Effects of an Aquaporin 4 Inhibitor, TGN-020, on Murine Diabetic Retina

 ,
,
Abstract
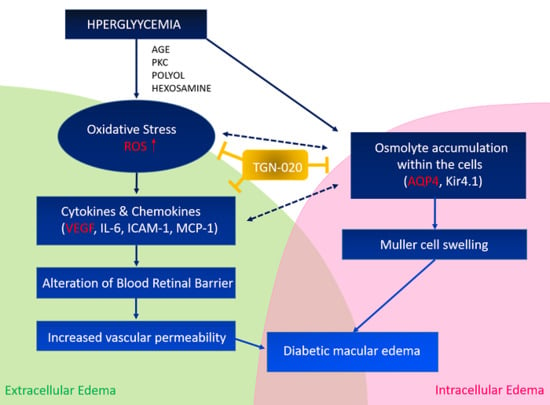
1. Introduction
2. Results
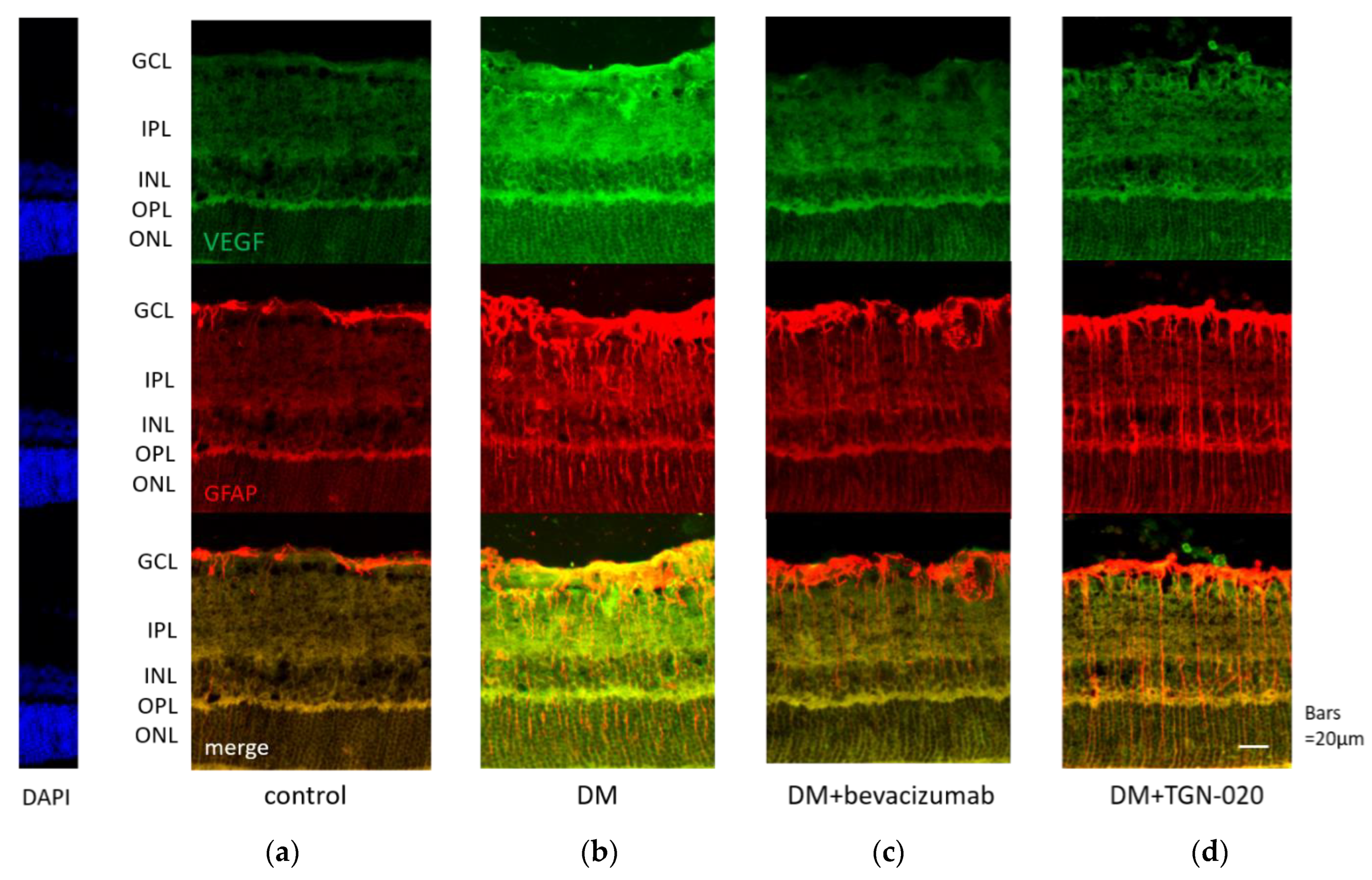
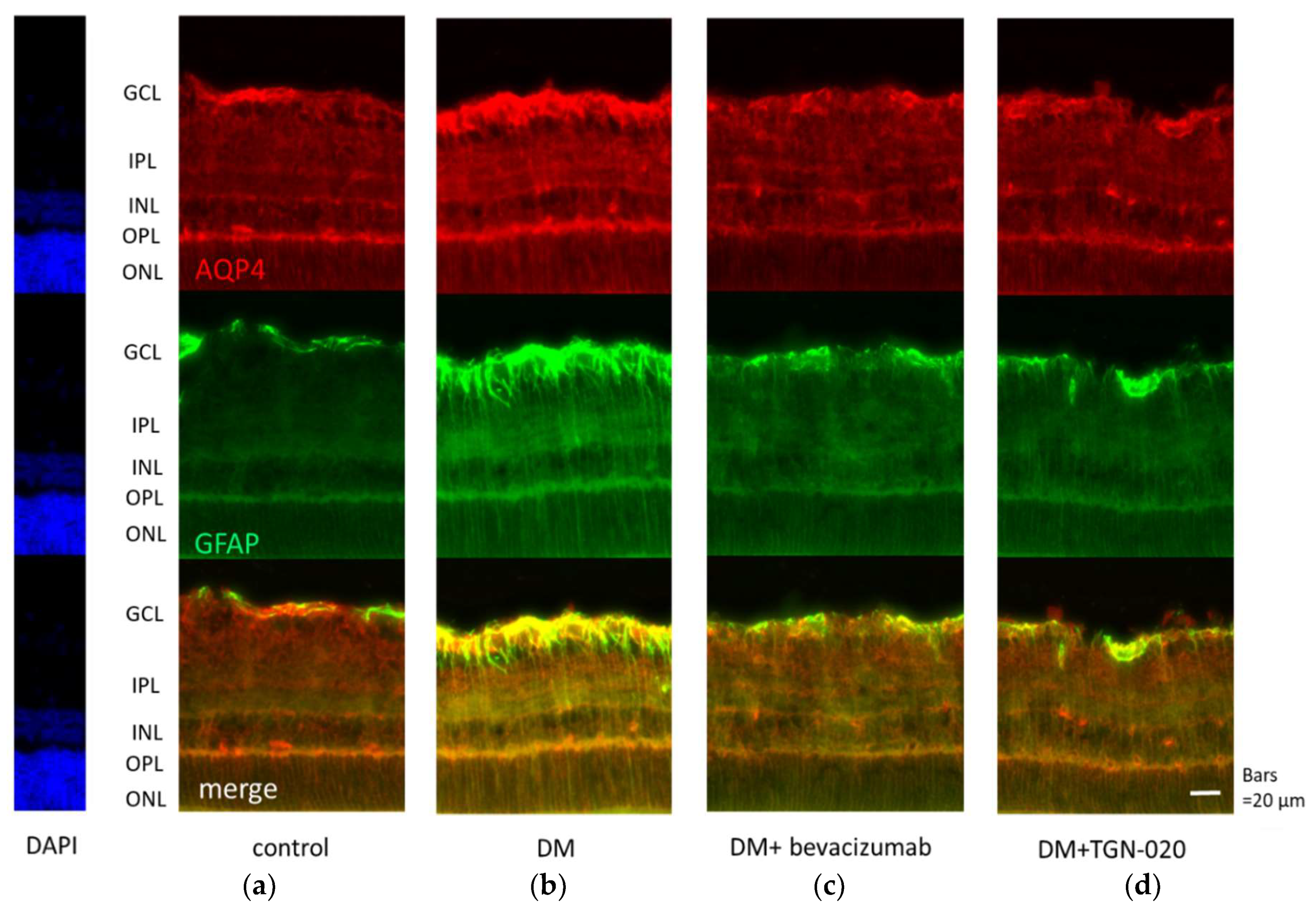
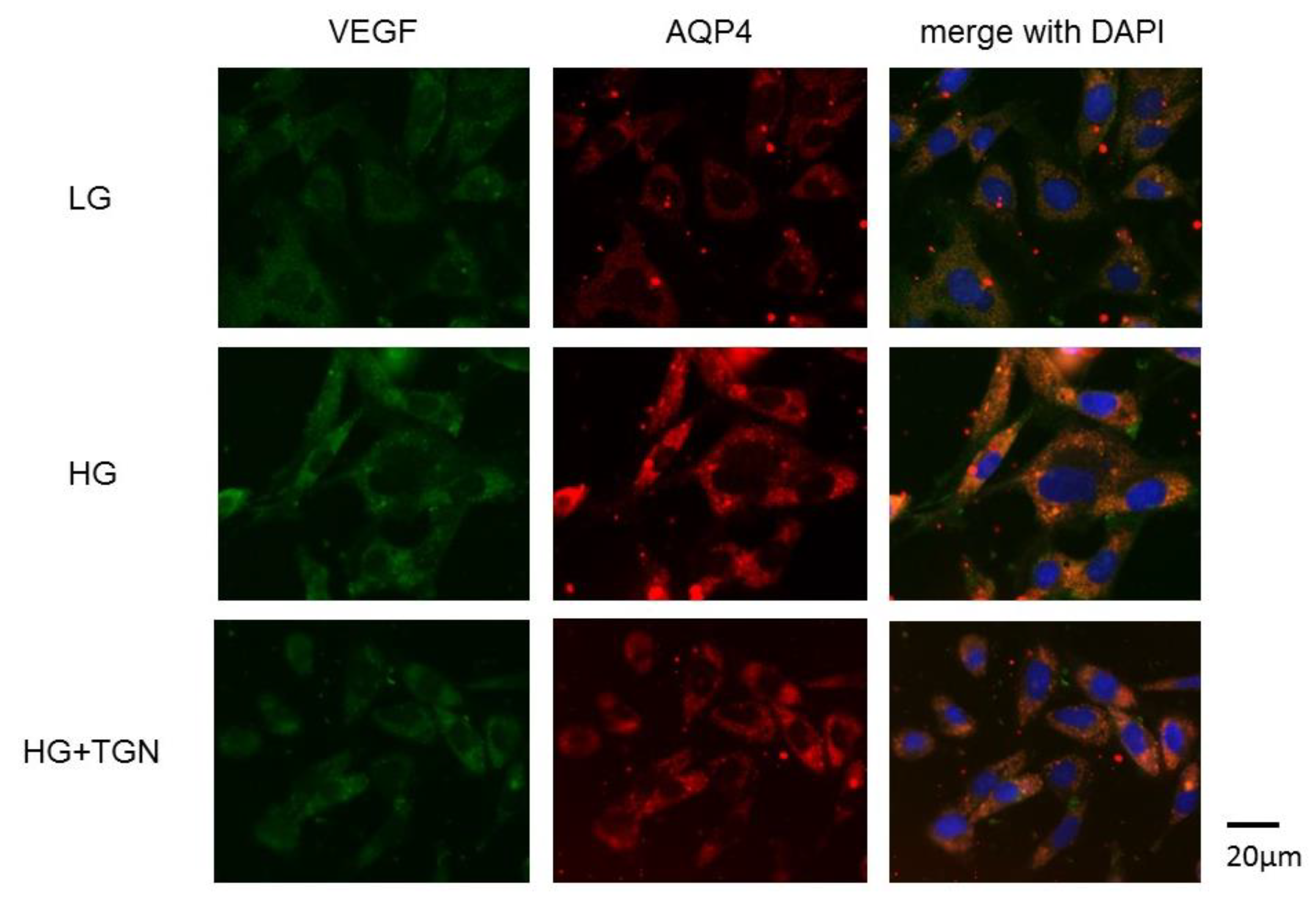
2.1. Immunohistochemistry of Retinal Slices
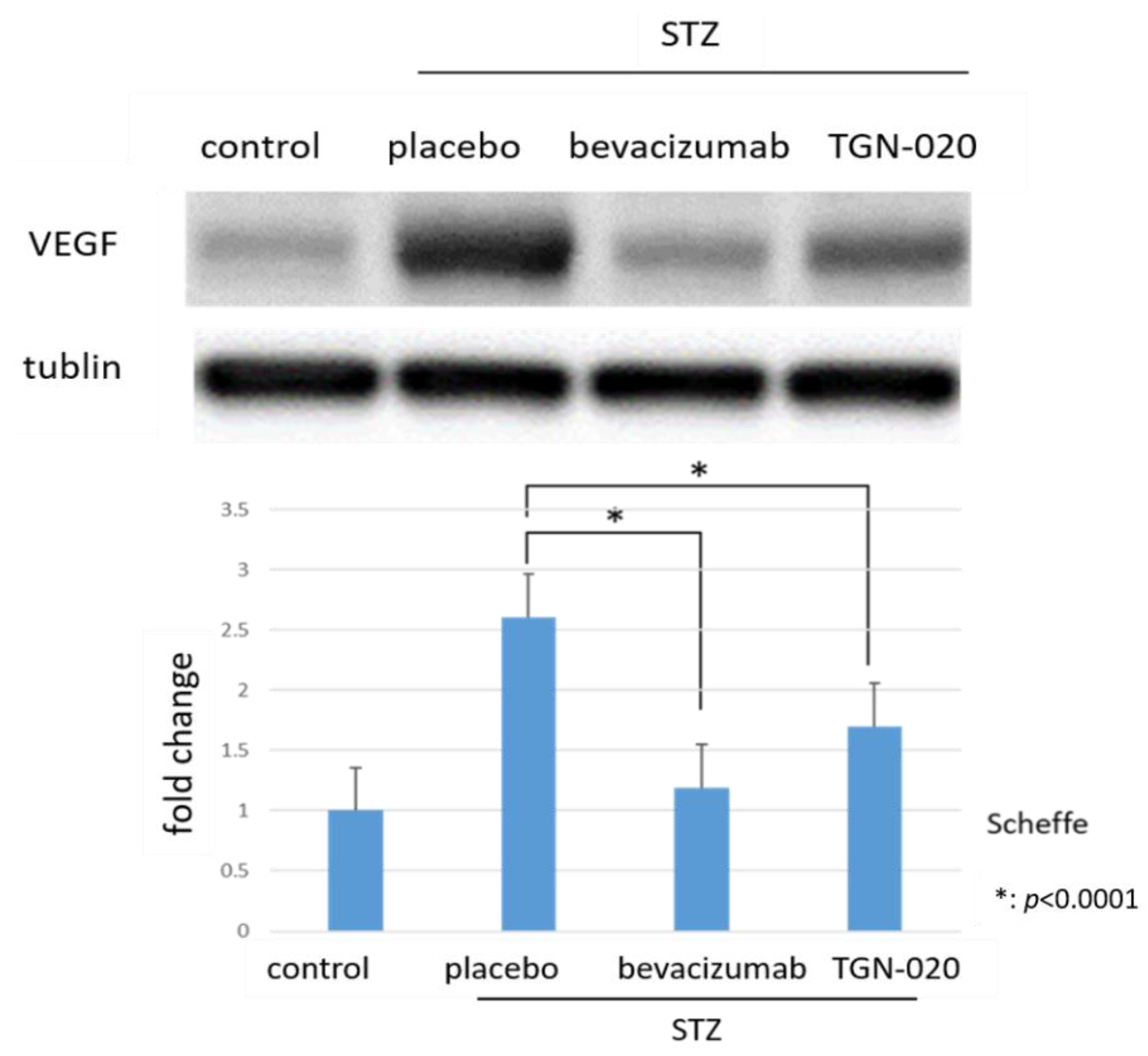
2.2. Protein Levels of VEGF in Retinas by Western Blot
2.3. Measurement of Retinal Thickness
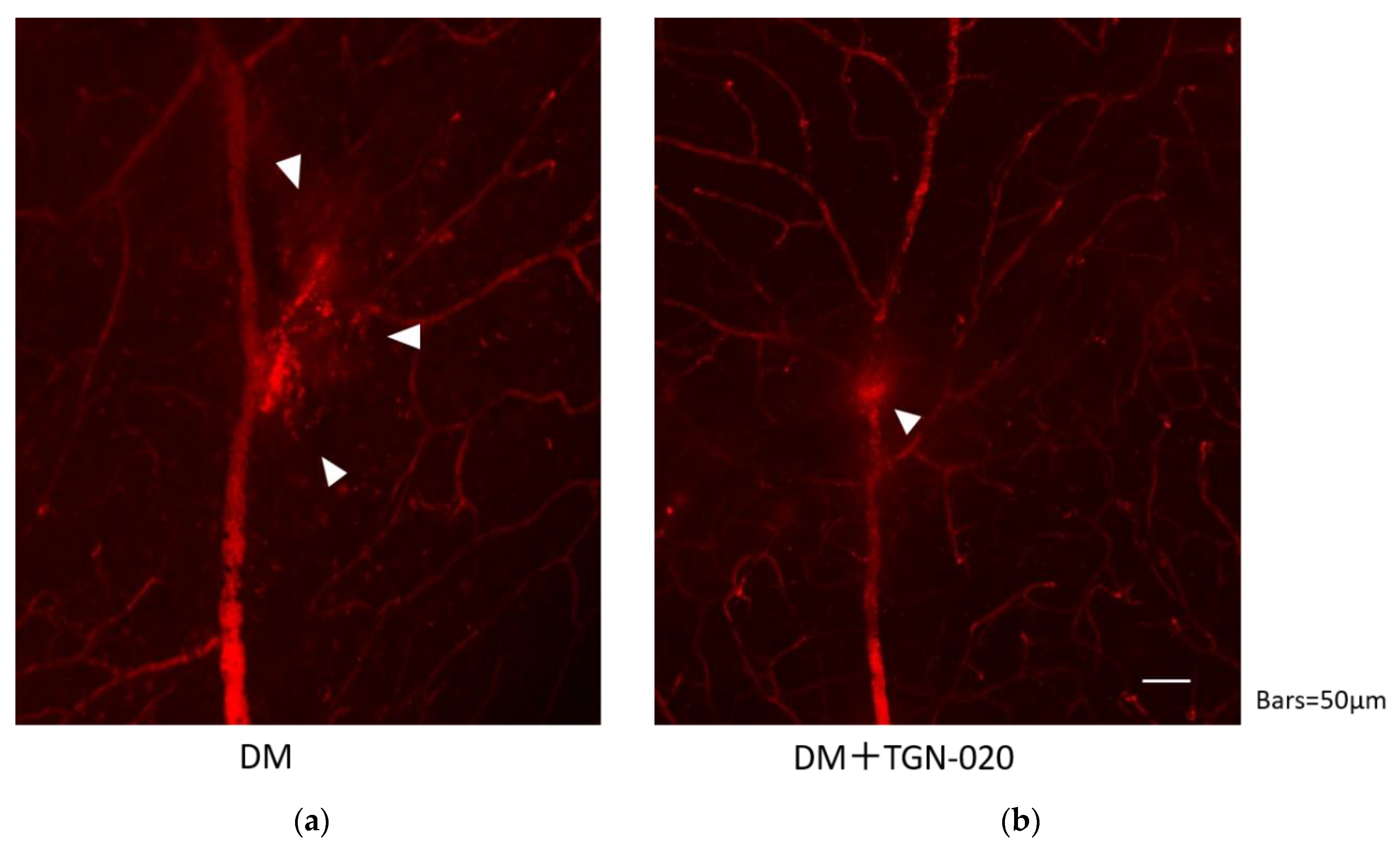
2.4. Images of Evans Blue Injected Flat-Mounted Retinas by Epifluorescence Microscope
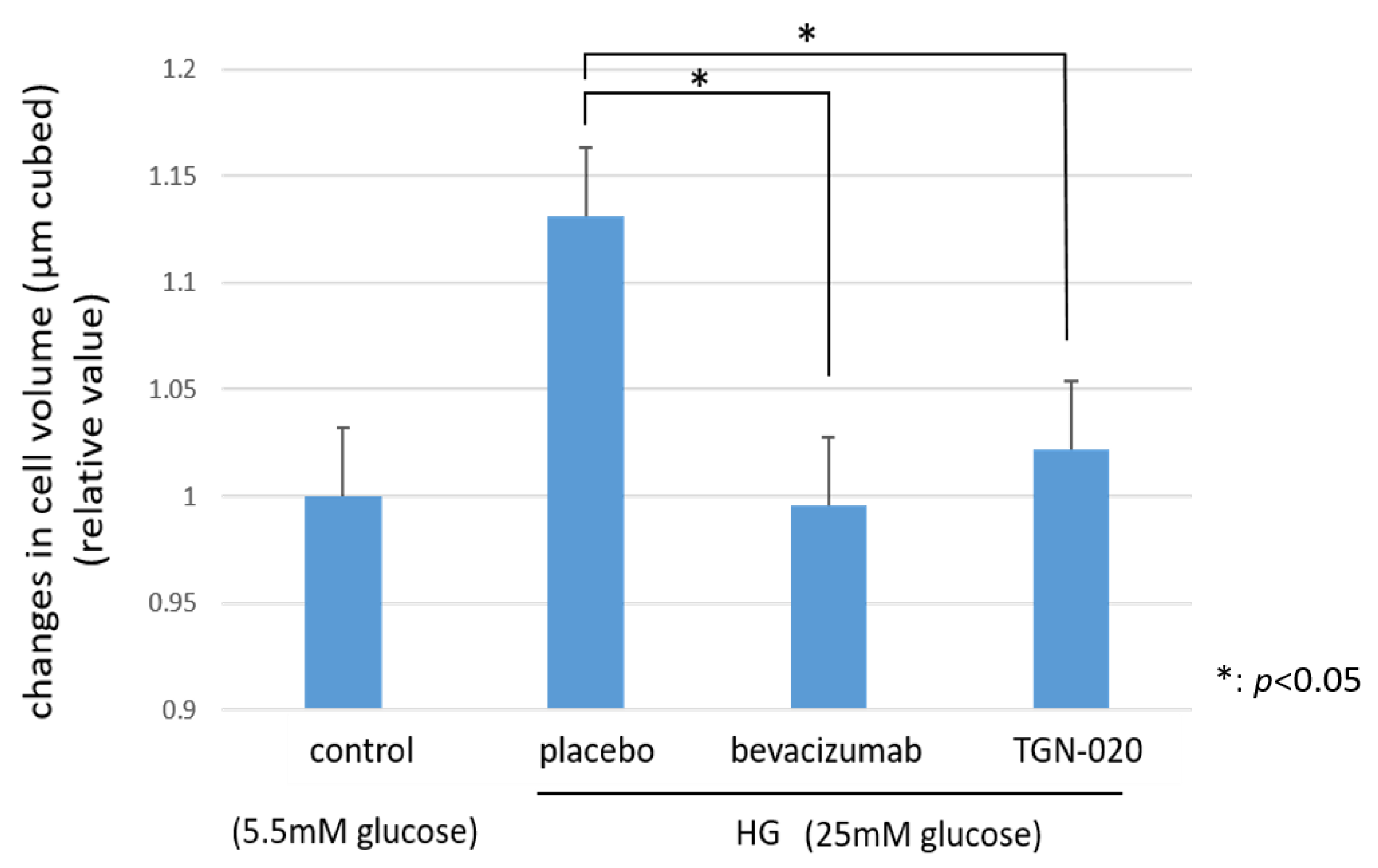
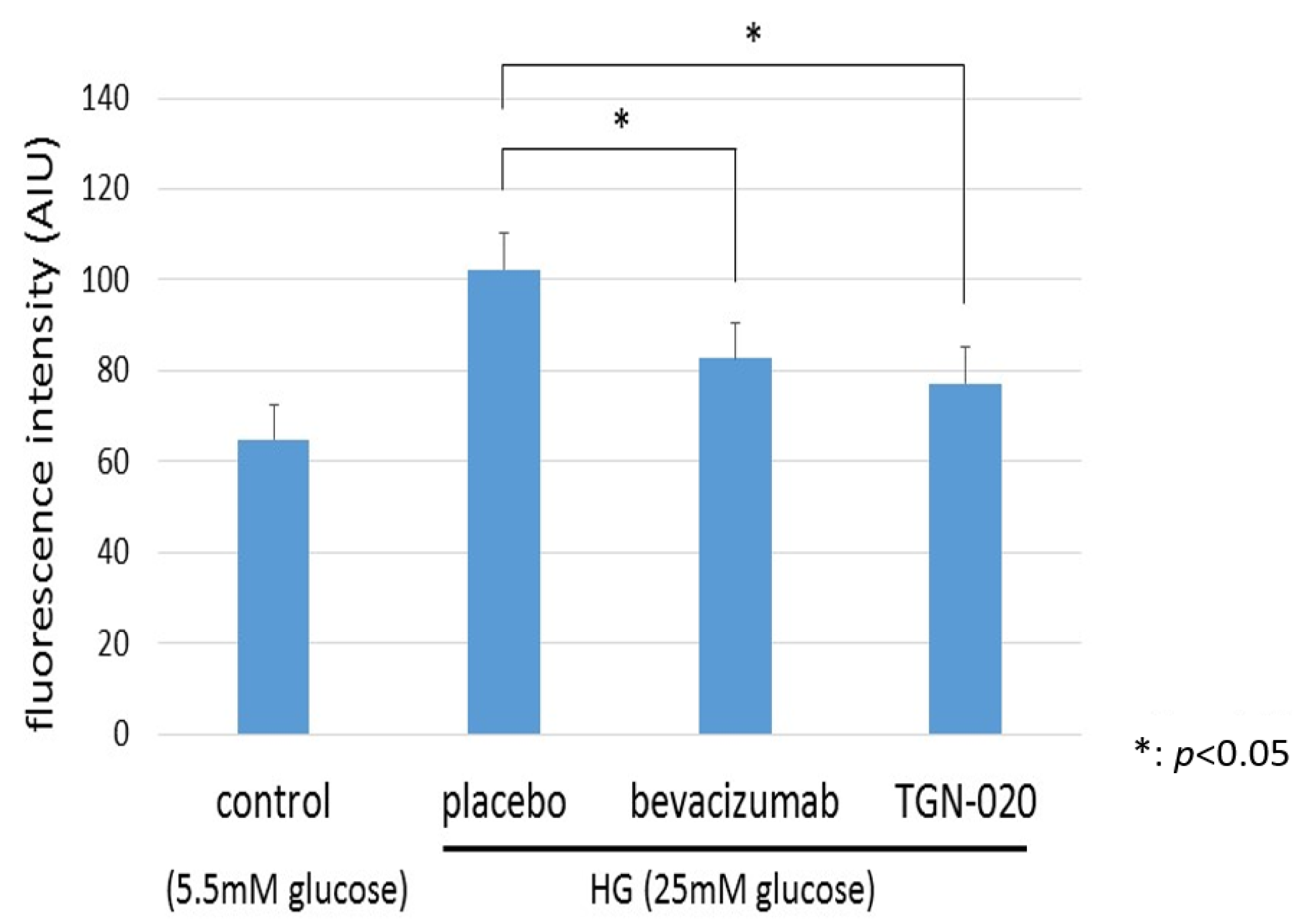
2.5. Cellular Volume and Intracellular Levels of ROS by Flow Cytometry
2.6. Immnostaining of Cultured Müller Cells (TR-MUL5)
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Induction of Diabetes
4.4. Intravitreal Injection
4.5. Immunohistochemistry of Retinal Slices
4.6. Protein Levels of VEGF in Retinas by Western Blot
4.7. Measurement of Retinal Thicknesses
4.8. Evans Blue Assay
4.9. Cell Culture
4.10. Measurement of Cell Volume and Intracellular Levels of ROS Production by Flow Cytometry
4.11. Immunostaining of Cultured Müller Cells
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AQP4 | aquaporin 4 |
| VEGF | vascular endothelial growth factor |
| ROS | reactive oxygen species |
| GFAP | anti-glial fibrillary acidic protein |
| STZ | streptozotocin |
| DME | diabetic macular edema |
| ILM | internal limiting membrane |
| IPL | inner plexiform layer |
| INL | inner nuclear layer |
| OPL | outer plexiform layer |
| ONL | outer nuclear layer |
References
- Whitmire, W.; Al-Gayyar, M.M.; Abdelsaid, M.; Yousufzai, B.K.; El-Remessy, A.B. Alteration of growth factors and neuronal death in diabetic retinopathy: What we have learned so far. Mol. Vis. 2011, 17, 300–308. [Google Scholar] [PubMed]
- Busch, S.; Kannt, A.; Kolibabka, M.; Schlotterer, A.; Wang, Q.; Lin, J.; Feng, Y.; Hoffmann, S.; Gretz, N.; Hammes, H.P. Systemic treatment with erythropoietin protects the neurovascular unit in a rat model of retinal neurodegeneration. PLoS ONE 2014, 9, e102013. [Google Scholar] [CrossRef] [PubMed]
- Bandello, F.; Tejerina, A.N.; Vujosevic, S.; Varano, M.; Egan, C.; Sivaprasad, S.; Menon, G.; Massin, P.; Verbraak, F.D.; Lund-Andersen, H.; et al. Retinal layer location of increased retinal thickness in eyes with subclinical and clinical macular edema in diabetes type 2. Ophthalmic. Res. 2015, 54, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Romero-Aroca, P.; De La Riva-Fernandez, S.; Valls-Mateu, A.; Sagarra-Alamo, R.; Moreno-Ribas, A.; Soler, N. Changes observed in diabetic retinopathy: Eight-year follow-up of a Spanish population. Br. J. Ophthalmol. 2016, 100, 1366–1371. [Google Scholar] [CrossRef] [PubMed]
- Lally, D.R.; Shah, C.P.; Heier, J.S. Vascular endothelial growth factor and diabetic macular edema. Surv. Ophthalmol. 2016, 61, 759–768. [Google Scholar] [CrossRef]
- Aiello, L.P.; Beck, R.W.; Bressler, N.M.; Browning, D.J.; Chalam, K.V.; Davis, M.; Ferris, F.L., 3rd; Glassman, A.R.; Maturi, R.K.; Stockdale, C.R.; et al. Rationale for the diabetic retinopathy clinical research network treatment protocol for center-involved diabetic macular edema. Ophthalmology 2011, 118, e5–e14. [Google Scholar] [CrossRef]
- Wells, J.A.; Glassman, A.R.; Ayala, A.R.; Jampol, L.M.; Aiello, L.P.; Antoszyk, A.N.; Arnold-Bush, B.; Baker, W.C.; Bressler, N.M.; Browning, D.J.; et al. Aflibercept, bevacizumab, or ranibizumab for diabetic macular edema. N. Engl. J. Med. 2015, 372, 1193–1203. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Kowluru, A.; Veluthakal, R.; Mohammad, G.; Syed, I.; Santos, J.M.; Mishra, M. TIAM1-RAC1 signalling axis-mediated activation of NADPH oxidase-2 initiates mitochondrial damage in the development of diabetic retinopathy. Diabetologia 2014, 57, 1047–1056. [Google Scholar] [CrossRef]
- Kida, T.; Oku, H.; Horie, T.; Matsuo, J.; Kobayashi, T.; Fukumoto, M.; Ikeda, T. NADPH oxidase-mediated ROS production determines insulin’s action on the retinal microvasculature. Invest. Ophthalmol. Vis. Sci. 2015, 56, 6754–6761. [Google Scholar] [CrossRef]
- Ola, M.S.; Nawaz, M.I.; Khan, H.A.; Alhomida, A.S. Neurodegeneration and neuroprotection in diabetic retinopathy. Int. J. Mol. Sci. 2013, 14, 2559–2572. [Google Scholar] [CrossRef]
- Alizadeh, E.; Mammadzada, P.; André, H. The different facades of retinal and choroidal endothelial cells in response to hypoxia. Int. J. Mol. Sci. 2018, 19, 3846. [Google Scholar] [CrossRef] [PubMed]
- Romero-Aroca, P.; Baget-Bernaldiz, M.; Pareja-Rios, A.; Lopez-Galvez, M.; Navarro-Gil, R.; Verges, R. Diabetic macular edema pathophysiology: Vasogenic versus inflammatory. J. Diabetes Res. 2016, 2156273. [Google Scholar] [CrossRef] [PubMed]
- Nagelhus, E.A.; Horio, Y.; Inanobe, A.; Fujita, A.; Haug, F.A.; Nielsen, S.; Kurachi, Y.; Ottersen, O.P. Immunogold evidence suggests that coupling of K+ siphoning and water transport in rat retinal Müller cells is mediated by a coenrichment of Kir4.1 and AQP4 in specific membrane domains. Glia 1999, 26, 47–54. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Manley, G.T.; Krishna, S.; Verkman, A.S. Aquaporin-4 facilitates reabsorption of excess fluid in vasogenic brain edema. FASEB J. 2004, 18, 1291–1293. [Google Scholar] [CrossRef]
- Kaur, C.; Sivakumar, V.; Zhang, Y.; Ling, E.A. Hypoxia-induced astrocytic reaction and increased vascular permeability in the rat cerebellum. Glia 2006, 54, 826–839. [Google Scholar] [CrossRef]
- Watkins, W.M.; McCollum, G.W.; Savage, S.R.; Capozzi, M.E.; Penn, J.S.; Morrison, D.G. Hypoxia-induced expression of VEGF splice variants and protein in four retinal cell types. Exp. Eye Res. 2013, 116, 240–246. [Google Scholar] [CrossRef]
- Schey, K.L.; Wang, Z.L.; Wenke, J.; Qi, Y. Aquaporins in the eye: Expression, function, and roles in ocular disease. Biochim. Biophys. Acta 2014, 1840, 1513–1523. [Google Scholar] [CrossRef]
- Bringmann, A.; Uckermann, O.; Pannicke, T.; Iandiev, I.; Reichenbach, A.; Wiedemann, P. Neuronal versus glial cell swelling in the ischaemic retina. Acta Ophthalmol. Scand. 2005, 83, 528–538. [Google Scholar] [CrossRef]
- Bi, C.; Tham, D.K.L.; Perronnet, C.; Joshi, B.; Nabi, I.R.; Moukhles, H. The oxidative stress-induced increase in the membrane expression of the water-permeable channel aquaporin-4 in astrocytes is regulated by caveolin-1 phosphorylation. Front. Cell Neurosci. 2017, 11, 412. [Google Scholar] [CrossRef]
- Da, T.; Verkman, A.S. Aquaporin-4 gene disruption in mice protects against impaired retinal function and cell death after ischemia. Invest. Ophthalmol. Vis. Sci. 2004, 45, 4477–4483. [Google Scholar] [CrossRef]
- Cui, B.; Sun, J.H.; Xiang, F.F.; Liu, L.; Li, W.J. Aquaporin 4 knockdown exacerbates streptozotocin-induced diabetic retinopathy through aggravating inflammatory response. Exp. Eye Res. 2012, 98, 37–43. [Google Scholar] [CrossRef]
- Kida, T.; Oku, H.; Horie, T.; Fukumoto, M.; Okuda, Y.; Morishita, S.; Ikeda, T. Implication of VEGF and aquaporin 4 mediating Müller cell swelling to diabetic retinal edema. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 1149–1157. [Google Scholar] [CrossRef]
- Huber, V.J.; Tsujita, M.; Nakada, T. Identification of aquaporin 4 inhibitors using in vitro and in silico methods. Bioorg. Med. Chem. 2009, 17, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Huber, V.J.; Tsujita, M.; Nakada, T. Pretreatment with a novel aquaporin 4 inhibitor, TGN-020, significantly reduces ischemic cerebral edema. Neurol. Sci. 2011, 32, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Wurm, A.; Pannicke, T.; Iandiev, I.; Wiedemann, P.; Bringmann, A. Müller cells as players in retinal degeneration and edema. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Fine, B.S.; Brucker, A.J. Macular edema and cystoid macular edema. Am. J. Ophthalmol. 1981, 92, 466–481. [Google Scholar] [CrossRef]
- Stokum, J.A.; Kurland, D.B.; Gerzanich, V.; Simard, J.M. Mechanisms of astrocyte-mediated cerebral edema. Neurochem Res. 2015, 40, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, A.; Reichenbach, A.; Wiedemann, P. Pathomechanisms of cystoid macular edema. Ophthalmic. Res. 2004, 36, 241–249. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Rose, C.F. Pathogenesis of hepatic encephalopathy in chronic liver disease. J. Clin. Exp. Hepatol. 2018, 8, 262–271. [Google Scholar] [CrossRef]
- Alrashdi, S.F.; Deliyanti, D.; Talia, D.M.; Wilkinson-Berka, J.L. Endothelin-2 injures the blood-retinal barrier and macroglial Müller cells: Interactions with angiotensin II, aldosterone, and NADPH oxidase. Am. J. Pathol. 2018, 188, 805–817. [Google Scholar] [CrossRef]
- Toyoda, F.; Tanaka, Y.; Shimmura, M.; Kinoshita, N.; Takano, H.; Kakehashi, A. Diabetic retinal and choroidal edema in SDT rats. J. Diabetes Res. 2016, 2016, 2345141. [Google Scholar] [CrossRef] [PubMed]
- Tomi, M.; Funaki, T.; Abukawa, H.; Katayama, K.; Kondo, T.; Ohtsuki, S.; Ueda, M.; Obinata, M.; Terasaki, T.; Hosoya, K. Expression and regulation of L-cystine transporter, system xc-, in the newly developed rat retinal Müller cell line (TR-MUL). Glia 2003, 43, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Obinata, M. The immortalized cell lines with differentiation potentials: Their establishment and possible application. Cancer Sci. 2007, 98, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Bindokas, V.P.; Jordán, J.; Lee, C.C.; Miller, R.J. Superoxide production in rat hippocampal neurons: Selective imaging with hydroethidine. J. Neurosci. 1996, 16, 1324–1336. [Google Scholar] [CrossRef]
- Zhao, H.; Joseph, J.; Fales, H.M.; Sokoloski, E.A.; Levine, R.L.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc. Natl. Acad. Sci. USA 2005, 102, 5727–5732. [Google Scholar] [CrossRef]
- Hoffman, R.A.; Johnson, T.S.; Britt, W.B. Flow cytometric electronic direct current volume and radiofrequency impedance measurements of single cells and particles. Cytometry 1981, 1, 377–384. [Google Scholar] [CrossRef]
- Potapova, T.A.; Seidel, C.W.; Box, A.C.; Rancati, G.; Li, R. Transcriptome analysis of tetraploid cells identifies cyclin D2 as a facilitator of adaptation to genome doubling in the presence of p53. Mol. Biol. Cell. 2016, 27, 3065–3084. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thickness (μm) | Control | DM | DM + TGN-020 |
|---|---|---|---|
| Total retina | 208.1 ± 19.6 | 253.7 ± 26.2 ** | 209.1 ± 33.1 |
| Between ILM and INL | 75. 7 ± 13.5 | 81.2 ± 9.8 | 69.0 ± 17.8 |
| INL | 31.3 ± 3.1 | 40.1 ± 5.0 ** | 33.2 ± 4.4 |
| OPL | 16.3 ± 6.7 | 18.1 ± 9.6 | 10.2 ± 7.1 |
| ONL | 57.7 ± 4.9 | 70.4 ± 6.8 ** | 58.0 ± 5.3 |
| Photoreceptor layer | 37.3 ± 5.0 | 44.3 ± 5.4 ** | 38.7 ± 6.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oosuka, S.; Kida, T.; Oku, H.; Horie, T.; Morishita, S.; Fukumoto, M.; Sato, T.; Ikeda, T. Effects of an Aquaporin 4 Inhibitor, TGN-020, on Murine Diabetic Retina. Int. J. Mol. Sci. 2020, 21, 2324. https://doi.org/10.3390/ijms21072324
Oosuka S, Kida T, Oku H, Horie T, Morishita S, Fukumoto M, Sato T, Ikeda T. Effects of an Aquaporin 4 Inhibitor, TGN-020, on Murine Diabetic Retina. International Journal of Molecular Sciences. 2020; 21(7):2324. https://doi.org/10.3390/ijms21072324
Chicago/Turabian StyleOosuka, Shou, Teruyo Kida, Hidehiro Oku, Taeko Horie, Seita Morishita, Masanori Fukumoto, Takaki Sato, and Tsunehiko Ikeda. 2020. "Effects of an Aquaporin 4 Inhibitor, TGN-020, on Murine Diabetic Retina" International Journal of Molecular Sciences 21, no. 7: 2324. https://doi.org/10.3390/ijms21072324
APA StyleOosuka, S., Kida, T., Oku, H., Horie, T., Morishita, S., Fukumoto, M., Sato, T., & Ikeda, T. (2020). Effects of an Aquaporin 4 Inhibitor, TGN-020, on Murine Diabetic Retina. International Journal of Molecular Sciences, 21(7), 2324. https://doi.org/10.3390/ijms21072324