Growth by Insertion: The Family of Bacterial DDxP Proteins



Abstract
1. Introduction
2. Results
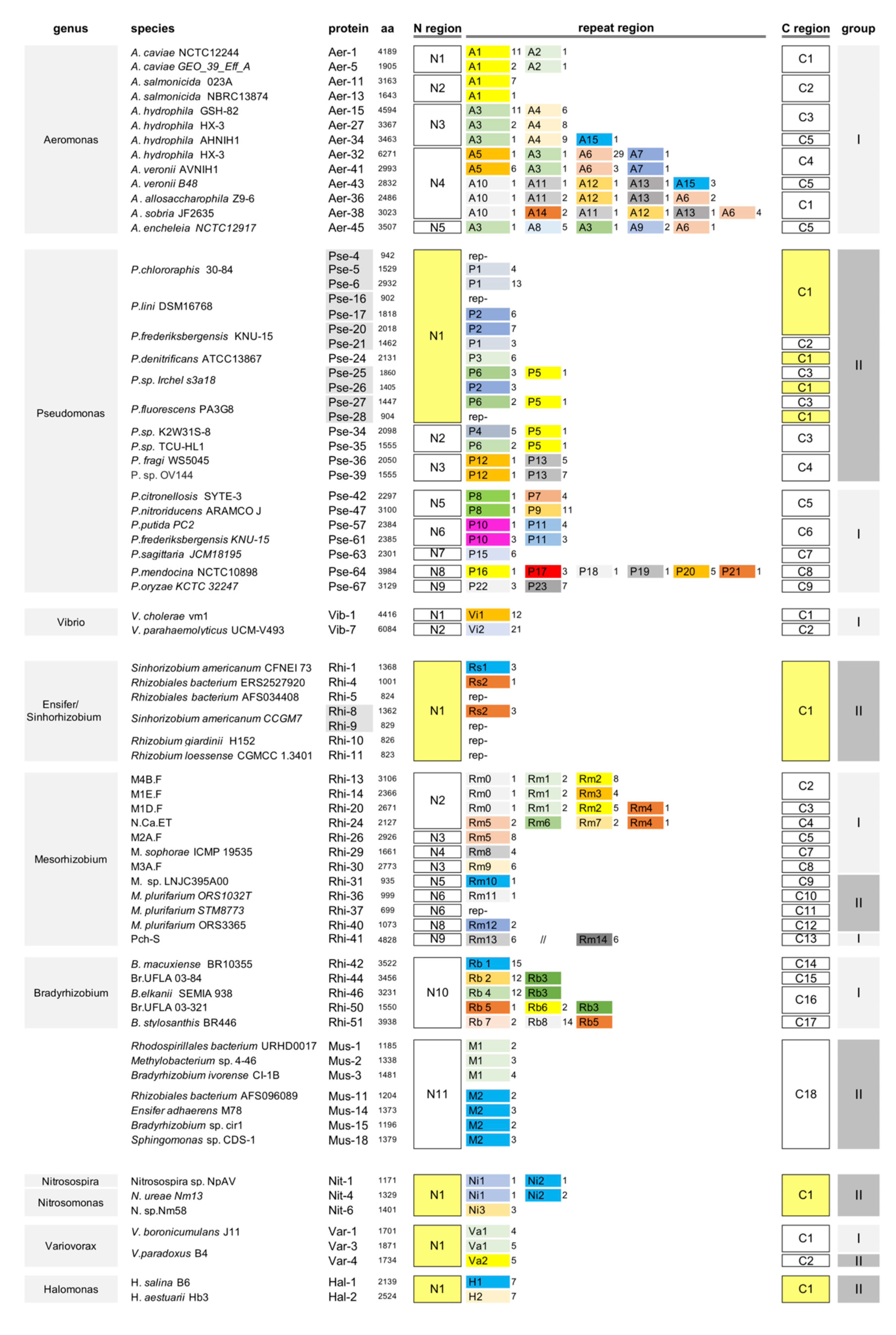
2.1. Multiple Repeat Types in Legionella Repeats-In-Toxins (RTX) Proteins
2.1.1. Search for DDxP Proteins
2.1.2. Gamma-Proteobacteria DDxP Proteins
2.1.3. Alpha-Proteobacteria DDxP Proteins
2.1.4. Additional DDxP Proteins
2.1.5. DDxP Proteins and T1SSs
2.1.6. DDxP Repeats
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | amino acids |
| RTX | repeats-in-toxins |
| T1SSs | type 1 secretion systems |
| TSDs | target site duplications |
| BIg-like | bacterial immunoglobulin-like |
References
- Green, E.R.; Mecsas, J. Bacterial Secretion Systems: An Overview. Microbiol. Spectr. 2016, 4, 213–239. [Google Scholar] [CrossRef] [PubMed]
- Guérin, J.; Bigot, S.; Schneider, R.; Buchanan, S.K.; Jacob-Dubuisson, F. Two-Partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-Host and Bacteria-Bacteria Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.A. Pore-forming cytolysins of Gram-negative bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O’Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef]
- Latasa, C.; Solano, C.; Penadés, J.R.; Lasa, I. Biofilm-associated proteins. C. R. Biol. 2006, 329, 849–857. [Google Scholar] [CrossRef]
- Yousef, F.; Espinosa-Urgel, M. In silico analysis of large microbial surface proteins. Res. Microbiol. 2007, 158, 545–550. [Google Scholar] [CrossRef]
- De Gregorio, E.; Del Franco, M.; Martinucci, M.; Roscetto, E.; Zarrilli, R.; Di Nocera, P.P. Biofilm-associated proteins: News from Acinetobacter. BMC Genom. 2015, 16, 933. [Google Scholar] [CrossRef]
- Guo, S.; Stevens, C.A.; Vance, T.D.R.; Olijve, L.L.C.; Graham, L.A.; Campbell, R.; Yazdi, S.R.; Escobedo, C.; Bar-Dolev, M.; Yashunsky, V.; et al. Structure of a 1.5-MDa adhesin that binds its Antarctic bacterium to diatoms and ice. Sci. Adv. 2017, 3, e1701440. [Google Scholar] [CrossRef]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii Biofilm-Associated Protein Plays a Role in Adherence to Human Epithelial Cells. Infect. Immun. 2011, 80, 228–233. [Google Scholar] [CrossRef]
- Barlag, B.; Hensel, M. The Giant Adhesin SiiE of Salmonella enterica. Molecules 2015, 20, 1134–1150. [Google Scholar] [CrossRef]
- Hayes, C.S.; Koskiniemi, S.; Ruhe, Z.C.; Poole, S.J.; Low, D.A. Mechanisms and Biological Roles of Contact-Dependent Growth Inhibition Systems. Cold Spring Harb. Perspect. Med. 2014, 4, a010025. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Zarrilli, R.; Di Nocera, P.P. Contact-dependent growth inhibition systems in Acinetobacter. Sci. Rep. 2019, 9, 154. [Google Scholar]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [PubMed]
- Satchell, K.J.F. Structure and Function of MARTX Toxins and Other Large Repetitive RTX Proteins. Annu. Rev. Microbiol. 2011, 65, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Vance, T.D.R.; Corey, A.; Stevens, C.A.; Voets, I.K.; Davies, P.L. RTX Adhesins are Key Bacterial Surface Megaproteins in the Formation of Biofilms. Trends Microbiol. 2019, 5, 470. [Google Scholar] [CrossRef] [PubMed]
- Bumba, L.; Masin, J.; Macek, P.; Wald, T.; Motlova, L.; Bibova, I.; Klimova, N.; Bednarova, L.; Veverka, V.; Kachala, M.; et al. Calcium-driven folding of RTX domain beta-rolls ratchets translocation of RTX proteins through type I secretion ducts. Mol. Cell. 2016, 62, 47–62. [Google Scholar] [CrossRef]
- Motlova, L.; Klimova, N.; Fiser, R.; Sebo, P.; Bumba, L. Continuous Assembly of β-Roll Structures Is Implicated in the Type I-Dependent Secretion of Large Repeat-in-Toxins (RTX) Proteins. J. Mol. Biol. 2020, 432, 5696–5710. [Google Scholar] [CrossRef]
- Peters, B.; Stein, J.; Klingl, S.; Sander, N.; Sandmann, A.; Taccardi, N.; Sticht, H.; Gerlach, R.; Muller, Y.A.; Hensel, M. Structural and functional dissection reveals distinct roles of Ca2+-binding sites in the giant adhesin SiiE of Salmonella enterica. PLoS Pathog. 2017, 13, e1006418. [Google Scholar] [CrossRef]
- Cirillo, S.L.; Bermudez, L.E.; El-Etr, S.H.; Duhamel, G.E.; Cirillo, J.D. Legionella pneumophila entry gene rtxA is involved in virulence. Infect. Immun. 2001, 69, 508–517. [Google Scholar] [CrossRef]
- D’Auria, G.; Jiménez, N.; Peris-Bondia, F.; Pelaz, C.; Latorre, A.; Moya, A. Virulence factor rtx in Legionella pneumophila, evidence suggesting it is a modular multifunctional protein. BMC Genom. 2008, 9, 14. [Google Scholar] [CrossRef]
- Fernández-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W., II; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative Genomics of Plant-Associated Pseudomonas spp.: Insights into Diversity and Inheritance of Traits Involved in Multitrophic Interactions. PLoS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef] [PubMed]
- Greenlon, A.; Chang, P.L.; Damtew, Z.M.; Muleta, A.; Carrasquilla-Garcia, N.; Kim, D.; Nguyen, H.P.; Suryawanshi, V.; Krieg, C.P.; Yadav, S.K.; et al. Global-level population genomics reveals differential effects of geography and phylogeny on horizontal gene transfer in soil bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 15200–15209. [Google Scholar] [CrossRef] [PubMed]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-Oxidizing Bacteria: A Model for Molecular Microbial Ecology. Annu. Rev. Microbiol. 2001, 55, 485–529. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-I.; Choi, H.-K.; Lee, S.-W.; Orwin, P.M.; Kim, J.; LaRoe, S.L.; Kim, T.-G.; O’Neil, J.; Leadbetter, J.R.; Hur, C.-G.; et al. Complete Genome Sequence of the Metabolically Versatile Plant Growth-Promoting Endophyte Variovorax paradoxus S110. J. Bacteriol. 2011, 193, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-L.; Yang, W.-L.; Fang, W.-W.; Zhao, Y.-X.; Guo, L.; Dai, Y.-J. The Plant Growth-Promoting RhizobacteriumVariovorax boronicumulansCGMCC 4969 Regulates the Level of Indole-3-Acetic Acid Synthesized from Indole-3-Acetonitrile. Appl. Environ. Microbiol. 2018, 84, 00298-18. [Google Scholar] [CrossRef]
- Smith, T.J.; Font, M.E.; Kelly, C.M.; Sondermann, H.; O’Toole, G.A. An N-terminal retention module anchors the giant adhesin LapA of Pseudomonas fluorescens at the cell surface: A novel subfamily of type I secretion systems. J. Bacteriol. 2018, 200, e00734-17. [Google Scholar] [CrossRef]
- Smith, T.J.; Sondermann, H.; O’Toole, G.A. Type 1 Does the Two-Step: Type 1 Secretion Substrates with a Functional Periplasmic Intermediate. J. Bacteriol. 2018, 200, 00168-18. [Google Scholar] [CrossRef]
- Chatterjee, D.; Boyd, C.D.; O’Toole, G.A.; Sondermann, H. Structural Characterization of a Conserved, Calcium-Dependent Periplasmic Protease from Legionella pneumophila. J. Bacteriol. 2012, 194, 4415–4425. [Google Scholar] [CrossRef]
- Boyd, C.D.; Smith, T.J.; El-Kirat-Chatel, S.; Newell, P.D.; Dufrêne, Y.F.; O’Toole, G.A. Structural features of the Pseudomonas fluorescens biofilm adhesin LapA required for LapG-dependent cleavage, biofilm formation, and cell surface localization. J. Bacteriol. 2014, 196, 2775–2788. [Google Scholar] [CrossRef]
- Fuche, F.; Vianney, A.; Andrea, C.; Doublet, P.; Gilbert, C. Functional Type 1 Secretion System Involved in Legionella pneumophila Virulence. J. Bacteriol. 2015, 197, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Marchler-Bauer, A. NCBI’s conserved domain database and tools for protein domain analysis. Curr. Protoc. Bioinform. 2020, 69, e90. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Fullner, K.J.; Clayton, R.; Sexton, J.A.; Rogers, M.B.; Calia, K.E.; Calderwood, S.B.; Fraser, C.; Mekalanos, J.J. Identification of a Vibrio cholerae RTX toxin gene cluster that is tightly linked to the cholera toxin prophage. Proc. Natl. Acad. Sci. USA 1999, 96, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Suarez, G.; Khajanchi, B.K.; Sierra, J.C.; Erova, T.E.; Sha, J.; Chopra, A.K. Actin cross-linking domain of Aeromonas hydrophila repeat in toxin A (RtxA) induces host cell rounding and apoptosis. Gene 2012, 506, 369–376. [Google Scholar] [CrossRef]
- Gerlach, R.G.; Cláudio, N.; Rohde, M.; Jäckel, D.; Wagner, C.; Hensel, M. Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell Microbiol. 2008, 11, 2364–2376. [Google Scholar] [CrossRef]
- Luo, Y.; Frey, E.A.; Pfuetzner, R.A.; Creagh, A.L.; Knoechel, D.G.; Haynes, C.A.; Finlay, B.B.; Strynadka, N.C.J. Crystal structure of enteropathogenic Escherichia coli intimin–receptor complex. Nat. Cell Biol. 2000, 405, 1073–1077. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Repeat | aa | Features | Genus | Repeat | aa | Features | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aeromonas | A1 | 254 | L | DDVP | Vibrio | Vi1 | 144 | S | het | DDKP | |||
| A2 | 260 | L | DDAP | Vi2 | 142 | S | het | DDxP | |||||
| A3 | 178 | S | DDGP | ||||||||||
| A4 | 171 | S | DDVP | Nitrosomonas | N1 | 161 | S | het | DDGP | ||||
| A5 | 166 | S | DDVP | N2 | 137 | S | DDTP | ||||||
| A6 | 158 | S | DDGP | N3 | 172 | S | DDGP | ||||||
| A7 | 169 | S | solo | DDGP | |||||||||
| A8 | 204 | S | DDGP | Variovorax | Va1 | 167 | S | DDGP | |||||
| A9 | 267 | L | DDMP | Va2 | 177 | S | DDGP | ||||||
| A10 | 205 | S | DDGP | ||||||||||
| A11 | 176 | S | DDGP | Bradyrhizobium | Rb1 | 181 | S | DDGP | |||||
| A12 | 144 | S | solo | DDTP | Rb2 | 200 | S | DDGP | |||||
| A13 | 304 | L | solo | DDGP | Rb3 | 215 | S | solo | DDGP | ||||
| A14 | 219 | S | DDGP | Rb4 | 186 | S | DSGP | ||||||
| A15 | 138 | S | solo | DDTP | Rb5 | 172 | S | DDGP | |||||
| A16 | 183 | S | solo | DDGP | Rb6 | 185 | S | DDGP | |||||
| Rb7 | 181 | S | DDGP | ||||||||||
| Legionella | L1 | 179 | S | DDGP | Rb8 | 177 | S | DDGP | |||||
| L2 | 185 | S | DDGP | ||||||||||
| L3 | 180 | S | DDGP | Mesorhizobium | Rm0 | 267 | L | solo | DDVP | ||||
| L4 | 174 | S | DDGP | Rm1 | 269 | L | DDAP | ||||||
| L5 | 153 | S | DDTP | Rm2 | 172 | S | DDIP | ||||||
| L6 | 168 | S | DDGP | Rm3 | 153 | S | DDGP | ||||||
| L7 | 183 | S | DDGP | Rm4 | 195 | S | DDGP | ||||||
| L8 | 153 | S | solo | DDVP | Rm5 | 243 | L | DDVP | |||||
| L9 | 174 | S | solo | DDGP | Rm6 | 245 | L | DDVP | |||||
| L10 | 155 | S | DDGP | Rm7 | 157 | S | DDGP | ||||||
| L11 | 247 | L | DDVP | Rm8 | 189 | S | DDGP | ||||||
| Rm9 | 269 | L | DDVP | ||||||||||
| Pseudomonas | P1 | 153 | S | DDTP | Rm10 | 162 | S | solo | DTGP | ||||
| P2 | 154 | S | DDGP | Rm11 | 160 | S | DVGP | ||||||
| P3 | 182 | S | DDGP | Rm12 | 157 | S | DDGP | ||||||
| P4 | 185 | S | DDGP | Rm13 | 180 | S | DDGP | ||||||
| P5 | 136 | S | solo | DDTP | Rm14 | 244 | L | DDVP | |||||
| P6 | 196 | S | DDGP | ||||||||||
| P7 | 180 | S | DDGP | Sinorhizobium | Rs1 | 183 | S | DDGP | |||||
| P8 | 179 | S | solo | DDGP | Rs2 | 177 | S | DDGP | |||||
| P9 | 159 | S | het | DDGP | |||||||||
| P10 | 245 | L | DDVP | Multispecies | M1 | 154 | S | DDGP | |||||
| P11 | 244 | L | DDVP | M2 | 168 | S | DDTP | ||||||
| P12 | 178 | S | DDTP | ||||||||||
| P13 | 170 | S | DDTP | Halomonas | H1 | 184 | S | DDGP | |||||
| P15 | 248 | L | DDVP | H2 | 201 | S | DDGP | ||||||
| P16 | 190 | S | solo | DDAP | |||||||||
| P17 | 202 | S | het | DDGP | |||||||||
| P18 | 144 | S | solo | DDTP | |||||||||
| P19 | 309 | L | solo | DDAP | |||||||||
| P20 | 158 | S | het | DDAP | |||||||||
| P21 | 183 | S | solo | DDGP | |||||||||
| P22 | 268 | L | het | DDMP | |||||||||
| P23 | 204 | S | DDGP | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Nocera, P.; De Gregorio, E. Growth by Insertion: The Family of Bacterial DDxP Proteins. Int. J. Mol. Sci. 2020, 21, 9184. https://doi.org/10.3390/ijms21239184
Di Nocera P, De Gregorio E. Growth by Insertion: The Family of Bacterial DDxP Proteins. International Journal of Molecular Sciences. 2020; 21(23):9184. https://doi.org/10.3390/ijms21239184
Chicago/Turabian StyleDi Nocera, Pierpaolo, and Eliana De Gregorio. 2020. "Growth by Insertion: The Family of Bacterial DDxP Proteins" International Journal of Molecular Sciences 21, no. 23: 9184. https://doi.org/10.3390/ijms21239184
APA StyleDi Nocera, P., & De Gregorio, E. (2020). Growth by Insertion: The Family of Bacterial DDxP Proteins. International Journal of Molecular Sciences, 21(23), 9184. https://doi.org/10.3390/ijms21239184