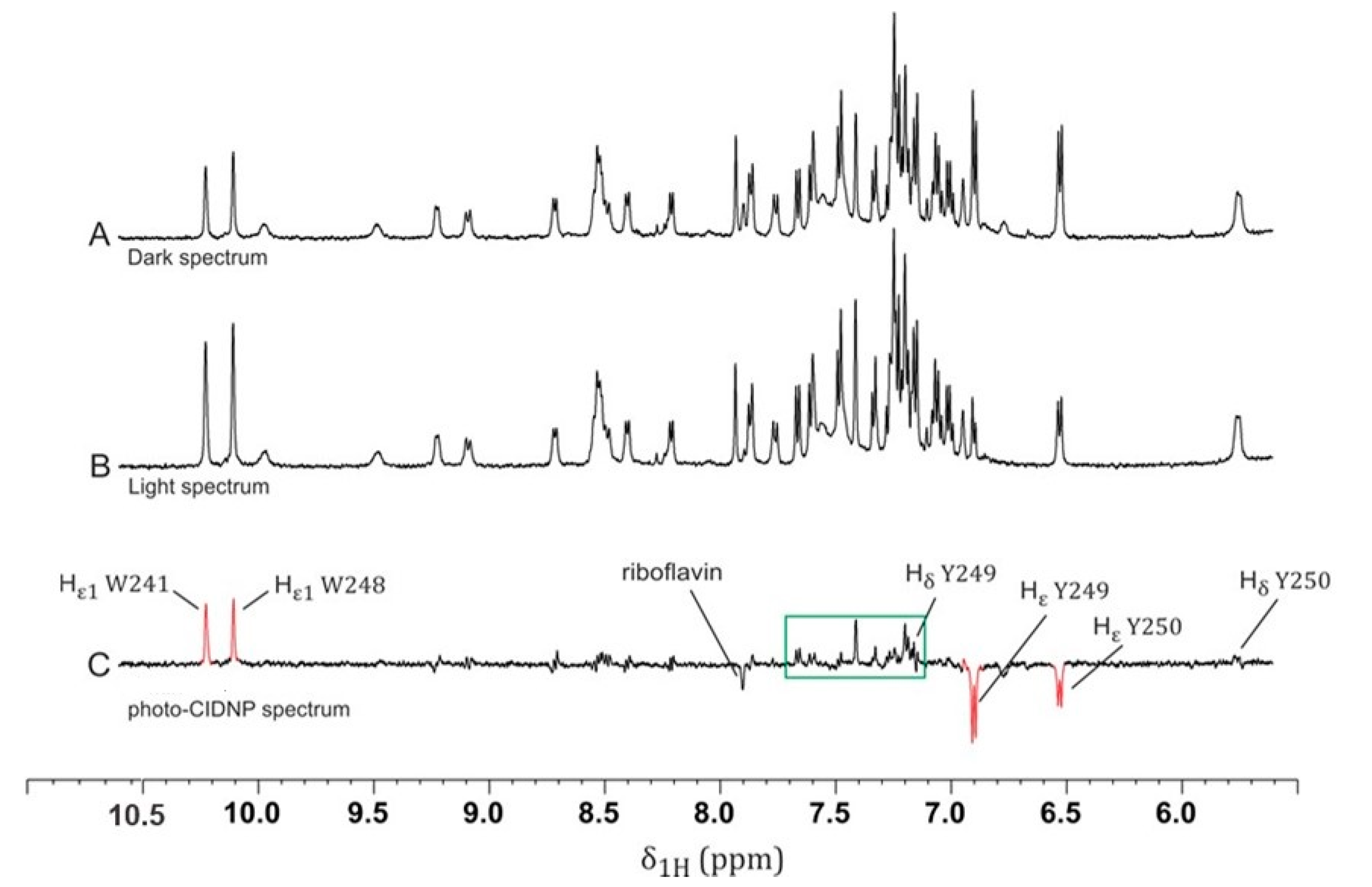
2.1. Photo-CIDNP Experiments of LytA239–252 Peptide in the Absence of DPC Micelles
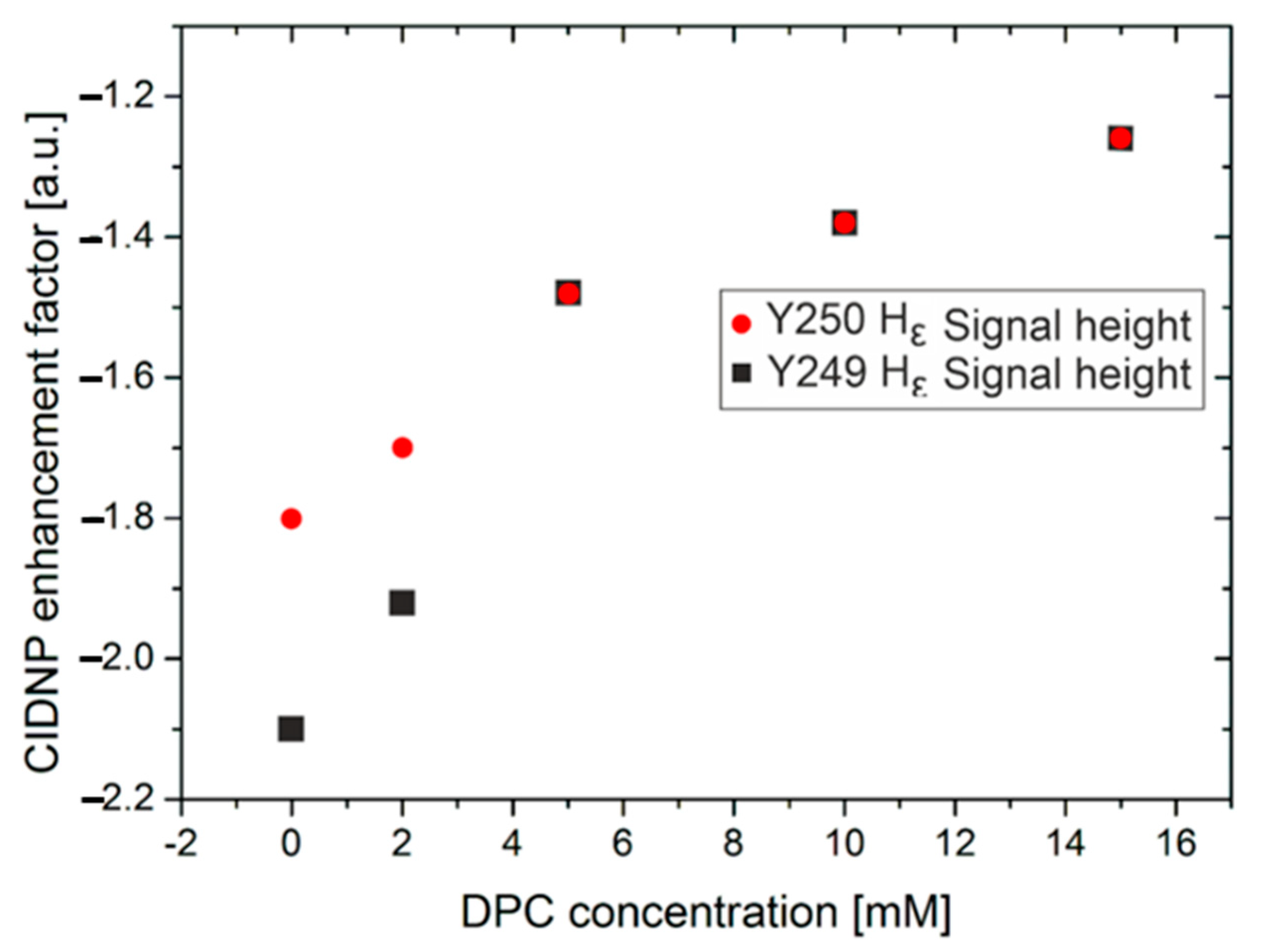
2.2. Photo-CIDNP Experiments of LytA239–252 Peptide in the Presence of DPC Micelles
2.3. Optimization of Light Intensity in NMR Active Volume to Observe Cross-Polarization Effects
2.4. Cross-Polarization Signals for Hδ of Y249 and Y250
3. Discussion
4. Materials and Methods
4.1. Light Sources
4.1.1. Light-Emitting Diode (LED)
4.1.2. Laser Diode
4.2. NMR Sample Preparation
4.3. NMR Experiments
4.3.1. Method A
4.3.2. Method B
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Berntsson, O.; Diensthuber, R.P.; Panman, M.R.; Bjorling, A.; Gustavsson, E.; Hoernke, M.; Hughes, A.J.; Henry, L.; Niebling, S.; Takala, H.; et al. Sequential Conformational Transitions and α-Helical Supercoiling Regulate a Sensor Histidine Kinase. Nat. Commun. 2017, 8, 284. [Google Scholar] [CrossRef] [PubMed]
- Newton, K. RIPK1 and RIPK3: Critical Regulators of Inflammation and Cell Death. Trends Cell Biol. 2015, 25, 347–353. [Google Scholar] [CrossRef]
- Carrell, R.W.; Gooptu, B. Conformational Changes and Disease—Serpins, Prions and Alzheimer’s. Curr. Opin. Struct. Biol. 1998, 8, 799–809. [Google Scholar] [CrossRef]
- Cohen, F.E. Protein Misfolding and Prion Diseases. J. Mol. Biol. 1999, 293, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Rochet, J.-C.; Lansbury, P.T.J. Amyloid Fibrillogenesis: Themes and Variations. Curr. Opin. Struct. Biol. 2000, 10, 60–68. [Google Scholar] [CrossRef]
- Wälti, M.A.; Ravotti, F.; Arai, H.; Glabe, C.G.; Wall, J.S.; Böckmann, A.; Güntert, P.; Meier, B.H.; Riek, R. Atomic-Resolution Structure of a Disease-Relevant Aβ(1-42) Amyloid Fibril. Proc. Natl. Acad. Sci. USA 2016, 113, E4976–E4984. [Google Scholar] [CrossRef]
- Colvin, M.T.; Silvers, R.; Ni, Q.Z.; Can, T.V.; Sergeyev, I.; Rosay, M.; Donovan, K.J.; Michael, B.; Wall, J.; Linse, S.; et al. Atomic Resolution Structure of Monomorphic Aβ42 Amyloid Fibrils. J. Am. Chem. Soc. 2016, 138, 9663–9674. [Google Scholar] [CrossRef]
- Baneyx, F.; Matthaei, J.F. Self-Assembled Two-Dimensional Protein Arrays in Bionanotechnology: From S-Layers to Designed Lattices. Curr. Opin. Biotechnol. 2014, 28, 39–45. [Google Scholar] [CrossRef]
- Zamora-Carreras, H.; Maestro, B.; Sanz, J.M.; Jimenez, M.A. Turncoat Polypeptides: We Adapt to Our Environment. ChemBioChem 2020, 21, 432–441. [Google Scholar] [CrossRef]
- Lella, M.; Mahalakshmi, R. Metamorphic Proteins: Emergence of Dual Protein Folds from One Primary Sequence. Biochemistry 2017, 56, 2971–2984. [Google Scholar] [CrossRef]
- Dishman, A.F.; Volkman, B.F. Unfolding the Mysteries of Protein Metamorphosis. ACS Chem. Biol. 2018, 13, 1438–1446. [Google Scholar] [CrossRef]
- Ambroggio, X.I.; Kuhlman, B. Design of Protein Conformational Switches. Curr. Opin. Struct. Biol. 2006, 16, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Ciani, B.; Hutchinson, E.G.; Sessions, R.B.; Woolfson, D.N. A Designed System for Assessing How Sequence Affects α to β Conformational Transitions in Proteins. J. Biol. Chem. 2002, 277, 10150–10155. [Google Scholar] [CrossRef]
- Kupfer, L.; Hinrichs, W.; Groschup, M.H. Prion Protein Misfolding. Curr. Mol. Med. 2009, 9, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Saucede, L.; Dos Santosa, S.; Chandravarkar, A.; Mandal, B.; Mimna, R.; Murat, K.; Camus, M.-S.; Berard, J.; Grouzmann, E.; Adrian, M.; et al. Switch-Peptides: From Conformational Studies to Alzheimer’s Disease. Chimia 2006, 60, 199–202. [Google Scholar] [CrossRef][Green Version]
- Ji, H.-F.; Gao, H.; Buchapudi, K.R.; Yang, X.; Xu, X.; Schulte, M.K. Microcantilever Biosensors Based on Conformational Change of Proteins. Analyst 2008, 133, 434–443. [Google Scholar] [CrossRef]
- Maestro, B.; Sanz, J.M. Choline Binding Proteins from Streptococcus pneumoniae: A Dual Role as Enzybiotics and Targets for the Design of New Antimicrobials. Antibiotics 2016, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Carreras, H.; Maestro, B.; Strandberg, E.; Ulrich, A.S.; Sanz, J.M.; Jiménez, M.Á. Micelle-Triggered β-Hairpin to α-Helix Transition in a 14-Residue Peptide from a Choline-Binding Repeat of the Pneumococcal Autolysin LytA. Chem. Eur. J. 2015, 21, 8076–8089. [Google Scholar] [CrossRef]
- Zamora-Carreras, H.; Maestro, B.; Strandberg, E.; Ulrich, A.S.; Sanz, J.M.; Jimenez, M.Á. Roles of Amphipathicity and Hydrophobicity in the Micelle-Driven Structural Switch of a 14-Mer Peptide Core from a Choline-Binding Repeat. Chem. Eur. J. 2018, 24, 5825–5839. [Google Scholar] [CrossRef]
- Kuhn, L.T. Photo-CIDNP NMR Spectroscopy of Amino Acids and Proteins. In Hyperpolarization Methods in Nmr Spectroscopy; Kuhn, L.T., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 338, pp. 229–300. [Google Scholar] [CrossRef]
- Zetta, L.; Kaptein, R.; Hore, P. A Photo-Cidnp Investigation of Tyrosine Mobility and Exposure in Human β-Endorphin in the Presence of Phospholipid Micelles. FEBS Lett. 1982, 145, 277–280. [Google Scholar] [CrossRef]
- Hors, P.J.; Egmond, M.R.; Edzes, H.T.; Kaptein, R. Cross-Relaxation Effects in the Photo-Cidnp Spectra of Amino-Acids and Proteins. J. Magn. Reson. 1982, 49, 122–150. [Google Scholar] [CrossRef]
- Feldmeier, C.; Bartling, H.; Riedle, E.; Gschwind, R.M. LED Based NMR Illumination Device for Mechanistic Studies on Photochemical Reactions—Versatile and Simple, yet Surprisingly Powerful. J. Magn. Reson. 2013, 232, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, P.; Lokesh, N.; Gschwind, R.M. Combination of Illumination and High Resolution NMR Spectroscopy: Key Features and Practical Aspects, Photochemical Applications, and New Concepts. Prog. Nucl. Magn. Reson. Spectrosc. 2019, 114–115, 86–134. [Google Scholar] [CrossRef] [PubMed]
- Seegerer, A.; Nitschke, P.; Gschwind, R.M. Combined in Situ Illumination-NMR-UV/Vis Spectroscopy: A New Mechanistic Tool in Photochemistry. Angew. Chem. Int. Ed. 2018, 57, 7493–7497. [Google Scholar] [CrossRef]
- Mompean, M.; Sanchez-Donoso, R.M.; de la Hoz, A.; Saggiomo, V.; Velders, A.H.; Gomez, M.V. Pushing Nuclear Magnetic Resonance Sensitivity Limits with Microfluidics and Photochemically Induced Dynamic Nuclear Polarization. Nat. Commun. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Mok, K.H.; Hore, P.J. Photo-CIDNP NMR Methods for Studying Protein Folding. Methods 2004, 34, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Goez, M. An Introduction to Chemically Induced Dynamic Nuclear Polarization. Concepts Magn. Reson. 1995, 7, 69–86. [Google Scholar] [CrossRef]
- Lin, P.-C. Assessment of Chemical Exchange in Tryptophan-Albumin Solution through (19)F Multicomponent Transverse Relaxation Dispersion Analysis. J. Biomol. NMR 2015, 62, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.L.; Moraes, A.; Nakaie, C.R.; Almeida, F.C.L.; Schreier, S.; Valente, A.P. Structural and Dynamic Insights of the Interaction between Tritrpticin and Micelles: An NMR Study. Biophys. J. 2016, 111, 2676–2688. [Google Scholar] [CrossRef]
- Gomez, M.V.; Juan, A.; Jimenez-Marquez, F.; de la Hoz, A.; Velders, A.H. Illumination of Nanoliter-NMR Spectroscopy Chips for Real-Time Photochemical Reaction Monitoring. Anal. Chem. 2018, 90, 1542–1546. [Google Scholar] [CrossRef]
- Ji, Y.; DiRocco, D.A.; Kind, J.; Thiele, C.M.; Gschwind, R.M.; Reibarkh, M. LED-Illuminated NMR Spectroscopy: A Practical Tool for Mechanistic Studies of Photochemical Reactions. ChemPhotoChem 2019, 3, 984–992. [Google Scholar] [CrossRef]
- Abdiaj, I.; Fontana, A.; Gomez, M.V.; de la Hoz, A.; Alcazar, J. Visible-Light-Induced Nickel-Catalyzed Negishi Cross-Couplings by Exogenous-Photosensitizer-Free Photocatalysis. Angew. Chem. Int. Ed. 2018, 57, 8473–8477. [Google Scholar] [CrossRef] [PubMed]
- Okuno, Y.; Cavagnero, S. Photochemically Induced Dynamic Nuclear Polarization: Basic Principles and Applications. Emagres 2017, 6, 283–314. [Google Scholar] [CrossRef]
- Dekanter, F.; Kaptein, R. Cidnp Transfer Via Nuclear Dipolar Relaxation and Spin-Spin Coupling. Chem. Phys. Lett. 1979, 62, 421–426. [Google Scholar] [CrossRef]
- Nguyen, P.H.; Ramamoorthy, A.; Sahoo, B.R.; Zheng, J.; Faller, P.; Straub, J.E.; Dominguez, L.; Shea, J.-E.; Dokholyan, N.V.; De Simone, A.; et al. Amyloid Oligomers: A Joint Experimental/Computational Perspective on Alzheimer’s Disease, Parkinson’s Disease, Type II Diabetes, and Amyotrophic Lateral Sclerosis. Chem. Rev. 2021, 121, 2545–2647. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| DPC | Hδ Y250 | Hδ Y249 |
|---|---|---|
| 0 mM | +1.78 | +9.55 |
| 2 mM | −1.33 | +6.65 |
| DPC | ρ Y249 | ρ Y250 |
|---|---|---|
| 0 mM | −0.25 | −0.21 |
| 2 mM | −0.22 | +0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez, M.V.; Ruiz-Castañeda, M.; Nitschke, P.; Gschwind, R.M.; Jiménez, M.A. Insights Into the Micelle-Induced β-Hairpin-to-α-Helix Transition of a LytA-Derived Peptide by Photo-CIDNP Spectroscopy. Int. J. Mol. Sci. 2021, 22, 6666. https://doi.org/10.3390/ijms22136666
Gomez MV, Ruiz-Castañeda M, Nitschke P, Gschwind RM, Jiménez MA. Insights Into the Micelle-Induced β-Hairpin-to-α-Helix Transition of a LytA-Derived Peptide by Photo-CIDNP Spectroscopy. International Journal of Molecular Sciences. 2021; 22(13):6666. https://doi.org/10.3390/ijms22136666
Chicago/Turabian StyleGomez, M. Victoria, Margarita Ruiz-Castañeda, Philipp Nitschke, Ruth M. Gschwind, and M. Angeles Jiménez. 2021. "Insights Into the Micelle-Induced β-Hairpin-to-α-Helix Transition of a LytA-Derived Peptide by Photo-CIDNP Spectroscopy" International Journal of Molecular Sciences 22, no. 13: 6666. https://doi.org/10.3390/ijms22136666
APA StyleGomez, M. V., Ruiz-Castañeda, M., Nitschke, P., Gschwind, R. M., & Jiménez, M. A. (2021). Insights Into the Micelle-Induced β-Hairpin-to-α-Helix Transition of a LytA-Derived Peptide by Photo-CIDNP Spectroscopy. International Journal of Molecular Sciences, 22(13), 6666. https://doi.org/10.3390/ijms22136666