Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS



Abstract

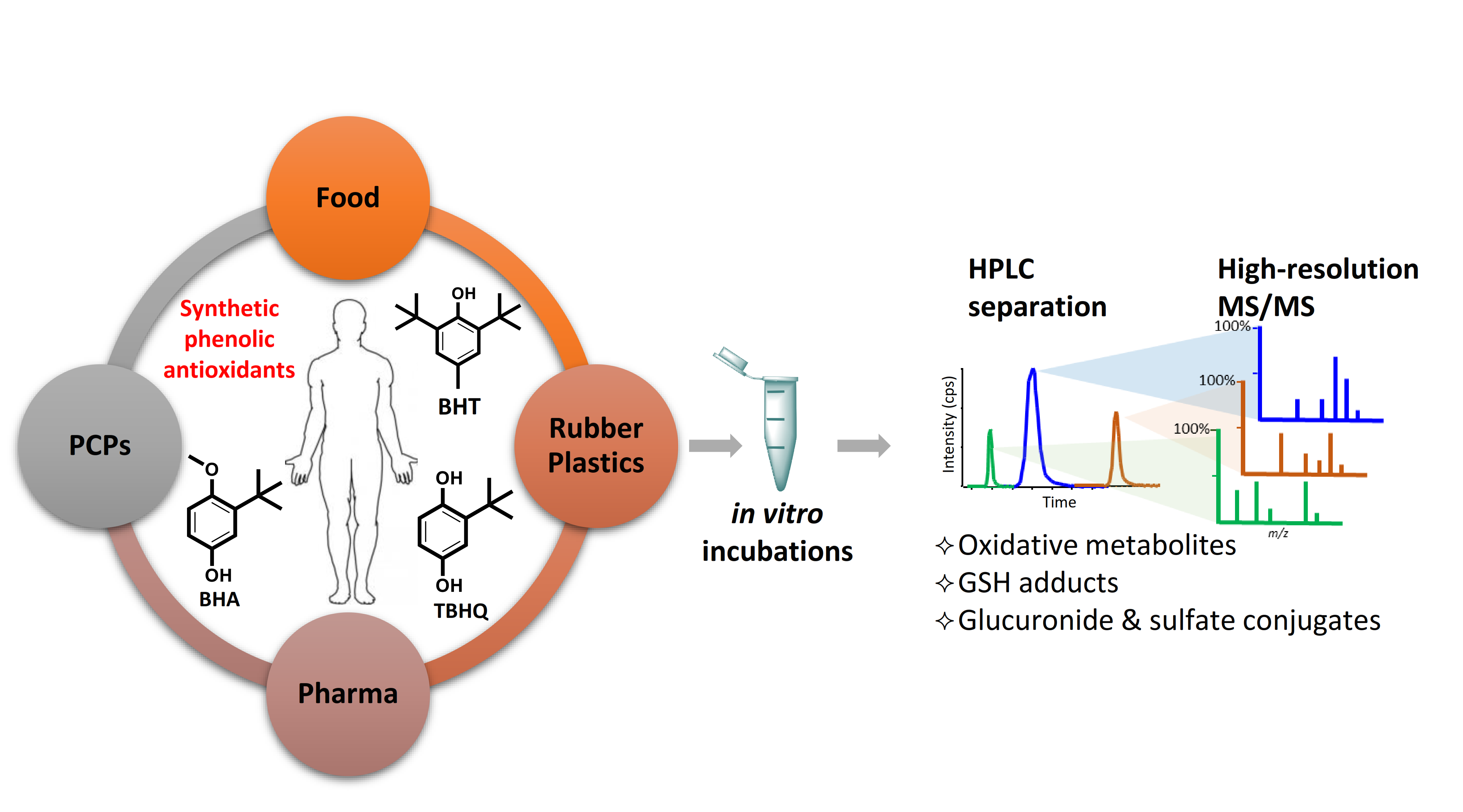
1. Introduction
2. Results and Discussion
2.1. Metabolism of BHT
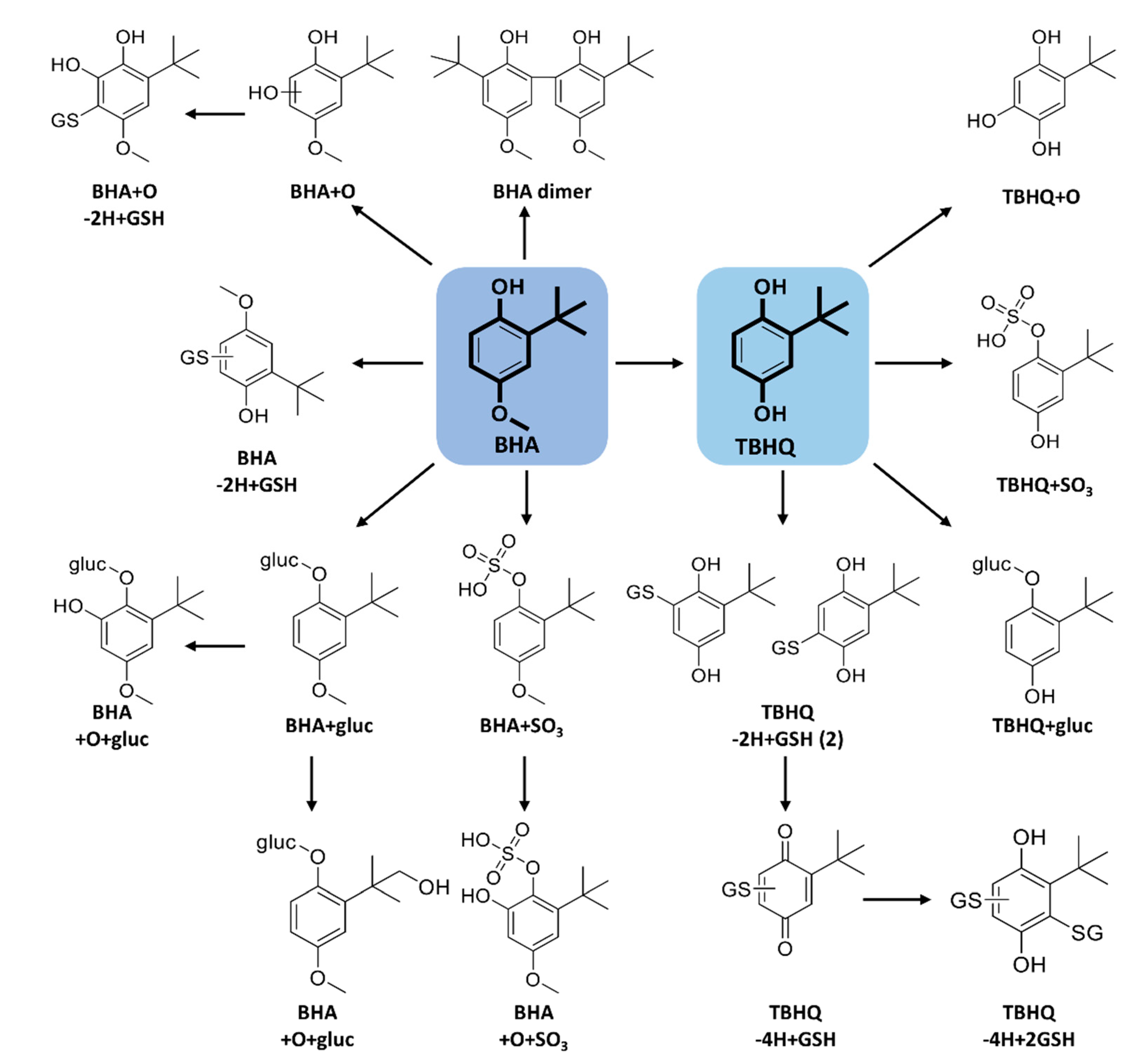
2.2. Metabolism of BHA and TBHQ
2.3. Metabolism of DBP and BHT Acid
3. Methods
3.1. Chemicals
3.2. In Vitro Incubations
3.2.1. Oxidative Metabolites and GSH Adducts
3.2.2. Phase II Metabolism—Glucuronidation and Sulfation
3.3. LC-HRMS/MS Analysis and Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nieva-Echevarría, B.; Manzanos, M.J.; Goicoechea, E.; Guillén, M.D. 2, 6-Di-tert-butyl-hydroxytoluene and its metabolites in foods. Compr. Rev. Food Sci. 2015, 14, 67–80. [Google Scholar] [CrossRef]
- Tortosa, V.; Pietropaolo, V.; Brandi, V.; Macari, G.; Pasquadibisceglie, A.; Polticelli, F. Computational Methods for the Identification of Molecular Targets of Toxic Food Additives. Butylated Hydroxytoluene as a Case Study. Molecules 2020, 25, 2229. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Mabury, S.A. Synthetic Phenolic Antioxidants in Personal Care Products in Toronto, Canada: Occurrence, Human Exposure, and Discharge via Greywater. Environ. Sci. Technol. 2019, 53, 13440–13448. [Google Scholar] [CrossRef] [PubMed]
- Saad, B.; Sing, Y.Y.; Nawi, M.A.; Hashim, N.; Ali, A.S.M.; Saleh, M.I.; Sulaiman, S.F.; Talib, K.M.; Ahmad, K. Determination of synthetic phenolic antioxidants in food items using reversed-phase HPLC. Food Chem. 2007, 105, 389–394. [Google Scholar] [CrossRef]
- Liu, R.; Mabury, S.A. Synthetic phenolic antioxidants and transformation products in human sera from United States donors. Environ. Sci. Technol. Lett. 2018, 5, 419–423. [Google Scholar] [CrossRef]
- Lanigan, R.S.; Yamarik, T.A. Final report on the safety assessment of BHT(1). Int. J. Toxicol. 2002, 21 (Suppl. 2), 19–94. [Google Scholar]
- Ahmad, H.; Sharma, R.; Mansour, A.; Awasthi, Y.C. t-butylated hydroxytoluene enhances intracellular levels of glutathione and related enzymes of rat lens in vitro organ culture. Exp. Eye Res. 1992, 54, 41–48. [Google Scholar] [CrossRef]
- Botterweck, A.A.; Verhagen, H.; Goldbohm, R.A.; Kleinjans, J.; van den Brandt, P.A. Intake of butylated hydroxyanisole and butylated hydroxytoluene and stomach cancer risk: Results from analyses in the Netherlands Cohort Study. Food Chem. Toxicol. 2000, 38, 599–605. [Google Scholar] [CrossRef]
- Nagai, F.; Ushiyama, K.; Kano, I. DNA cleavage by metabolites of butylated hydroxytoluene. Arch. Toxicol. 1993, 67, 552–557. [Google Scholar] [CrossRef]
- Kupfer, R.; Dwyer-Nield, L.D.; Malkinson, A.M.; Thompson, J.A. Lung toxicity and tumor promotion by hydroxylated derivatives of 2,6-di-tert-butyl-4-methylphenol (BHT) and 2-tert-butyl-4-methyl-6-iso-propylphenol: Correlation with quinone methide reactivity. Chem. Res. Toxicol. 2002, 15, 1106–1112. [Google Scholar] [CrossRef]
- Sun, Z.; Yang, X.; Liu, Q.S.; Li, C.; Zhou, Q.; Fiedler, H.; Liao, C.; Zhang, J.; Jiang, G. Butylated hydroxyanisole isomers induce distinct adipogenesis in 3T3-L1 cells. J. Hazard Mater. 2019, 379, 120794. [Google Scholar] [CrossRef] [PubMed]
- Goodman, D.L.; McDonnell, J.T.; Nelson, H.S.; Vaughan, T.R.; Weber, R.W. Chronic urticaria exacerbated by the antioxidant food preservatives, butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). J. Allergy Clin. Immunol. 1990, 86, 570–575. [Google Scholar] [CrossRef]
- World Health Organization (WHO); International Agency for Research on Cancer (IARC). Some naturally occurring and synthetic food components, furocoumarins and ultraviolet radiation, Summary of Data Reported and Evaluation. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1986; Volume 40, pp. 123–206. [Google Scholar]
- Ansar, S.; Iqbal, M. Antioxidant and nephroprotective potential of butylated hydroxyanisole against ferric nitrilotriacetate-induced oxidative stress and early tumor events. Hum. Exp. Toxicol. 2016, 35, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Mense, S.M.; Remotti, F.; Liu, X.; Bhat, H.K. Antioxidant butylated hydroxyanisole inhibits estrogen-induced breast carcinogenesis in female ACI rats. J. Biochem. Mol. Toxicol. 2009, 23, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Fukushima, S.; Tsuda, H. Carcinogenicity and modification of the carcinogenic response by BHA, BHT, and other antioxidants. Crit. Rev. Toxicol. 1985, 15, 109–150. [Google Scholar] [CrossRef] [PubMed]
- Pop, A.; Kiss, B.; Loghin, F. Endocrine disrupting effects of butylated hydroxyanisole (BHA-E320). Clujul Med. 2013, 86, 16–20. [Google Scholar]
- Choe, S.; Yang, K. Toxicological studies of antioxidants, butylated hydroxytoluene (BHT) and butylated hydroxyanisole (BHA). Korean J. Food Nutr. 1982, 14, 283–288. [Google Scholar]
- Gharavi, N.; Haggarty, S.; El-Kadi, A.O. Chemoprotective and carcinogenic effects of tert-butylhydroquinone and its metabolites. Curr. Drug Metab. 2007, 8, 1–7. [Google Scholar] [CrossRef]
- Almeida, E.S.; Portela, F.M.; Sousa, R.M.; Daniel, D.; Terrones, M.G.; Richter, E.M.; Muñoz, R.A. Behaviour of the antioxidant tert-butylhydroquinone on the storage stability and corrosive character of biodiesel. Fuel 2011, 90, 3480–3484. [Google Scholar] [CrossRef]
- Van Esch, G.J. Toxicology of tert-butylhydroquinone (TBHQ). Food Chem. Toxicol. 1986, 24, 1063–1065. [Google Scholar] [CrossRef]
- Li, C.; Cui, X.; Chen, Y.; Liao, C.; Ma, L.Q. Synthetic phenolic antioxidants and their major metabolites in human fingernail. Environ. Res. 2019, 169, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, C.; Li, Y.; Cui, X.; Ma, L.Q. Determination of 2,6-di-tert-butyl-hydroxytoluene and its transformation products in indoor dust and sediment by gas chromatography-mass spectrometry coupled with precolumn derivatization. Sci. Total Environ. 2018, 619–620, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Kannan, K. Quantitative identification of and exposure to synthetic phenolic antioxidants, including butylated hydroxytoluene, in urine. Environ. Int. 2019, 128, 24–29. [Google Scholar] [CrossRef]
- E 320 Butylated Hydroxy-Anisole (BHA). The UK Food Guide. Available online: http://ukfoodguide.net/e321.htm (accessed on 25 October 2020).
- Health Canada. 2020. List of Permitted Preservatives. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-safety/food-additives/lists-permitted/11-preservatives.html#a4 (accessed on 30 August 2020).
- Suh, H.J.; Chung, M.S.; Cho, Y.H.; Kim, J.W.; Kim, D.H.; Han, K.W.; Kim, C.J. Estimated daily intakes of butylated hydroxyanisole (BHA), butylated hydroxytoluene (BHT) and tert-butyl hydroquinone (TBHQ) antioxidants in Korea. Food Addit. Contam. 2005, 22, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Fries, E.; Puttmann, W. Analysis of the antioxidant butylated hydroxytoluene (BHT) in water by means of solid phase extraction combined with GC/MS. Water Res. 2002, 36, 2319–2327. [Google Scholar] [CrossRef]
- Zhang, R.; Li, J.; Cui, X. Tissue distribution, excretion, and metabolism of 2, 6-di-tert-butyl-hydroxytoluene in mice. Sci. Total Environ. 2020, 739, 139862. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hu, C.; Zhong, H.; Chen, X.; Chen, R.; Yam, K.L. Effects of Ultraviolet (UV) on Degradation of Irgafos 168 and Migration of Its Degradation Products from Polypropylene Films. J. Agric. Food Chem. 2016, 64, 7866–7873. [Google Scholar] [CrossRef]
- Pont, M.M. Health-Based Reassessment of Administrative Occupational Exposure Limits: 2, 6-Di-tert-butyl-p-cresol (BHT/Butylated Hydroxytoluene); Gezondheidsraad: The Hague, The Netherlands, 2004.
- Thompson, J.A.; Malkinson, A.M.; Wand, M.D.; Mastovich, S.L.; Mead, E.W.; Schullek, K.M.; Laudenschlager, W.G. Oxidative metabolism of butylated hydroxytoluene by hepatic and pulmonary microsomes from rats and mice. Drug Metab. Dispos. 1987, 15, 833–840. [Google Scholar]
- Conning, D.M.; Phillips, J.C. Comparative metabolism of BHA, BHT and other phenolic antioxidants and its toxicological relevance. Food Chem. Toxicol. 1986, 24, 1145–1148. [Google Scholar] [CrossRef]
- Tajima, K.; Yamamoto, K.; Mizutani, T. Identification and determination of glutathione and glucuronide conjugates formed from butylated hydroxytoluene in rats. Chem. Pharm. Bull. 1983, 31, 3671–3677. [Google Scholar] [CrossRef]
- Madsen, K.G.; Olsen, J.; Skonberg, C.; Hansen, S.H.; Jurva, U. Development and evaluation of an electrochemical method for studying reactive phase-I metabolites: Correlation to in vitro drug metabolism. Chem. Res. Toxicol. 2007, 20, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, K.E.; Wattenberg, L.W. Metabolism of 3-tert-butyl-4-hydroxyanisole to 3-tert-butyl-4,5-dihydroxyanisole by rat liver microsomes. Cancer Res. 1985, 45, 1507–1510. [Google Scholar] [PubMed]
- Sgaragli, G.; Corte, L.D.; Puliti, R.; De Sarlo, F.; Francalanci, R.; Guarna, A.; Dolara, P.; Komarynsky, M. Oxidation of 2-t-butyl-4-methoxyphenol (BHA) by horseradish and mammalian peroxidase systems. Biochem. Pharmacol. 1980, 29, 763–769. [Google Scholar] [CrossRef]
- Peters, M.M.; Lau, S.S.; Dulik, D.; Murphy, D.; van Ommen, B.; van Bladeren, P.J.; Monks, T.J. Metabolism of tert-butylhydroquinone to S-substituted conjugates in the male Fischer 344 rat. Chem. Res. Toxicol. 1996, 9, 133–139. [Google Scholar] [CrossRef]
- El-Rashidy, R.; Niazi, S. A new metabolite of butylated hydroxyanisole in man. Biopharm. Drug Dispos. 1983, 4, 389–396. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Formula | m/z [M-H]− | ppm | RT (min) | Selected MS/MS Fragments * |
|---|---|---|---|---|---|
| BHT | C15H24O | 219.1760 | 2.7 | 14.3 | 203.1439 |
| BHT + O (1) | C15H24O2 | 235.1708 | 2.1 | 13.4 | 217.1598, 160.0895, 145.0657 |
| BHT + O (2) | C15H24O2 | 235.1705 | 0.8 | 14.1 | 217.1597, 205.1597 |
| BHT + 2O | C15H24O3 | 251.1660 | 2.9 | 11.9 | 233.1547, 221.1557, 203.1444, 160.0893, 146.0746 |
| BHT−aldehyde | C15H22O2 | 233.1552 | 2.2 | 14.0 | 217.1234 |
| BHT−acid | C15H22O3 | 249.1501 | 2.2 | 9.8 | 205.1592 |
| BHT−aldehyde + O | C15H22O3 | 249.1503 | 3.0 | 13.5 | 231.1400, 219.1399 |
| DBP | C14H22O | 205.1600 | 1.2 | 14.2 | 189.1289 |
| BHQ | C14H22O2 | 221.1549 | 0.9 | 8.3 | 164.0848, 149.0601 |
| BHT−2H + GSH | C25H39N3O7S | 524.2436 | 0.1 | 8.5 | 306.0772, 272.0892, 254.0788, 160.0078, 143.0463, 128.0353 |
| BHT + O-2H + GSH | C25H39N3O8S | 540.2381 | −0.7 | 7.6 | 306.0758, 288.0646, 272.0882, 254.0778, 160.0078, 143.0465, 128.0348 |
| DBP-2H + GSH | C24H37N3O7S | 510.2287 | 1.5 | 8.2 | 492.2179, 272.0886, 254.0793, 237.1316, 143.0462, 128.0353 |
| DBP + gluc | C20H30O7 | 381.1924 | 1.4 | 8.7 | 205.1595, 175.0247, 113.0240, 85.0291 |
| BHT + O + gluc | C21H32O8 | 411.2026 | 0.5 | 8.8 | 193.0360, 131.0341, 113.0244, 85.0289 |
| BHT−acid + gluc | C21H30O9 | 425.1820 | 0.9 | 8.3 | 249.1491, 193.0354, 175.0247, 131.0344, 113.0239, 72.9929 |
| BHQ + gluc | C20H30O8 | 397.1864 | −1.0 | 7.1 | 221.1537, 175.0135, 113.0242, 85.0290 |
| BHQ + SO3 | C14H22O5S | 301.1117 | 0.8 | 9.0 | 221.1539, 164.0844, 80.9645, 79.9576 |
| Metabolite | BHT-d3 Formula m/z (ppm) | BHT-d20 Formula m/z (ppm) | Comments |
|---|---|---|---|
| BHT + O (1) | C15D2H22O2 237.1834 (2.1) | C15D20H4O2 255.2967 (3.2) | O added on p-CH3 |
| BHT + O (2) | C15D3H21O2 238.1893 (0.7) | C15D19H4O2 254.2903 (2.6) | O added on t-butyl |
| BHT + 2O | C15D2H22O3 238.1893 (0.7) | C15D19H4O3 270.2855 (3.6) | O added on p-CH3 and t-butyl |
| BHT−aldehyde | C15H22O2 233.1553 (2.5) | C15D20H2O2 253.2811 (3.4) | Aldehyde in para position |
| BHT−acid | C15H22O3 249.1505 (3.5) | C15D20H2O3 269.2757 (2) | Carboxylic acid in para position |
| BHT−aldehyde + O | C15H22O3 249.1504 (3.1) | C15D19H2O3 268.2689 (0.1) | CHO on p-methyl, OH on t-butyl |
| DBP | C14H22O 205.1607 (4.4) | C14D20H2O 225.2861 (3.4) | DBP structure |
| BHQ | C14H22O2 221.1548 (0.4) | C14D20H2O2 241.2797 (−2.2) | O added to DBP in para position |
| BHT−2H + GSH | C25D2H37N3O7S 526.2560 (−0.2) | C25D20H19N3O7S 544.3691 (−0.1) | SG group added to p-CH3 |
| BHT + O−2H + GSH | C25D2H37N3O8S 542.2508 (−0.4) | C25D19H20N3O8S 559.3588 (1.8) | SG added to p-CH3 and O on t-butyl |
| DBP−2H + GSH | C24H37N3O7S 510.2274 (−1.1) | C24D20H17N3O7S 530.3541 (1.2) | SG added to ring in para position |
| DBP + gluc | C20H30O7 381.1924 (1.4) | C20D20H10O7 401.3184 (2.5) | Glucuronide on hydroxyl of DBP |
| BHT + O + gluc | C21D3H29O8 414.2222 (2.2) | C21D19H13O8 430.3218 (0.2) | Glucuronide on O added on t-butyl |
| BHT−acid + gluc | C21H30O9 425.1821 (0.9) | C21D20H10O9 445.3068 (−1.0) | Glucuronide on carboxylic acid |
| BHQ + gluc | C20H30O8 397.1877 (2.3) | C20D20H10O8 417.3120 (0.8) | Glucuronide added to para-OH |
| BHQ + SO3 | C14H22O5S 301.1116 (0.5) | C14D20H10O9 321.2369 (−0.4) | SO3 attached to BHQ |
| Metabolite | Formula | m/z [M-H]− | ppm | RT (min) | Selected MS/MS Fragments * |
|---|---|---|---|---|---|
| BHA | C11H16O2 | 179.1084 | 3.6 | 12.7 | 164.0838, 149.0604, 121.0656, 108.0213 |
| BHA + O (1) | C11H16O3 | 195.1027 | 2.3 | 11.8 | 180.0795, 165.0560, 137.0609 |
| BHA + O (2) | C11H16O3 | 195.1031 | 2.4 | 11.4 | 180.0784, 165.0552 |
| BHA dimer | C22H30O4 | 357.2076 | 1.5 | 14.4 | 342.1831, 327.1608 |
| BHA-2H + GSH | C21H31N3O8S | 484.1760 | 0.2 | 5.8 | 272.0898, 254.0791, 211.0810, 210.0884, 179.0456, 143.0461, 128.0345 |
| BHA + O-2H + GSH | C21H31N3O9S | 500.1709 | 0.3 | 5.0 | 306.0759, 272.0864, 254.0750, 210.0892, 179.0450, 160.0056, 143.0470, 128.0371 |
| TBHQ-2H + GSH (1) | C20H29N3O8S | 470.1602 | −0.1 | 5.1 | 272.0924, 254.0822, 210.0916, 197.0660, 179.0479, 146.0468, 143.0471, 128.0358 |
| TBHQ-2H + GSH (2) | C20H29N3O8S | 470.1675 | 2.3 | 5.4 | 272.0925, 254.0821, 210.0911, 197.0662, 185.0586, 179.0476, 166.0998, 146.0462, 143.0466, 128.0358 |
| TBHQ-4H + GSH | C20H27N3O8S | 468.1446 | 0.6 | 6.4 | 339.1025, 272.0930, 254.0811, 210.0896, 197.0657, 195.0503, 192.1049, 179.0465, 146.0442, 143.0466, 128.0353 |
| TBHQ-4H + 2GSH | C30H44N6O14S2 | 775.2288 | 0.6 | 1.7 | 646.1861, 502.1334, 468.1445, 306.0769, 272.0890, 254.0790, 229.0358, 195.0488, 143.0463 |
| BHA + gluc | C17H24O8 | 355.1401 | 0.9 | 5.6 | 337.1300, 179.1082, 175.0252, 164.0845, 117.0196, 113.0243, 85.0296, 75.0090, 59.0139 |
| BHA + O + gluc (1) | C17H24O9 | 371.1351 | 1 | 5.7 | 353.1259, 265.1083, 195.1035, 180.0800, 175.0254, 113.0245, 85.0293, 75.0080, 59.0135 |
| BHA + O + gluc (2) | C17H24O9 | 371.1357 | 2.6 | 6.6 | 195.1027, 180.0792, 175.0242, 113.0237, 85.0287 |
| TBHQ + gluc | C16H22O8 | 341.1240 | −0.4 | 3.4 | 175.0253, 165.0920, 113.0247, 85.0295, 59.0137 |
| BHA + SO3 (1) | C11H16O5S | 259.0647 | 4.7 | 8.3 | 244.0405, 179.1070, 164.0842, 149.0606, 70.9571 |
| BHA + SO3 (2) | C11H16O5S | 259.0645 | 0.4 | 7.2 | 239.0718, 179.1077, 164.0844, 149.0635, 79.9571 |
| BHA + O + SO3 | C11H16O6S | 275.0597 | 0.8 | 8.4 | 195.1032,180.0794, 165.0559, 79.9574 |
| TBHQ + SO3 | C10H14O5S | 245.0489 | 1.4 | 5.7 | 165.0923, 149.0607, 108.0213, 80.9651, 79.9572 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ousji, O.; Sleno, L. Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS. Int. J. Mol. Sci. 2020, 21, 9525. https://doi.org/10.3390/ijms21249525
Ousji O, Sleno L. Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS. International Journal of Molecular Sciences. 2020; 21(24):9525. https://doi.org/10.3390/ijms21249525
Chicago/Turabian StyleOusji, Ons, and Lekha Sleno. 2020. "Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS" International Journal of Molecular Sciences 21, no. 24: 9525. https://doi.org/10.3390/ijms21249525
APA StyleOusji, O., & Sleno, L. (2020). Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS. International Journal of Molecular Sciences, 21(24), 9525. https://doi.org/10.3390/ijms21249525