SMAD3 Hypomethylation as a Biomarker for Early Prediction of Colorectal Cancer

 , and
, and
Abstract
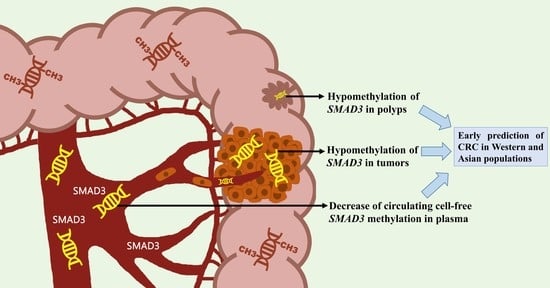
1. Introduction
2. Results
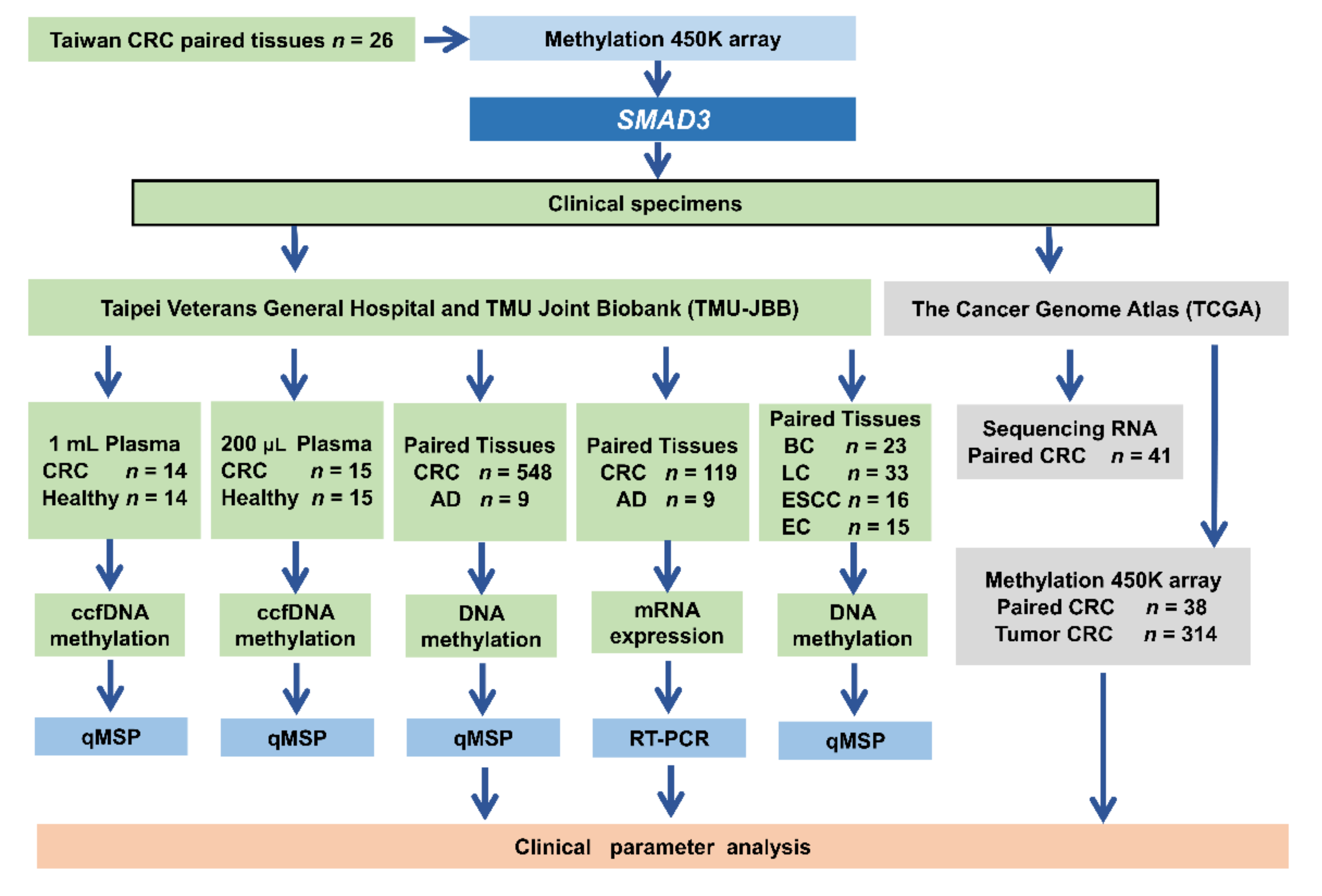
2.1. SMAD3 Was Identified from Taiwanese and Western Patients with CRC through Genome-Wide Methylation Analysis
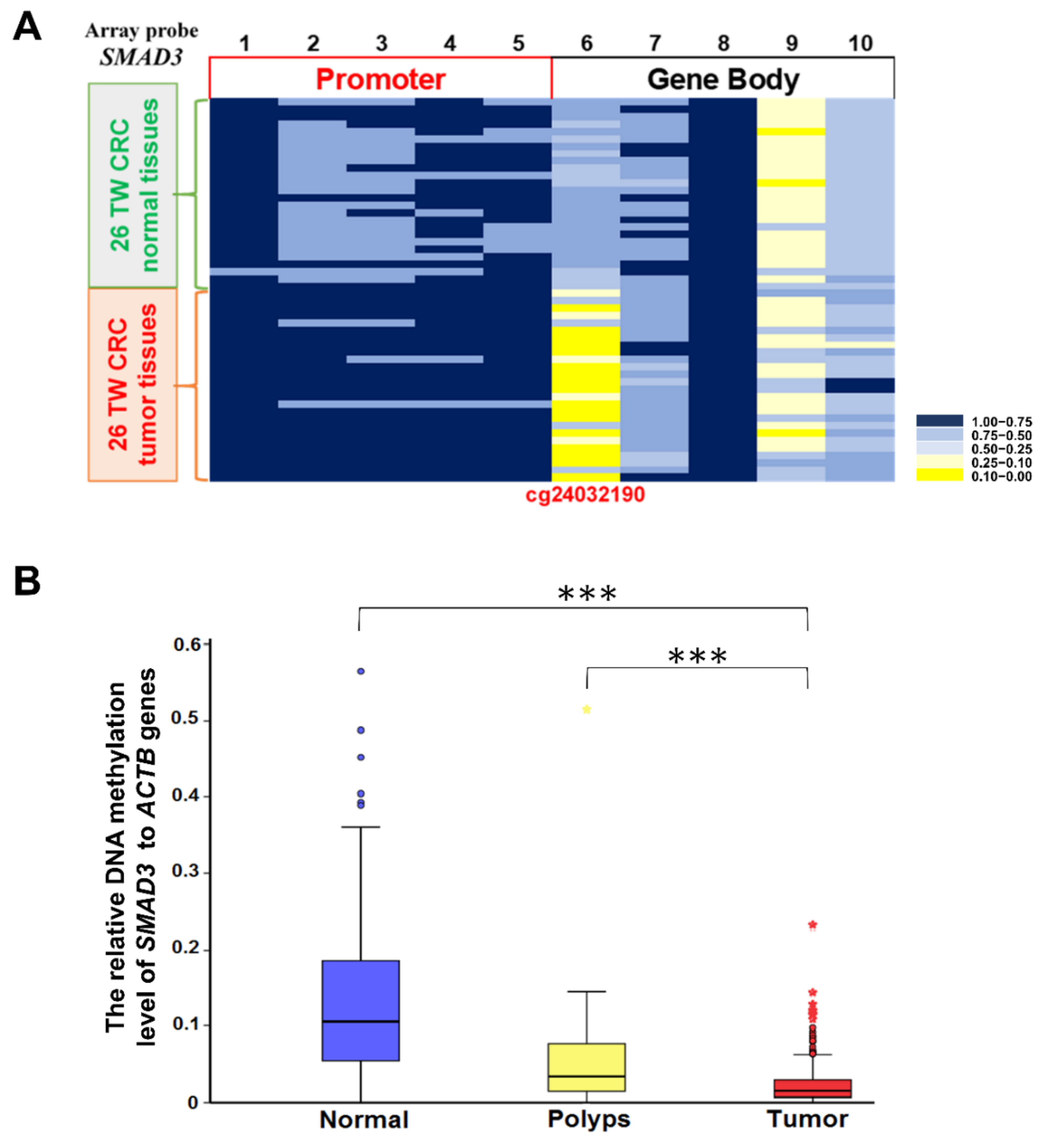
2.2. Methylation Level of SMAD3 in Tissues from Taiwanese Patients with CRC
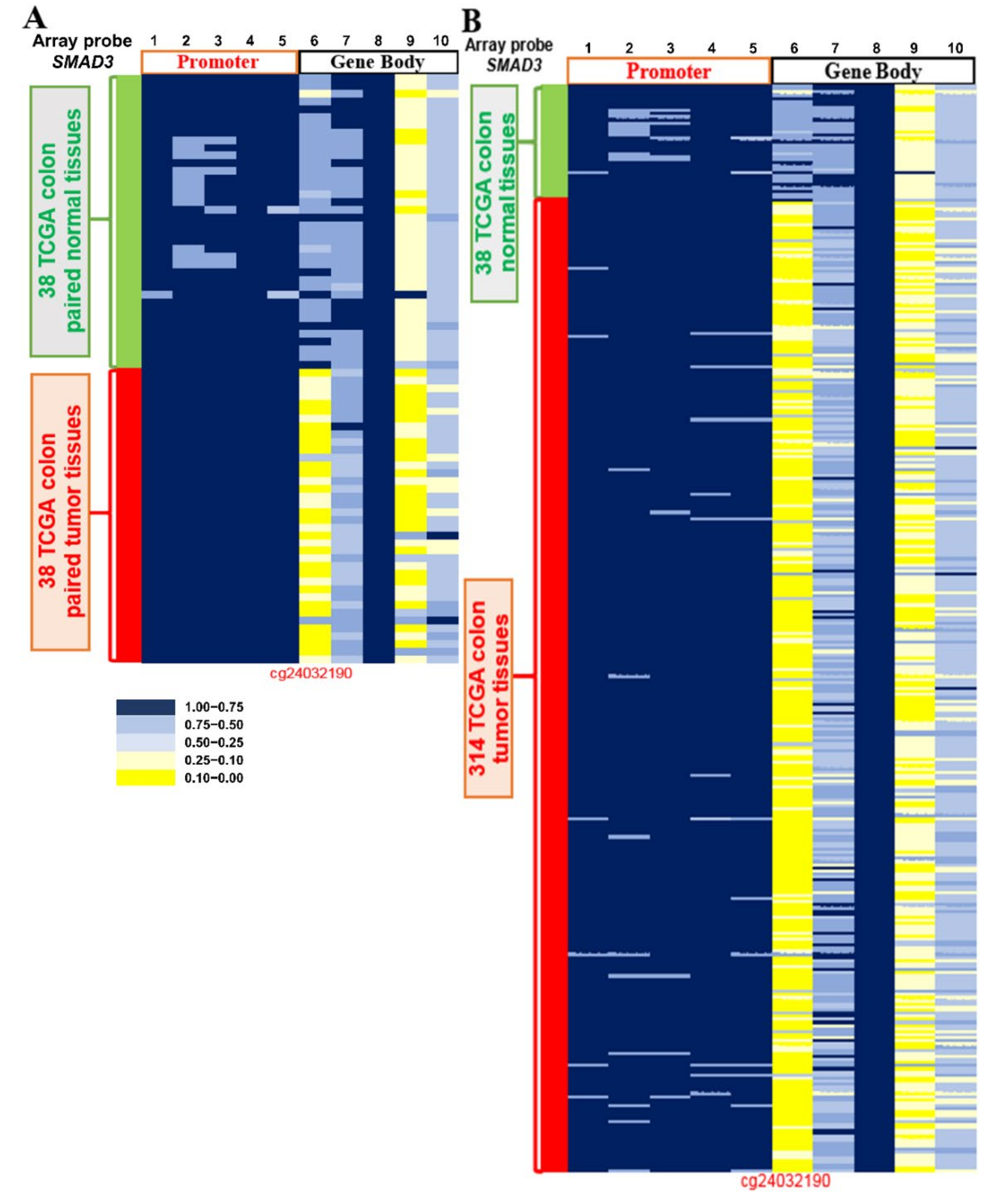
2.3. Methylation Level of SMAD3 in CRC Tissues from TCGA Datasets
2.4. Methylation Level of ccfDNA of SMAD3 in Plasma
2.5. SMAD3 mRNA Expression in Taiwanese CRC Paired Tissues and the TCGA Dataset
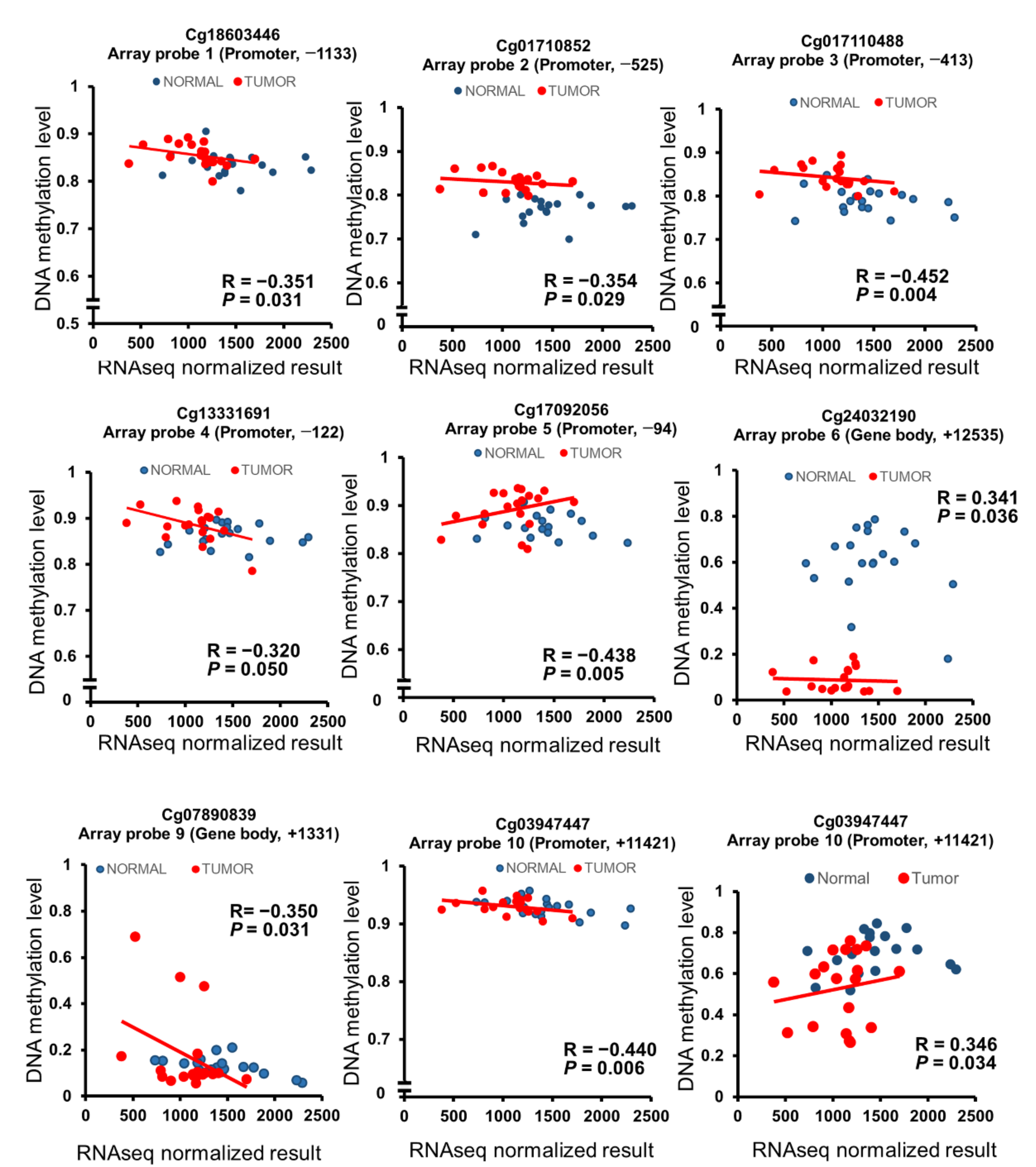
2.6. Clinical Characteristics of DNA Methylation and mRNA Expression
2.7. Methylation Level of SMAD3 in Different Cancers
2.8. Correlation between Prognosis and SMAD3 Methylation Level
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue and Plasma Collection
4.2. Genomic DNA, ccfDNA, and RNA Extraction
4.3. Ethical Approval and Consent to Participate
4.4. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
4.5. TaqMan Quantitative Methylation-Specific PCR (QMSP)
4.6. Genome-Wide Methylation Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CRC | colorectal cancer |
| TCGA | The Cancer Genome Atlas |
| ccfDNA | circulating cell-free DNA |
| BEND5 | BEN Domain Containing 5 |
| ACOT7 | Acyl-CoA Thioesterase 7 |
| RASA3 | RAS P21 Protein Activator 3 |
| B3GNTL1 | UDP-GlcNAc:BetaGal Beta-1,3-N-Acetylglucosaminyltransferase Like 1 |
| PIGB | Phosphatidylinositol Glycan Anchor Biosynthesis Class B |
| MAP3K5 | Mitogen-Activated Protein Kinase Kinase Kinase 5 |
| LPIN 1 | Lipin 1 |
| MYBPC3 | Myosin Binding Protein C3 |
| FDA | Food and Drug Administration |
| mcfDNA | methylated circulating cell-free DNA |
| SMAD3 | Mothers Against Decapentaplegic Homolog 3 |
| TGF-β | Transforming Growth Factor-β |
| PAX6 | Paired Box 6 |
| MSI | Microsatellite Instability |
| KRAS | Kirsten Rat Sarcoma Viral Oncogene Homologue |
| ESCC | Esophageal Squamous Cell Cancer |
| Lung AD | Lung Adenocarcinoma |
| Lung SQ | Lung Squamous Cell Carcinoma |
| IBD | inflammatory bowel disease |
| LINE-1 | Long Interspersed Nuclear Element-1 |
| SNP | single-nucleotide polymorphisms |
| qRT-PCR | quantitative reverse-transcription polymerase chain reaction |
| qMSP | TaqMan quantitative methylation-specific PCR |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| ACTB | Beta-actin |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef]
- Ministry of Health and Welfare 2018, Taiwan, R.O.C. 2018. Available online: https://www.hpa.gov.tw/Pages/Detail.aspx?nodeid=269&pid=10227 (accessed on 2 August 2020).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683. [Google Scholar] [CrossRef]
- Cancer.Net. 2019. Available online: https://www.cancer.net/cancer-types/colorectal-cancer/statistics (accessed on 20 May 2020).
- Simon, K. Colorectal cancer development and advances in screening. Clin. Interv. Aging 2016, 11, 967–976. [Google Scholar] [PubMed]
- Weisenberger, D.J.; Liang, G.; Lenz, H.J. DNA methylation aberrancies delineate clinically distinct subsets of colorectal cancer and provide novel targets for epigenetic therapies. Oncogene 2018, 37, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Werner, R.J.; Kelly, A.D.; Issa, J.-P.J. Epigenetics and Precision Oncology. Cancer J. 2017, 23, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Bergheim, J.; Semaan, A.; Gevensleben, H.; Groening, S.; Knoblich, A.; Dietrich, J.; Dietrich, D. Potential of quantitative SEPT9 and SHOX2 methylation in plasmatic circulating cell-free DNA as auxiliary staging parameter in colorectal cancer: A prospective observational cohort study. Br. J. Cancer 2018, 118, 1217–1228. [Google Scholar] [CrossRef]
- Hao, T.B.; Shi, W.; Shen, X.J.; Qi, J.; Wu, X.H.; Wu, Y.; Ju, S.Q. Circulating cell-free DNA in serum as a biomarker for diagnosis and prognostic prediction of colorectal cancer. Br. J. Cancer 2014, 111, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Williams, M.; Cheng, Y.Y.; Leung, W.K. Roles of Methylated DNA Biomarkers in Patients with Colorectal Cancer. Dis. Markers 2019, 2019, 2673543. [Google Scholar] [CrossRef]
- Lin, R.K.; Hung, W.Y.; Huang, Y.F.; Chang, Y.J.; Lin, C.H.; Chen, W.Y.; Tsai, S.F. Hypermethylation of BEND5 contributes to cell proliferation and is a prognostic marker of colorectal cancer. Oncotarget 2017, 8, 113431–113443. [Google Scholar] [CrossRef]
- Amatu, A.; Schirripa, M.; Tosi, F.; Lonardi, S.; Bencardino, K.; Bonazzina, E.; Bergamo, F. High Circulating Methylated DNA Is a Negative Predictive and Prognostic Marker in Metastatic Colorectal Cancer Patients Treated with Regorafenib. Front. Oncol. 2019, 9, 622. [Google Scholar] [CrossRef] [PubMed]
- Tang PM, K.; Zhou, S.; Meng, X.M.; Wang, Q.M.; Li, C.J.; Lian, G.Y.; To, K.F. Smad3 promotes cancer progression by inhibiting E4BP4-mediated NK cell development. Nat. Commun. 2017, 8, 14677. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Zhang, Q.; Hu, Y.; Zhang, T.; Li, J.; Liu, Z.; Li, B. Investigating the mechanism by which SMAD3 induces PAX6 transcription to promote the development of non-small cell lung cancer. Respir. Res. 2018, 19, 262. [Google Scholar] [CrossRef] [PubMed]
- Frigola, J.; Solé, X.; Paz, M.F.; Moreno, V.; Esteller, M.; Capellà, G.; Peinado, M.A. Differential DNA hypermethylation and hypomethylation signatures in colorectal cancer. Hum. Mol. Genet. 2004, 14, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, B.; Zhang, J.; Yang, X.; Zhang, G.; Yang, S.; Jing, H. Expression level of ACOT7 influences the prognosis in acute myeloid leukemia patients. Cancer Biomark. 2019, 26, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Fan, X.; He, L.; Zhou, D. Methylation patterns of RASA3 associated with clinicopathological factors in hepatocellular carcinoma. J. Cancer 2018, 9, 2116–2122. [Google Scholar] [CrossRef]
- Sandanger, T.M.; Nøst, T.H.; Guida, F.; Rylander, C.; Campanella, G.; Muller, D.C.; Vermeulen, R. DNA methylation and associated gene expression in blood prior to lung cancer diagnosis in the Norwegian Women and Cancer cohort. Sci. Rep. 2018, 8, 16714. [Google Scholar] [CrossRef]
- Murakami, Y.; Nguyen, T.T.M.; Baratang, N.; Raju, P.K.; Knaus, A.; Ellard, S.; Kamei, A. Mutations in PIGB Cause an Inherited GPI Biosynthesis Defect with an Axonal Neuropathy and Metabolic Abnormality in Severe Cases. Am. J. Hum. Genet. 2019, 105, 384–394. [Google Scholar] [CrossRef]
- Pressinotti, N.C.; Klocker, H.; Schäfer, G.; Luu, V.D.; Ruschhaupt, M.; Kuner, R.; Sültmann, H. Differential expression of apoptotic genes PDIA3 and MAP3K5 distinguishes between low- and high-risk prostate cancer. Mol. Cancer 2009, 8, 130. [Google Scholar] [CrossRef]
- Nunes, D.; Nogueira, C.; Lopes, A.; Chaves, P.; Rodrigues, E.; Cardoso, T.; Vilarinho, L. LPIN1 deficiency: A novel mutation associated with different phenotypes in the same family. Mol. Genet. Metab. Rep. 2016, 9, 29–30. [Google Scholar] [CrossRef][Green Version]
- Carrier, L.; Mearini, G.; Stathopoulou, K.; Cuello, F. Cardiac myosin-binding protein C (MYBPC3) in cardiac pathophysiology. Gene 2015, 573, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Chiba, H.; Kakuta, Y.; Kinouchi, Y.; Kawai, Y.; Watanabe, K.; Nagao, M.; Kanazawa, Y. Allele-specific DNA methylation of disease susceptibility genes in Japanese patients with inflammatory bowel disease. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.A.; Ananthakrishnan, A.N.; Gardet, A.; Stevens, C.R.; Korzenik, J.R.; Sands, B.E.; Yajnik, V. SMAD3 gene variant is a risk factor for recurrent surgery in patients with Crohn’s disease. J. Crohns Colitis 2014, 8, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.; McGovern, D.P.; Barrett, J.C.; Wang, K.; Radford-Smith, G.L.; Ahmad, T.; Anderson, C.A. Genome-wide meta-analysis increases to 71 the number of confirmed Crohn’s disease susceptibility loci. Nat. Genet. 2010, 42, 1118–1125. [Google Scholar] [CrossRef]
- Hur, K.; Cejas, P.; Feliu, J.; Moreno-Rubio, J.; Burgos, E.; Boland, C.R.; Goel, A. Hypomethylation of long interspersed nuclear element-1 (LINE-1) leads to activation of proto-oncogenes in human colorectal cancer metastasis. Gut 2014, 63, 635–646. [Google Scholar] [CrossRef]
- Chang, S.C.; Liew, P.L.; Ansar, M.; Lin, S.Y.; Wang, S.C.; Hung, C.S.; Lin, R.K. Hypermethylation and decreased expression of TMEM240 are potential early-onset biomarkers for colorectal cancer detection, poor prognosis, and early recurrence prediction. Clin. Epigenetics 2020, 12, 67. [Google Scholar] [CrossRef]
- Sato, T.; Issa, J.J.; Kropf, P. DNA Hypomethylating Drugs in Cancer Therapy. Cold Spring Harb. Perspect. Med. 2017, 7. [Google Scholar] [CrossRef]
- Sodir, N.M.; Chen, X.; Park, R.; Nickel, A.E.; Conti, P.S.; Moats, R.; Laird, P.W. Smad3 deficiency promotes tumorigenesis in the distal colon of ApcMin/+ mice. Cancer Res. 2006, 66, 8430–8438. [Google Scholar] [CrossRef]
- Han, S.U.; Kim, H.T.; Kim, Y.S.; Park, Y.S.; Bang, Y.J.; Yang, H.K.; Kim, S.J. Loss of the Smad3 expression increases susceptibility to tumorigenicity in human gastric cancer. Oncogene 2004, 23, 1333–1341. [Google Scholar] [CrossRef]
- Schmidt, B.; Weickmann, S.; Witt, C.; Fleischhacker, M. Improved Method for Isolating Cell-Free DNA. Clin. Chem. 2005, 51, 1561–1563. [Google Scholar] [CrossRef]
- Ali, N.; Rampazzo, R.D.C.P.; Costa, A.D.T.; Krieger, M. A. Current Nucleic Acid Extraction Methods and Their Implications to Point-of-Care Diagnostics. BioMed Res. Int. 2017, 2017, 9306564. [Google Scholar] [CrossRef] [PubMed]
- Boldrin, E.; Curtarello, M.; Dallan, M.; Alfieri, R.; Realdon, S.; Fassan, M.; Saggioro, D. Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Kriseman, M.; Monsivais, D.; Agno, J.; Masand, R.P.; Creighton, C.J.; Matzuk, M.M. Uterine double-conditional inactivation of Smad2 and Smad3 in mice causes endometrial dysregulation, infertility, and uterine cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 3873–3882. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total n | SMAD3 Methylation | Total n | SMAD3 mRNA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low n (%) a | Normal n (%) | Low n (%) b | Moderate n (%) | High n (%) | ||||||||
| Overall | 548 | 501 | (91.4) | 47 | (8.6) | 119 | 51 | (42.9) | 33 | (27.7) | 35 | (29.4) |
| Age | ||||||||||||
| <65 | 240 | 213 | (88.8) | 27 | (11.2) | 46 | 23 | (50.0) | 11 | (23.9) | 12 | (26.1) |
| 65 | 287 | 268 | (93.4) | 19 | (6.6) | 60 | 24 | (40.0) | 19 | (31.7) | 17 | (28.3) |
| Sex | ||||||||||||
| Male | 314 | 289 | (92.0) | 25 | (8.0) | 59 | 24 | (40.7) | 19 | (32.2) | 16 | (27.1) |
| Female | 214 | 193 | (90.2) | 21 | (9.8) | 54 | 26 | (48.1) | 13 | (24.1) | 15 | (27.8) |
| Tumor Type | ||||||||||||
| Adeno | 492 | 451 | (91.7) | 41 | (8.3) | 104 | 45 | (41.7) | 32 | (29.6) | 27 | (25.0) |
| Mucinous | 56 | 50 | (89.3) | 6 | (10.7) | 4 | 2 | (50.0) | 0 | (0) | 2 | (50.0) |
| Tumor Stage | ||||||||||||
| 0 and I | 53 | 47 | (88.7) | 6 | (11.3) | 10 | 4 | (40.0) | 3 | (30.0) | 3 | (30.0) |
| II, III, and IV | 451 | 411 | (91.1) | 40 | (8.9) | 98 | 44 | (44.9) | 28 | (28.6) | 26 | (26.5) |
| Tumor Size | ||||||||||||
| T0–T1 | 34 | 31 | (91.2) | 3 | (8.8) | 6 | 2 | (33.3) | 1 | (16.7) | 3 | (50.0) |
| T2–T4 | 485 | 442 | (91.1) | 43 | (8.9) | 100 | 45 | (45.0) | 29 | (29.0) | 26 | (26.0) |
| Regional lymph nodes metastasis | ||||||||||||
| N = 0 | 270 | 250 | (92.6) | 20 | (7.4) | 57 | 25 | (43.9) | 16 | (28.1) | 16 | (28.1) |
| N ≥ 1 | 249 | 223 | (89.6) | 26 | (10.4) | 51 | 23 | (45.1) | 15 | (29.4) | 13 | (25.5) |
| Distant metastasis | ||||||||||||
| M = 0 | 398 | 361 | (90.7) | 37 | (9.3) | 76 | 36 | (47.4) | 21 | (27.6) | 19 | (25.0) |
| M ≥ 1 | 102 | 93 | (91.2) | 9 | (8.8) | 28 | 11 | (39.3) | 8 | (28.6) | 9 | (32.1) |
| Differentiation grade | ||||||||||||
| Well/Moderate | 481 | 439 | (91.3) | 42 | (8.7) | 101 | 45 | (44.6) | 30 | (29.7) | 26 | (25.7) |
| Poor/undifferentiation | 33 | 29 | (87.9) | 4 | (12.1) | 6 | 2 | (33.3) | 1 | (16.7) | 3 | (50.0) |
| Vascular invasion | ||||||||||||
| No invasion | 408 | 373 | (91.4) | 35 | (8.6) | 60 | 24 | (40.0) | 16 | (26.7) | 20 | (33.3) |
| Invasion | 77 | 70 | (90.9) | 7 | (9.1) | 18 | 8 | (44.4) | 3 | (16.7) | 7 | (38.9) |
| Location | ||||||||||||
| Cecum, appendix | 46 | 43 | (93.5) | 3 | (6.5) | 8 | 2 | (25.0) | 1 | (12.5) | 5 | (62.5) |
| Ascending colon | 88 | 83 | (94.3) | 5 | (5.7) | 13 | 4 | (30.8) | 4 | (30.8) | 5 | (38.5) |
| Transverse colon | 22 | 20 | (90.9) | 2 | (9.1) | 5 | 1 | (20.0) | 0 | (0) | 4 | (80.0) |
| Descending colon | 49 | 45 | (91.8) | 4 | (8.2) | 12 | 6 | (50.0) | 1 | (8.3) | 5 | (41.7) |
| Sigmoid colon | 160 | 142 | (88.8) | 18 | (11.2) | 23 | 12 | (52.2) | 6 | (26.1) | 5 | (21.7) |
| Rectum | 129 | 119 | (82.2) | 10 | (7.8) | 18 | 7 | (38.9) | 8 | (44.4) | 3 | (16.7) |
| MSI c | ||||||||||||
| MSS | 50 | 47 | (94.0) | 3 | (6.0) | 38 | 12 | (31.6) | 14 | (36.8) | 12 | (31.6) |
| MSI-L | 6 | 6 | (100.0) | 0 | (0.0) | 5 | 2 | (40.0) | 2 | (40.0) | 1 | (20.0) |
| MSI-H | 9 | 9 | (100.0) | 0 | (0.0) | 7 | 4 | (57.1) | 0 | (0) | 3 | (42.9) |
| TCGA | TAIWAN | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total n | Avgβ (T) | Avgβ (N) | Avgβ (T − N) | Total n | Avg (T) | Avg (N) | Pair T/N a < 0.5 | Pair T = N | Pair T/N > 2 | ||
| Colon | 38 | 0.11 | 0.63 | −0.52 | Colon | 548 | 0.230 | 0.010 | 91.4% | 3.6% | 5.4% |
| ESCC b | 15 | 0.06 | 0.09 | −0.03 | ESCC | 16 | 0.002 | 0.001 | 62.5% | 25% | 12.5% |
| Breast | 87 | 0.13 | 0.11 | 0.02 | Breast | 23 | 0.020 | 0.010 | 30.4% | 47.8% | 21.7% |
| Gastric | 2 | 0.06 | 0.13 | −0.07 | Lung | 33 | 0.002 | 0.003 | 42.4% | 33.3% | 24.4% |
| Liver | 12 | 0.13 | 0.28 | −0.15 | Endometrial | 15 | 0.003 | 0.020 | 0% | 40% | 60% |
| Lung AD c | 29 | 0.17 | 0.19 | −0.02 | |||||||
| Lung SQd | 40 | 0.10 | 0.09 | 0.01 | |||||||
| Pancreatic | 10 | 0.07 | 0.08 | −0.01 | |||||||
| Uterine | 7 | 0.32 | 0.13 | 0.17 | |||||||
| Sequence (5′ to 3′) | Probe | Tm (°C) | Size (bp) | ||
|---|---|---|---|---|---|
| Real-time RT-PCRa | |||||
| GAPDH | Forward | AGCCACATCGCTCAGACAC | #60 | 60 | 66 |
| Reverse | GCCCAATACGACCAAATCC | ||||
| SMAD3 | Forward | GTCTGCAAGATCCCACCA | #79 | 59 | 88 |
| Reverse | AGCCCTGGTTGACCGACT | ||||
| Quantitative Methylation-Specific PCR | |||||
| ACTB | Forward | TGGTGATGGAGGAGGTTTAGTAAGT | 60 | 132 | |
| Reverse | AACCAATAAAACCTACTCCTCCCTTAA | ||||
| TagMan probe | ACCACCACCCAACACACAATAACAAACACA | ||||
| SMAD3 | Forward | GAATAAGGTCGTTAGTTATTATCGT | 54.48 | 172 | |
| Reverse | AATCAAATCTACCCGAATCGAA | ||||
| TaqMan probe | GAAAGAAAGAAAGAAAGTAAATTTTATTTTTAAGCG | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansar, M.; Wang, C.-J.; Wang, Y.-H.; Shen, T.-H.; Hung, C.-S.; Chang, S.-C.; Lin, R.-K. SMAD3 Hypomethylation as a Biomarker for Early Prediction of Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 7395. https://doi.org/10.3390/ijms21197395
Ansar M, Wang C-J, Wang Y-H, Shen T-H, Hung C-S, Chang S-C, Lin R-K. SMAD3 Hypomethylation as a Biomarker for Early Prediction of Colorectal Cancer. International Journal of Molecular Sciences. 2020; 21(19):7395. https://doi.org/10.3390/ijms21197395
Chicago/Turabian StyleAnsar, Muhamad, Chun-Jung Wang, Yu-Han Wang, Tsung-Hua Shen, Chin-Sheng Hung, Shih-Ching Chang, and Ruo-Kai Lin. 2020. "SMAD3 Hypomethylation as a Biomarker for Early Prediction of Colorectal Cancer" International Journal of Molecular Sciences 21, no. 19: 7395. https://doi.org/10.3390/ijms21197395
APA StyleAnsar, M., Wang, C.-J., Wang, Y.-H., Shen, T.-H., Hung, C.-S., Chang, S.-C., & Lin, R.-K. (2020). SMAD3 Hypomethylation as a Biomarker for Early Prediction of Colorectal Cancer. International Journal of Molecular Sciences, 21(19), 7395. https://doi.org/10.3390/ijms21197395