Molecular Players of EF-hand Containing Calcium Signaling Event in Plants


 ,
, 
 and
and
Abstract
1. Introduction
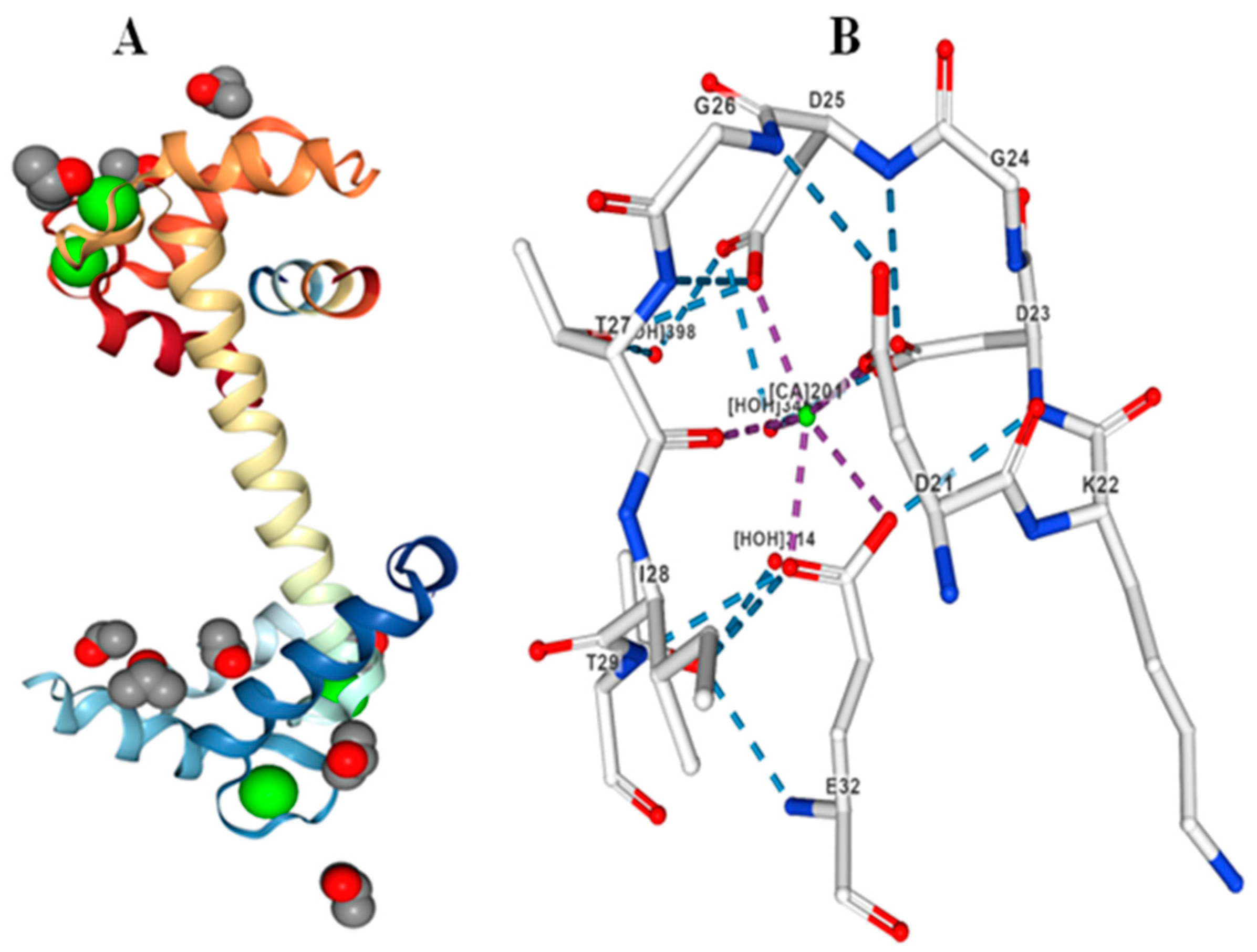
2. Calmodulins (CaMs)
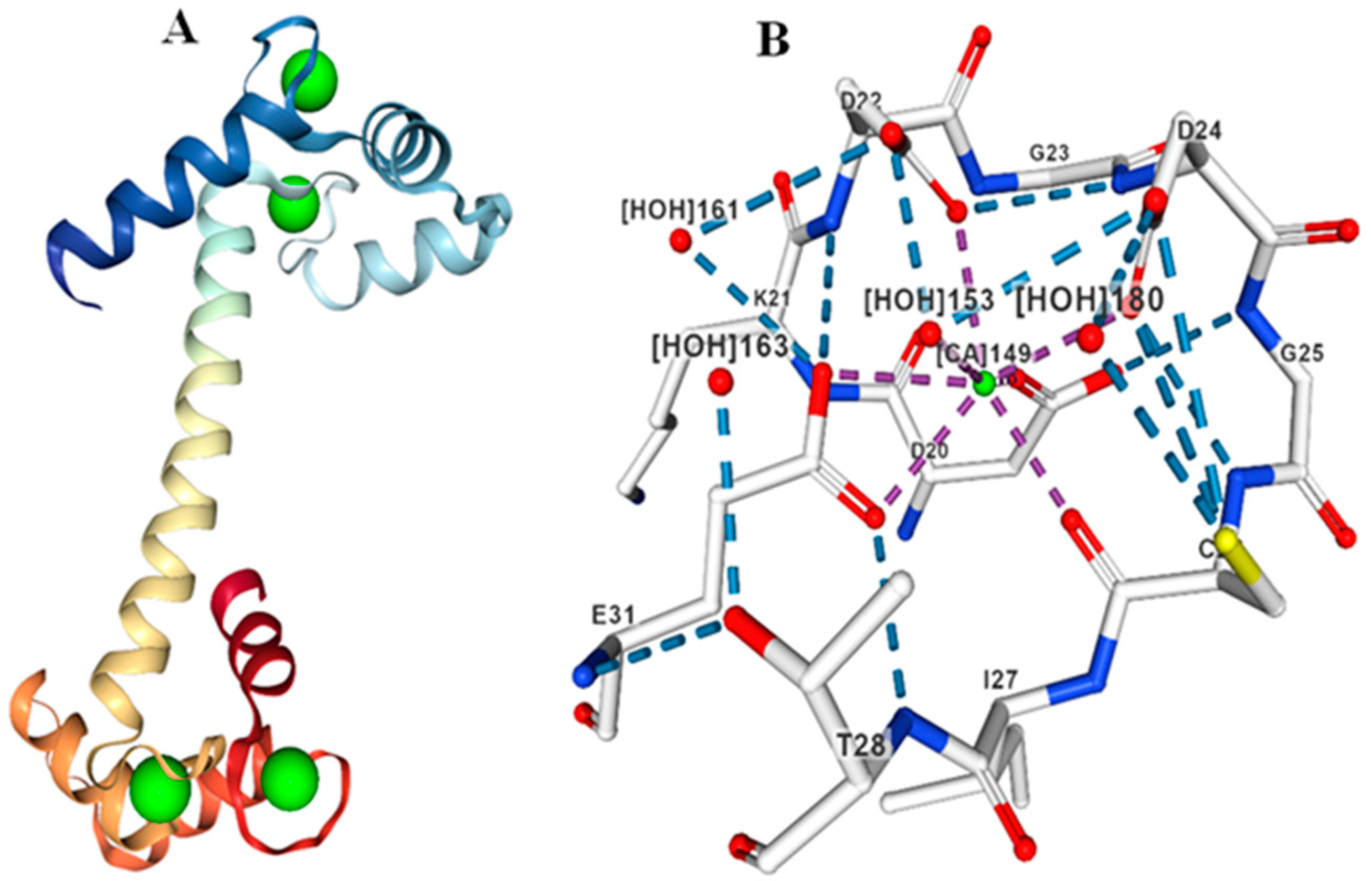
3. Calmodulin-Like (CMLs)
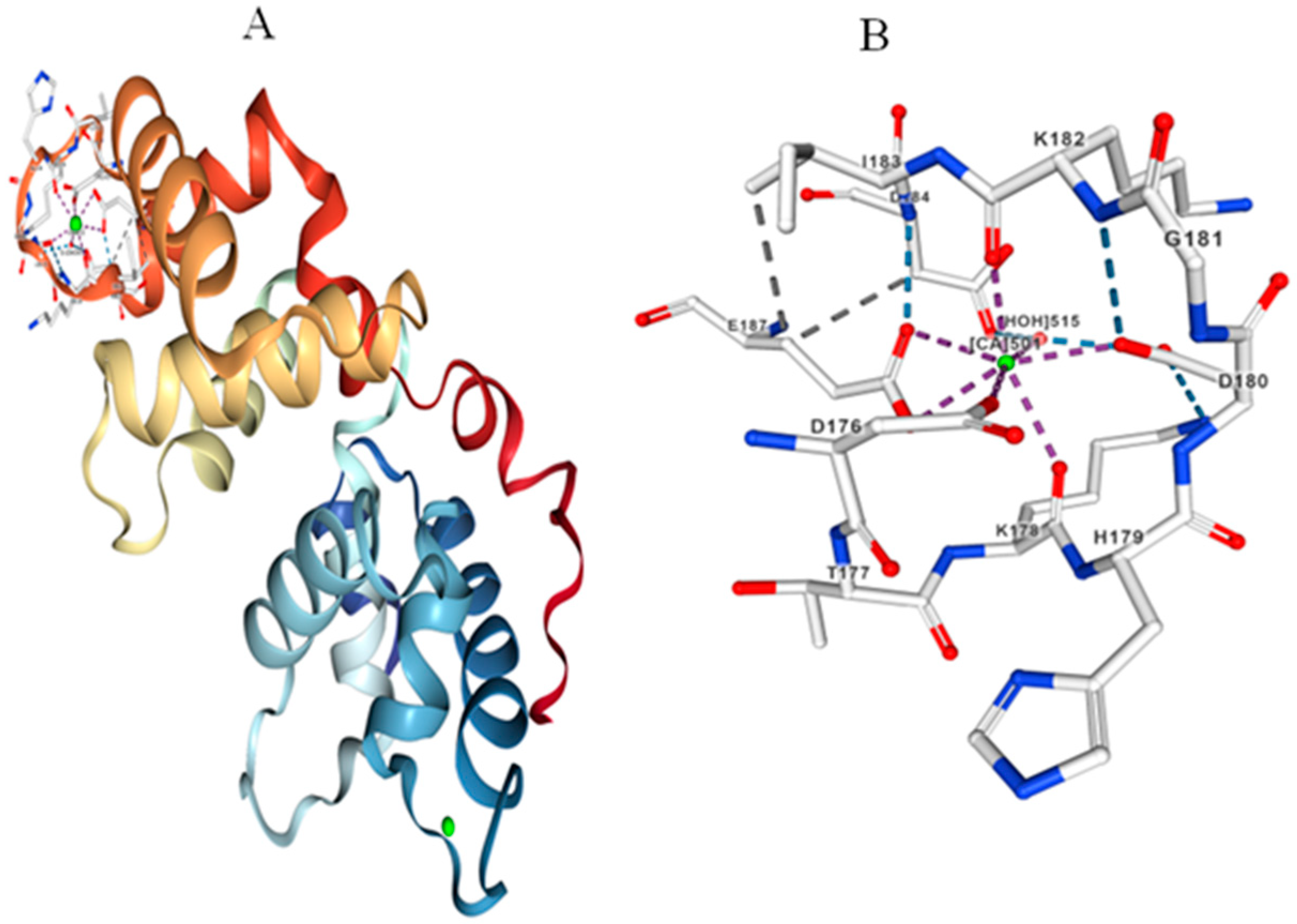
4. Calcineurin B-like (CBL)
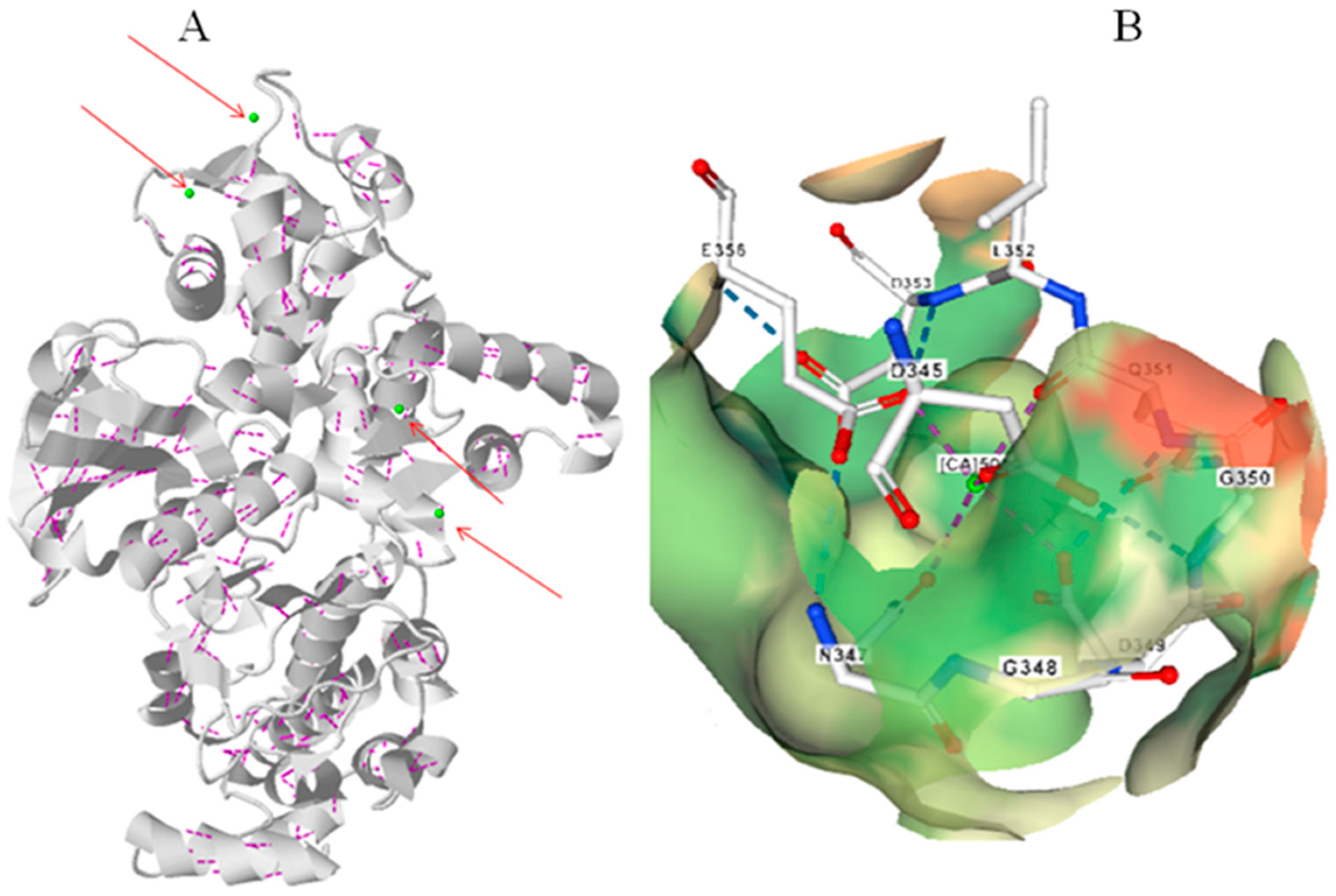
5. Calcium Dependent Protein Kinases (CPKs)
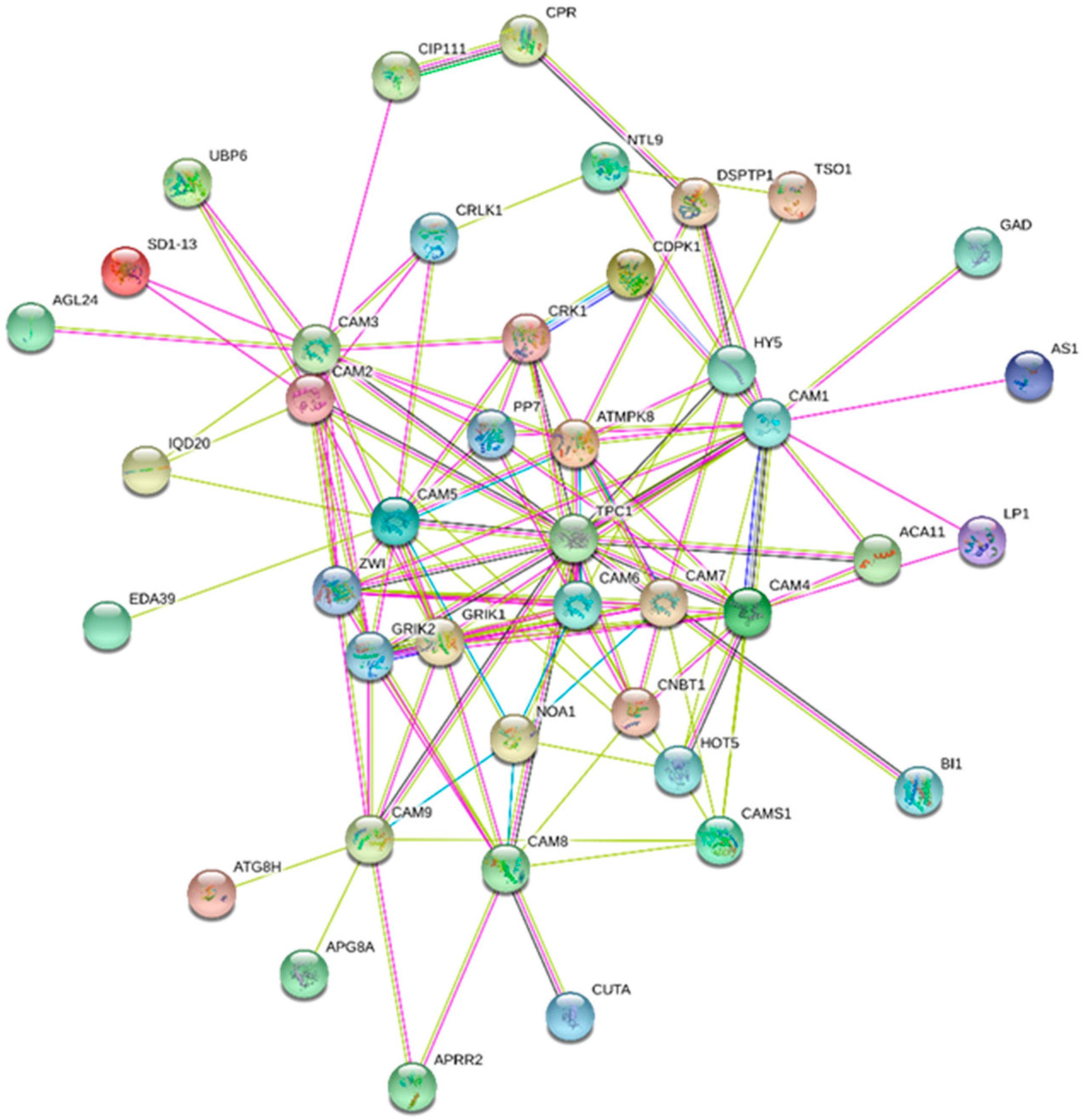
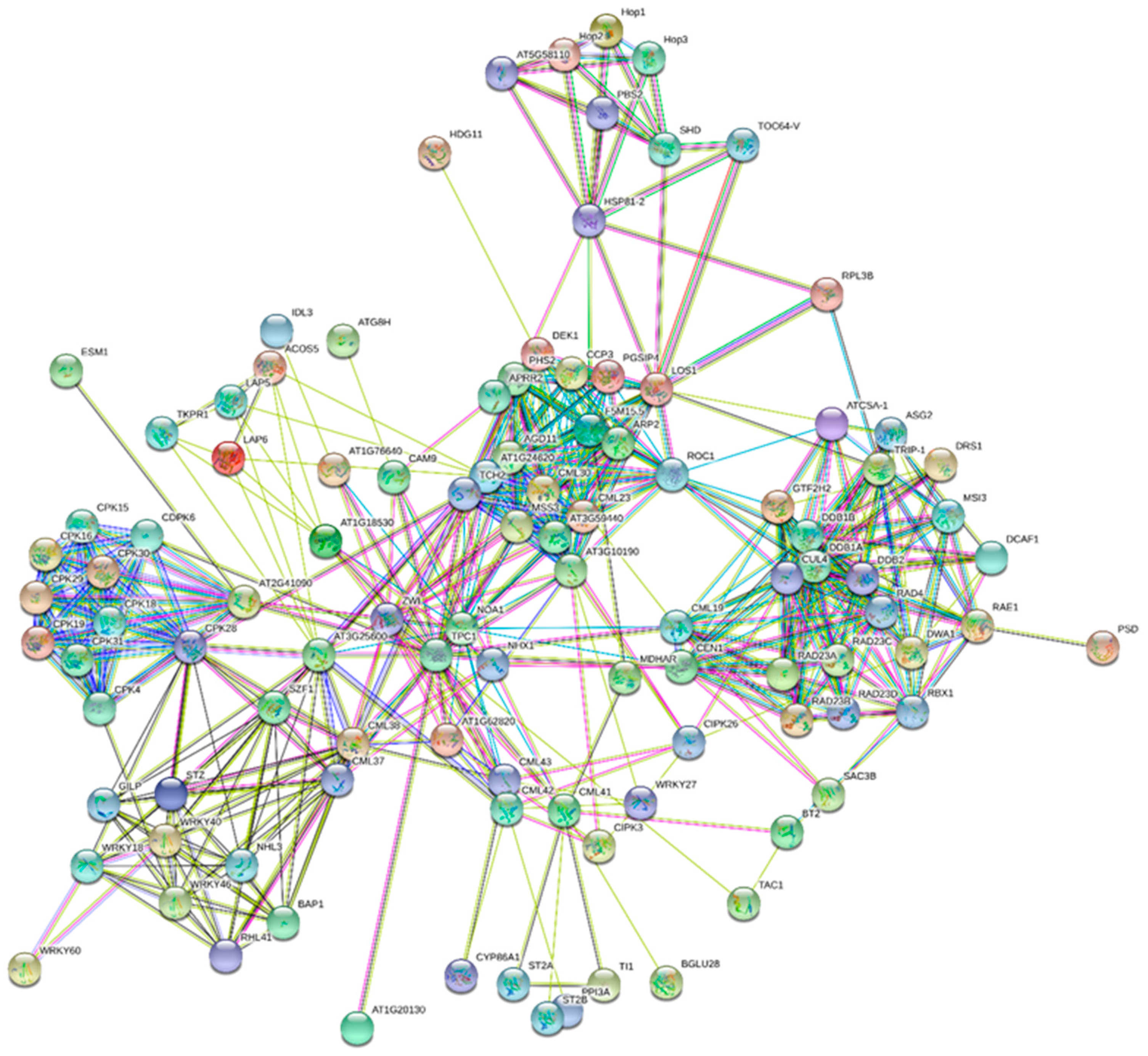
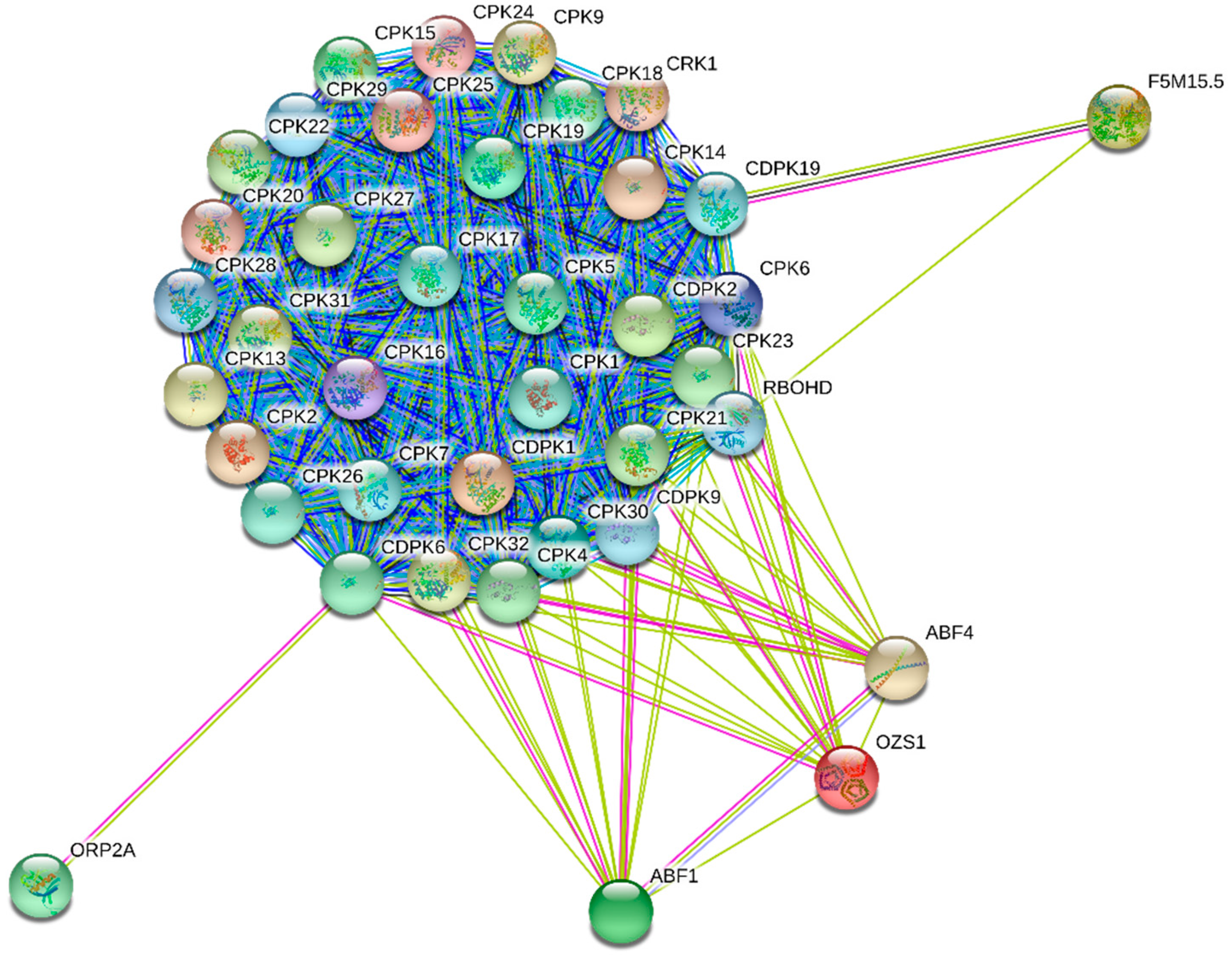
6. Interactome map of CaMs, CMLs, CBLs and CDPKs
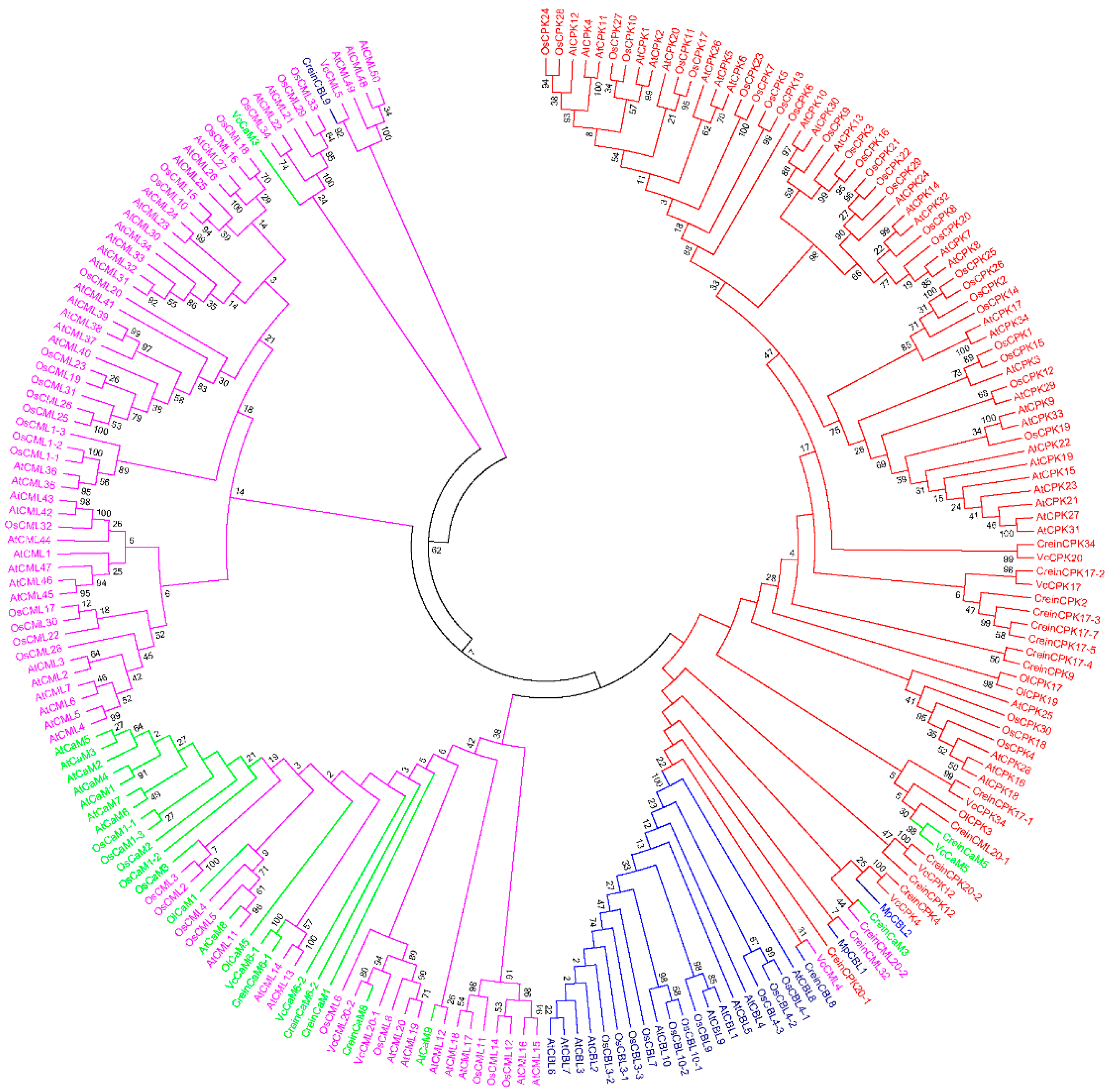
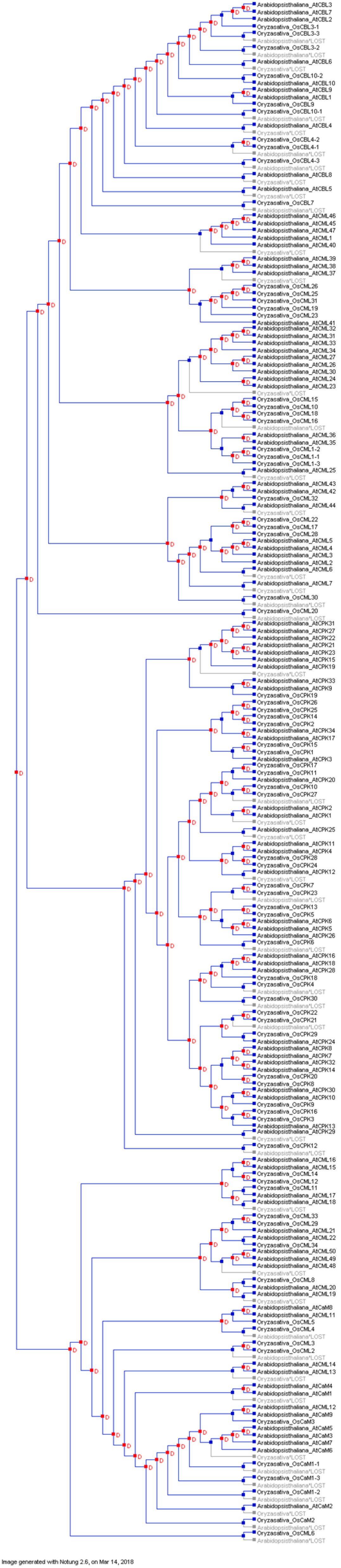
7. Evolution of Calcium Signaling Events
8. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Burstrom, H.G. Calcium and plant growth. Biol. Rev. 1968, 43, 287–316. [Google Scholar] [CrossRef]
- Tuteja, N.; Mahajan, S. Calcium Signaling Network in Plants. Plant Signal. Behav. 2007, 79–85. [Google Scholar] [CrossRef]
- Qin, T.; Li, J.; Yuan, M.; Mao, T. Characterization of the role of calcium in regulating the microtubule-destabilizing activity of MDP25. Plant Signal. Behav. 2012, 7, 708–710. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K. The Cytoskeleton and Its Regulation by Calcium and Protons. Plant Physiol. 2016, 170, 3. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, I.V.; Weber, K. Calcium-Induced Inactivation of Microtubule Formation in Brain Extracts: Presence of a Calcium-Dependent Protease Acting on Polymerization-Stimulating Microtubule-Associated Proteins. Eur. J. Biochem. 1978, 92, 463–470. [Google Scholar] [CrossRef]
- Vater, W.; Bohm, K.J.; Unger, E. Tubulin assembly in the presence of calcium ions and taxol: Microtubule bundling and formation of macrotubule—Ring complexes. Cell Motil. 1998, 36, 76–83. [Google Scholar] [CrossRef]
- Dematry, M.; Morvan, C.; Thellier, M. Calcium and the cell wall. Plant. Cell Environ. 2006, 7, 441–448. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd_Allah, E.F.; Khan, A.L.; Al-Harrasi, A.S. Early Events in Plant Abiotic Stress Signaling: Interplay Between Calcium, Reactive Oxygen Species and Phytohormones. J. Plant Growth Regul. 2018. [Google Scholar] [CrossRef]
- Bootman, M. Calcium Signaling. Cold Cold Spring Harb Perspect Biol. 2012, 4, a011171. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N. Integrated Calcium Signaling in Plants. In Signaling in Plants; Mancuso, S., Balu¿ka, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 29–49. ISBN 978-3-540-89228-1. [Google Scholar]
- Xiong, T.-C.; Bourque, S.; Lecourieux, D.; Amelot, N.; Grat, S.; Brière, C.; Mazars, C.; Pugin, A.; Ranjeva, R. Calcium signaling in plant cell organelles delimited by a double membrane. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Edel, K.H.; Marchadier, E.; Brownlee, C.; Kudla, J.; Hetherington, A.M. The Evolution of Calcium-Based Signalling in Plants. Curr. Biol. 2017, 27, R667–R679. [Google Scholar] [CrossRef] [PubMed]
- Kazmierczak, J.; Kempe, S.; Kremer, B. Calcium in the Early Evolution of Living Systems: A Biohistorical Approach. Curr. Org. Chem. 2013, 17, 1738–1750. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium Signaling. Cell 2014, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Bush, D.S.; Biswas, A.K.; Jones, R.L. Gibberellic-acid-stimulated Ca2+ accumulation in endoplasmic reticulum of barley aleurone: Ca2+ transport and steady-state levels. Planta 1989, 178, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Logan, D.C.; Knight, M.R. Mitochondrial and Cytosolic Calcium Dynamics Are Differentially Regulated in Plants. Plant Physiol. 2003, 133, 21–24. [Google Scholar] [CrossRef]
- Pauly, N.; Knight, M.R.; Thuleau, P.; van der Luit, A.H.; Moreau, M.; Trewavas, A.J.; Ranjeva, R.; Mazars, C. Control of free calcium in plant cell nuclei. Nature 2000, 405, 754. [Google Scholar] [CrossRef]
- Brauer, M.; Zhong, W.-J.; Jelitto, T.; Schobert, C.; Sanders, D.; Komor, E. Free calcium ion concentration in the sieve-tube sap of Ricinus communis L. seedlings. Planta 1998, 206, 103–107. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Kumar, P.; Bae, H. Genomics and evolutionary aspect of calcium signaling event in calmodulin and calmodulin-like proteins in plants. BMC Plant Biol. 2017, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Mohanta, N.; Mohanta, Y.K.; Parida, P.; Bae, H. Genome-wide Identification of Calcineurin B-Like (CBL) Gene Family Of Plants Reveals Novel Conserved Motifs and Evolutionary Aspects In Calcium Signaling Events. BMC Plant Biol. 2015, 15, 189. [Google Scholar] [CrossRef]
- Mohanta, T.; Mohanta, N.; Mohanta, Y.; Bae, H. Genome-Wide Identification of Calcium Dependent Protein Kinase Gene Family in Plant Lineage Shows Presence of Novel D-x-D and D-E-L Motifs in EF-Hand Domain. Front. Plant Sci. 2015, 6, 1146. [Google Scholar] [CrossRef] [PubMed]
- Yamniuk, A.P.; Vogel, H.J. Structural Investigation into the Differential Target Enzyme Regulation Displayed by Plant Calmodulin Isoforms. Biochemistry 2005, 44, 3101–3111. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Kudla, J.; Rodriguez-concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and Calcineurin B –like Proteins: Calcium Sensors for Specific Signal Response Coupling in Plants. Plant Cell 2002, 14, 389–400. [Google Scholar] [CrossRef]
- Lee, S.H.; Johnson, J.D.; Walsh, M.P.; van Lierop, J.E.; Sutherland, C.; Xu, A.; Snedden, W.A.; Kosk-Kosicka, D.; Fromm, H.; Narayanan, N.; Cho, M.J. Differential regulation of Ca2+/calmodulin-dependent enzymes by plant calmodulin isoforms and free Ca2+ concentration. Biochem. J. 2000, 350, 299–306. [Google Scholar] [CrossRef]
- Yoo, J.H.; Park, C.Y.; Kim, J.C.; Do Heo, W.; Cheong, M.S.; Park, H.C.; Kim, M.C.; Moon, B.C.; Choi, M.S.; Kang, Y.H.; et al. Direct Interaction of a Divergent CaM Isoform and the Transcription Factor, MYB2, Enhances Salt Tolerance in Arabidopsis. J. Biol. Chem. 2005, 280, 3697–3706. [Google Scholar] [CrossRef] [PubMed]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Busch, E.; Hohenester, E.; Timpl, R.; Paulsson, M.; Maurer, P. Calcium affinity, cooperativity, and domain interactions of extracellular EF-hands present in BM-40. J. Biol. Chem. 2000, 275, 25508–25515. [Google Scholar] [CrossRef] [PubMed]
- Rupp, B.; Marshak, D.R.; Parkin, S. Crystallization and preliminary X-ray analysis of two new crystal forms of calmodulin. Acta Crystallogr. Sect. D 1996, 52, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, R.; Friedberg, F. Sequence motifs for Calmodulin recognition. FASEB J. 1997, 11, 331–340. [Google Scholar] [CrossRef]
- Hoeflich, K.P.; Ikura, M. Calmodulin in Action: Diversity in Target Recognition and Activation Mechanisms. Cell 2002, 108, 739–742. [Google Scholar] [CrossRef]
- McCormack, E.; Braam, J. Calmodulins and related potential calcium sensors of Arabidopsis. New Phytol. 2003, 159, 585–598. [Google Scholar] [CrossRef]
- Perochon, A.; Aldon, D.; Galaud, J.-P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, R.E. Characterization of three new members of the Arabidopsis thaliana calmodulin gene family: conserved and highly diverged members of the gene family functionally complement a yeast calmodulin null. Planta 2002, 214, 446–455. [Google Scholar] [CrossRef]
- Vanderbeld, B.; Snedden, W.A. Developmental and stimulus-induced expression patterns of Arabidopsis calmodulin-like genes CML37, CML38 and CML39. Plant Mol. Biol. 2007, 64, 683–697. [Google Scholar] [CrossRef]
- Akaboshi, M.; Hashimoto, H.; Ishida, H.; Saijo, S.; Koizumi, N.; Sato, M.; Shimizu, T. The Crystal Structure of Plant-Specific Calcium-Binding Protein AtCBL2 in Complex with the Regulatory Domain of AtCIPK14. J. Mol. Biol. 2008, 377, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Chandran, V.; Stollar, E.J.; Lindorff-Larsen, K.; Harper, J.F.; Chazin, W.J.; Dobson, C.M.; Luisi, B.F.; Christodoulou, J. Structure of the Regulatory Apparatus of a Calcium-dependent Protein Kinase (CDPK): A Novel Mode of Calmodulin-target Recognition. J. Mol. Biol. 2006, 357, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Hrabak, E.; Chan, C.; Gribskov, M. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003, 132, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Campos-Soriano, L.; Gómez-Ariza, J.; Bonfante, P.; San Segundo, B. A rice calcium-dependent protein kinase is expressed in cortical root cells during the presymbiotic phase of the arbuscular mycorrhizal symbiosis. BMC Plant Biol. 2011, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Liu, Y.; Yang, K.-Y.; Han, L.; Mao, G.; Glazebrook, J.; Zhang, S. A fungal-responsive MAPK cascade regulates phytoalexin biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 5638–5643. [Google Scholar] [CrossRef] [PubMed]
- Smotrys, J.E.; Linder, M.E. Palmitoylation of intracellular signaling proteins: Regulation and function. Annu. Rev. Biochem. 2004, 73, 559–587. [Google Scholar] [CrossRef]
- Martín, M.L.; Busconi, L. A rice membrane-bound calcium-dependent protein kinase is activated in response to low temperature. Plant Physiol. 2001, 125, 1442–1449. [Google Scholar] [CrossRef]
- Cheng, S.; Willmann, M.; Chen, H.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef] [PubMed]
- Vaandrager, A.B.; Ehlert, E.M.; Jarchu, T.; Lohmann, S.M.; Jonge, H.R. N-terminal myristoylation is required for membrane localization of cGMP-dependent protein kinase type II. J. Biol. Chem 1996, 271, 7025–7029. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Martin, K.J.; Arthur, J.S.C. CREB phosphorylation at Ser133 regulates transcription via distinct mechanisms downstream of cAMP and MAPK signalling. Biochem. J. 2014, 458, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Astegno, A.; Capitani, G.; Dominici, P. Functional roles of the hexamer organization of plant glutamate decarboxylase. Biochim. Biophys. Acta Proteins Proteomics 2015, 1854, 1229–1237. [Google Scholar] [CrossRef]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Lipid Transfer Proteins As Components of the Plant Innate Immune System: Structure, Functions, and Applications. Acta Naturae 2016, 8, 47–61. [Google Scholar] [PubMed]
- Viola, R.E. The Central Enzymes of the Aspartate Family of Amino Acid Biosynthesis. Acc. Chem. Res. 2001, 34, 339–349. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J.I.; Pysh, L.; Chapple, C. Four Isoforms of Arabidopsis 4-Coumarate: CoA Ligase Have Overlapping yet Distinct Roles in Phenylpropanoid Metabolism. Plant Physiol. 2015, 169, 2409–2421. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Occhipinti, A.; Zebelo, S.; Foti, M.; Fliegmann, J.; Bossi, S.; Maffei, M.E.; Bertea, C.M. Ginkgo biloba responds to herbivory by activating early signaling and direct defenses. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Peiter, E.; Maathuis, F.J.M.; Mills, L.N.; Knight, H.; Pelloux, J.; Hetherington, A.M.; Sanders, D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement. Nature 2005, 434, 404. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Kilpatrick, B.S. Two-pore channels and disease. Biochim. Biophys. Acta. Mol. cell Res. 1865, 1678–1686. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef]
- Park, C.-J.; Seo, Y.-S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Thorneycroft, D.; Schupp, N.; Chapple, A.; Weck, M.; Dunstan, H.; Haldimann, P.; Bechtold, N.; Smith, A.M.; Smith, S.M. Plastidial α-Glucan Phosphorylase Is Not Required for Starch Degradation in Arabidopsis Leaves But Has a Role in the Tolerance of Abiotic Stress. Plant Physiol. 2004, 135, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Post, S.M.; Tomkinson, A.E.; Lee, E.Y.-H.P. The human checkpoint Rad protein Rad17 is chromatin-associated throughout the cell cycle, localizes to DNA replication sites, and interacts with DNA polymerase epsilon. Nucleic Acids Res. 2003, 31, 5568–5575. [Google Scholar] [CrossRef]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 Transcription Factors Are Involved in Brassinosteroid-Regulated Plant Growth and Drought Responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.; Xu, X. Roles of arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biol. 2010, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, S.; Li, Y.; Hua, J. The Arabidopsis BAP1 and BAP2 genes are general inhibitors of programmed cell death. Plant Physiol. 2007, 145, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Parasuraman, B.; Laxmi, A.; Chattopadhyay, D. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants. Plant J. 2009, 58, 778–790. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Q.-Q.; Zhou, L.; Ren, F.; Li, D.-D.; Li, X.-B. Arabidopsis CBL-interacting protein kinase (CIPK6) is involved in plant response to salt/osmotic stress and ABA. Mol. Biol. Rep. 2013, 40, 4759–4767. [Google Scholar] [CrossRef]
- Sardar, A.; Chattopadhyay, D.; Nandi, A.K. CBL-interacting protein kinase 6 negatively regulates immune response to Pseudomonas syringae in Arabidopsis. J. Exp. Bot. 2017, 68, 3573–3584. [Google Scholar] [CrossRef]
- Jin, X.; Sun, T.; Wang, X.; Su, P.; Ma, J.; He, G.; Yang, G. Wheat CBL-interacting protein kinase 25 negatively regulates salt tolerance in transgenic wheat. Sci. Rep. 2016, 6, 28884. [Google Scholar] [CrossRef] [PubMed]
- Bürstenbinder, K.; Rzewuski, G.; Wirtz, M.; Hell, R.; Sauter, M. The role of methionine recycling for ethylene synthesis in Arabidopsis. Plant J. 2007, 49, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Halfter, U.; Ishitani, M.; Zhu, J.-K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. USA 2000, 97, 3735–3740. [Google Scholar] [CrossRef] [PubMed]
- Aghdasi, B.; Ye, K.; Resnick, A.; Huang, A.; Ha, H.C.; Guo, X.; Dawson, T.M.; Dawson, V.L.; Snyder, S.H. FKBP12, the 12-kDa FK506-binding protein, is a physiologic regulator of the cell cycle. Proc. Natl. Acad. Sci. USA 2001, 98, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Gollan, P.J.; Bhave, M.; Aro, E.-M. The FKBP families of higher plants: Exploring the structures and functions of protein interaction specialists. FEBS Lett. 2012, 586, 3539–3547. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.-M.; Chiu, J.; Hsieh, M.-H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.X.; Evans, A.M.; Ma, J.; Parrington, J.; Galione, A. Two-pore channels for integrative Ca signaling. Commun. Integr. Biol. 2010, 3, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Avila, E.L.; Surpin, M.; Brown, M.; Pan, S.; Girke, T.; Raikhel, N.V.; Desikan, R.; Neill, S.J. Expression analysis of Arabidopsis vacuolar sorting receptor 3 reveals a putative function in guard cells. J. Exp. Bot. 2008, 59, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Schulz, P.; Herde, M.; Romeis, T. Calcium-dependent protein kinases: Hubs in plant stress signaling and development. Plant Physiol. 2013, 163, 523–530. [Google Scholar] [CrossRef]
- Boudsocq, M.; Willmann, M.R.; McCormack, M.; Lee, H.; Shan, L.; He, P.; Bush, J.; Cheng, S.-H.; Sheen, J. Differential innate immune signalling via Ca2+ sensor protein kinases. Nature 2010, 464, 418. [Google Scholar] [CrossRef]
- Dubiella, U.; Seybold, H.; Durian, G.; Komander, E.; Lassig, R.; Witte, C.-P.; Schulze, W.X.; Romeis, T. Calcium-dependent protein kinase/NADPH oxidase activation circuit is required for rapid defense signal propagation. Proc. Natl. Acad. Sci. USA 2013, 110, 8744–8749. [Google Scholar] [CrossRef] [PubMed]
- Franz, S.; Ehlert, B.; Liese, A.; Kurth, J.; Cazale, A.-C.; Romies, T. Calcium-dependent protein kinase CPK21 functions in abiotic stress response in Arabidopsis thaliana. Mol. Plant 2015, 4, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Bouschet, T.; Henley, J.M. Calcium as an extracellular signalling molecule: Perspectives on the Calcium Sensing Receptor in the brain. C. R. Biol. 2005, 328, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Krebs, J. Why calcium? How calcium became the best communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef]
- Naraghi, M. T-jump study of calcium binding kinetics of calcium chelators. Cell Calcium 1997, 22, 255–268. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Yoo, B.-C.; Harmon, A.C. Kinetic and Calcium-Binding Properties of Three Calcium-Dependent Protein Kinase Isoenzymes from Soybean. Biochemistry 1998, 37, 6801–6809. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C. Ionic gradients, membrane potential and ionic currents, Chapter 3. In Cellular and Molecular Neurophysiology, 4th ed.; Academic Press: Boston, MA, USA, 2015; pp. 39–54. ISBN 978-0-12-397032-9. [Google Scholar]
- Carafoli, E. The Calcium Pumping ATPase of the Plasma Membrane. Annu. Rev. Physiol. 1991, 53, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Stokes, D.L.; Green, N.M. Structure and Function of the Calcium Pump. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 445–468. [Google Scholar] [CrossRef]
- Brini, M.; Carafoli, E. The Plasma Membrane Ca2+ ATPase and the Plasma Membrane Sodium Calcium Exchanger Cooperate in the Regulation of Cell Calcium. Cold Spring Harb. Perspect. Biol. 2011, 3, a004168. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, B.; Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Celsi, F.; Pizzo, P.; Brini, M.; Leo, S.; Fotino, C.; Pinton, P.; Rizzuto, R. Mitochondria, calcium and cell death: A deadly triad in neurodegeneration. Biochim. Biophys. Acta 2009, 1787, 335–344. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef] [PubMed]
- Dubois, C.; Prevarskaya, N.; Vanden Abeele, F. The calcium-signaling toolkit: Updates needed. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Edel, K.H.; Kudla, J. Increasing complexity and versatility: How the calcium signaling toolkit was shaped during plant land colonization. Cell Calcium 2015, 57, 231–246. [Google Scholar] [CrossRef]
- Marchadier, E.; Oates, M.E.; Fang, H.; Donoghue, P.C.J.; Hetherington, A.M.; Gough, J. Evolution of the Calcium-Based Intracellular Signaling System. Genome Biol. Evol. 2016, 8, 2118–2132. [Google Scholar] [CrossRef] [PubMed]
- Contreras, L.; Drago, I.; Zampese, E.; Pozzan, T. Mitochondria: The calcium connection. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 607–618. [Google Scholar] [CrossRef]
- Duchen, M.R. Mitochondria and calcium: From cell signalling to cell death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef]
- Schönknecht, G. Calcium Signals from the Vacuole. Plants 2013, 2, 589–614. [Google Scholar] [CrossRef] [PubMed]
- Sello, S.; Moscatiello, R.; Mehlmer, N.; Leonardelli, M.; Carraretto, L.; Cortese, E.; Zanella, F.G.; Baldan, B.; Szabò, I.; Vothknecht, U.C.; Navazio, L. Chloroplast Ca2+ fluxes into and across thylakoids revealed by thylakoid-targeted aequorin probes. Plant Physiol. 2018, 177, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Peiter, E. The plant vacuole: Emitter and receiver of calcium signals. Cell Calcium 2011, 50, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.-P.; Thuleau, P.; Mazars, C. Calcium Sensors as Key Hubs in Plant Responses to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Name of the Species | Taxonomy | Genome Size | Total No. of Protein Coding Genes | No. of CaMs | No. of CMLs | No. of CPKs | No. of CBLs |
|---|---|---|---|---|---|---|---|---|
| 1 | Aquilegia coerulea | Dicot | 306.5 | 30023 | 5 | 21 | 16 | 5 |
| 2 | Arabidopsis thaliana | Dicot | 135 | 27416 | 9 | 47 | 34 | 10 |
| 3 | Brachypodium distachyon | Monocot | 272 | 34310 | 5 | 23 | 27 | 9 |
| 4 | Brassica rapa | Dicot | 283.8 | 40492 | 13 | 36 | 49 | 14 |
| 5 | Capsella rubella | Dicot | 134.8 | 26521 | 10 | 29 | 32 | 9 |
| 6 | Carica papaya | Dicot | 135 | 27332 | 5 | 15 | 15 | 4 |
| 7 | Chlamydomonas reinhardtii | Algae | 111.1 | 17741 | 6 | 3 | 14 | 2 |
| 8 | Citrus clementina | Dicot | 301.4 | 24533 | 8 | 19 | 26 | 7 |
| 9 | Citrus sinensis | Dicot | 319 | 25376 | 6 | 20 | 24 | 8 |
| 10 | Coccomyxa subellipsoidea | Algae | 49 | 9629 | 3 | 2 | 2 | 0 |
| 11 | Cucumis sativus | Dicot | 203 | 21491 | 6 | 21 | 18 | 7 |
| 12 | Eucalyptus grandis | Dicot | 691 | 36349 | 1 | 25 | 22 | 12 |
| 13 | Fragaria vesca | Dicot | 240 | 32831 | 5 | 19 | 14 | 6 |
| 14 | Glycine max | Dicot | 978 | 56044 | 6 | 27 | 41 | 9 |
| 15 | Gossypium raimondii | Dicot | 761.4 | 55294 | 6 | 30 | 40 | 13 |
| 16 | Linum usitatissimum | Dicot | 318.3 | 43471 | 11 | 21 | 47 | 12 |
| 17 | Malus domestica | Dicot | 881.3 | 63514 | 9 | 32 | 28 | 11 |
| 18 | Manihot esculenta | Dicot | 532.5 | 33033 | 9 | 22 | 26 | 9 |
| 19 | Medicago truncatula | Dicot | 360 | 50894 | 4 | 24 | 11 | 11 |
| 20 | Micromonas pusilla | Algae | 22 | 10660 | 5 | 8 | 2 | 3 |
| 21 | Mimulus guttatus | Dicot | 321.7 | 28140 | 13 | 19 | 25 | 9 |
| 22 | Oryza sativa | Monocot | 372 | 42189 | 5 | 33 | 30 | 11 |
| 23 | Ostreococcus lucimarinus | Algae | 13.2 | 7796 | 2 | 2 | 3 | 0 |
| 24 | Panicum virgatum | Monocot | 1358 | 102065 | 9 | 20 | 53 | 10 |
| 25 | Phaseolus vulgaris | Dicot | 537.2 | 27433 | 9 | 26 | 25 | 10 |
| 26 | Physcomitrella patens | Bryophyte | 480 | 32926 | 7 | 17 | 25 | 4 |
| 27 | Picea abies | Pinophyta | 1960 | 28354 | 9 | 15 | 11 | 13 |
| 28 | Populus trichocarpa | Dicot | 422.9 | 42950 | 8 | 26 | 28 | 11 |
| 29 | Prunus persica | Dicot | 225.7 | 26873 | 4 | 21 | 17 | 7 |
| 30 | Ricinus communis | Dicot | 400 | 31221 | 4 | 8 | 15 | 8 |
| 31 | Selaginella moellendorffii | Pteridophyte | 212.5 | 22273 | 6 | 11 | 9/11 | 4 |
| 32 | Setaria italica | Monocot | 405.7 | 34584 | 5 | 17 | 27 | 7 |
| 33 | Solanum lycopersicum | Dicot | 900 | 34727 | 9 | 27 | 28 | 11 |
| 34 | Solanum tuberosum | Dicot | 723 | 39028 | 5 | 27 | 21 | 12 |
| 35 | Sorghum bicolor | Monocot | 693.9 | 35490 | 8 | 22 | 28 | 8 |
| 36 | Thellungiella halophila | Dicot | 238.5 | 26351 | 10 | 27 | 31 | 9 |
| 37 | Theobroma cacao | Dicot | 346 | 29452 | 2 | 14 | 17 | 7 |
| 38 | Vitis vinifera | Dicot | 487 | 26346 | 5 | 13 | 17 | 9 |
| 39 | Volvox carteri | Algae | 131.2 | 14247 | 4 | 4 | 6 | 0 |
| 40 | Zea mays | Monocot | 2400 | 63540 | 8 | 21 | 47 | 9 |
| EF-Hands | CaMs | CMLs | CBLs | CPKs |
|---|---|---|---|---|
| 1st | D-x-D, E-x2-E | D-x3-D, F-x2-F | V-F-H-P-N | D-x-D, D-E-E-L, E-E-I, E-M-F |
| 2nd | D-x-D, D-F-x-E-F | D-x3-D | D & E | D-x-D, D-E-E-L, E-x-E |
| 3rd | D-x-D | D & E | E-E-x-D, D-D-x2-E | D-x-D, D-E-E-L, x-E-D |
| 4th | D-x-D | D-x-D-x-D, F-x-E-F | D-x-D, D-E-E-L, D/E |
| Calcium Binding Proteins | Myristoylation Sites | Palmitoylation Sites |
|---|---|---|
| CaM | Not present | Not present |
| CML | M-G-F, M-G-G, M-G-A | Not present |
| CBL | M-G-C | M-G-C, M-L-Q-C |
| CPK | MGN, MGC, QFG, MGL, MGS, MGI, MGQ, MGV, | M-G-N-C, M-G-N-C-C, M-G-C, M-G-N-T-C-V, Q-F-G-T-T-Y-L-C, M-G-N-C-C-R, M-G-L-C, M-G-G-C, M-G-N-N-C, M-G-S-C, M-G-N-S-C, Q-F-G-T-T-F-L-C, M-G-I-C, M-G-N-C-N-A-C, M-G-Q-C, M-G-N-A-C, Q-F-G-T-T-Y-Q-C, M-G-N-V-C, M-G-V-C, M-G-N-Q-C, Q-F-G-I-T-Y-L-C, M-G-N-C-N-T-C, M-E-L-C |
| Gene Name | Interacting Proteins |
|---|---|
| CaM | |
| CaM8 | APRR2, TPC1, At2g01210, NOA1, At2g20050, At1g73440, CUTA, GRIK2, GRIK1, CAMS1 |
| CaM1 | ZW1, CDPK1, NTL9, At2g01210, DSPTP1, CRCK, GAD, At5g62570, At5g28300, AS1 |
| CaM2 | UBP6, ZWI, At3g07670, At5g14260, CIP111, At1g24610, At5g53920, IQD20, SD1-13 |
| CaM3 | PP7, ZWI, UBP6, MPK8, TPC1, CRK1, SD1-13, AGL24, At3g07670, At5g14260 |
| CaM5 | MPK8, At2g19750, TPC1, EDA39, CRK1, NOA1, PP7, CRLK1, At2g02970 |
| CaM4 | HOT5, CNBT1, At4g33080, ACA11, TPC1, PP7, MPK8, At2g01820, LP1, CaM1 |
| CaM7 | At5g57110, MPK8, HY5, CNBT1, BI1, TPC1, At2g18750, NOA1, TSO1-like |
| CaM8 | APRR2, TPC1, At2g01210, NOA1, At2g20050, At1g73440, CUTA, GRIK2, GRIK1, CAMS1 |
| CaM9 | APRR2, At1g73440, TPC1, ATG8H, APG8A, At2g27480, TSO1-like |
| CML | |
| CML15: AT1G18530 | ACOS5, TPC1, LAP6, At2g27480, LAP5, GDSL, TKPR1, NHX1, CYP704B1 |
| CML25: AT1G24620 | CCP3, HSP81-3, ARP2, HSP81-4, DEK1, PHS2, LOS1, PGSIP4, HSP90.1 |
| CML14: AT1G62820 | CML43, CML42, TPC1, LRR, ZWI, NOA1, BIP2 |
| CML23: AT1G66400 | TCH2, MSS3, ARP2, DEK1, PHS2, LOS1, HSP81.3, HSP81.4, HSP90.1 |
| CML39: AT1G76640 | At1g54850, IDL3, TPC1, NOA1 |
| CML38: AT1G76650 | WRKY40, SZF1, STZ, BAP1, TPC1, RHL41, |
| CML30: AT2G15680 | ARP2, DEK1, PHS2, LOS1, HSP81, ROC1 |
| CML10: AT2G41090 | At3g19100, CPK30, CDPK6, CPK31, CPK29, CPK16, CPK18, CPK4, CPK28, CPK15 |
| CML5: AT2G43290 | TCH2, CML23, ARP2, DEK1, PHS2, LOS1, At2g41410, At1g07940 |
| CML3: AT3G07490 | HSP70, ARP2, DEK1, PHS2, LOS1, PGSIP4, HSP81 |
| CML36: AT3G10190 | At1g62820, ARP2, DEK1, LOS1, HSP81, HSP90, CML30 |
| CML16: AT3G25600 | At4g27280, At3g10300, TPC1, At4g26470, SYP122, GILP, NHL3 |
| CML20: AT3G50360 | RAD4, RAD23B, RAD23A, RAD23D, RAD23C, DDB1B, DDB1A, GTF2H2, SAC3B |
| CML41: AT3G50770 | At5g43260, TPC1, BGLU28, TI1, MDHAR, BT2, NOA1, ST2A |
| CML9: AT3G51920 | APRR2, TPC1, ATG8H, APG8A, CDPK19, |
| CML4: AT3G59440 | F5M15.5, ARP2, DEK1, PHS2, LOS1, HSP81, HSP90, ROC1 |
| CML42: AT4G20780 | TPC1, PPI3A, CYP86A |
| CML19: AT4G37010 | RAD4, RAD23B, RAD23A, RAD23D, RAD23C, DDB2, SAC3B, At5g16090, CUL4, GTF2H2 |
| CML37: AT5G42380 | WRKY46, RHL41, BAP1, WRKY40, TPC1, GILP |
| CML43: AT5G44460 | TPC1, CYP86A1, WRKY27, NOA1, CIPK26, CIPK3 |
| CBL | |
| CBL1 | CIPK23, AKT1, SOS2, CIPK1, CIPK7, At2g20050, CIPK15, CIPK8, CIPK26 |
| CBL2 | CIPK14, SIP4, At2g20050, CIPK23, At3g51390, SOS2, SIP3, CIPK12, CIPK18, MTN1 |
| CBL3 | CIPK23, At2g20050, SIP3, CIPK9, MTN1, CIPK1, MTN2, At3g51390, SOS2, CIPK14 |
| CBL4 | SOS2, SOS1, SIP3, NHX1, AKT1, At2g20050, At1g61575, CIPK8, CIPK14 |
| CBL5 | At2g20050, FKBP12, AKT1, CIPK1, CIPK23, SOS2, SIP4, CIPK14, TPC1, At3g59440 |
| CBL6 | VSR2, MTN1, FKBP12, CIPK9, CIPK1, TPC1, At4g10170 |
| CBL7 | GLR2.8, SOS2, CIPK3, CIPK26, CIPK9, FIBP12 |
| CBL8 | CIPK23, FKBP12, CIPK14, HAK5, TPC1, CIPK9, AKT1, SOS2, CIPK12 |
| CBL9 | CIPK23, AKT1, CIPK1, SIP3, CIPK8, SOS2, CIPK9, CIPK3, CIPK14 |
| CBL10 | SOS2, AKT1, CIPK23, SIP3, SOS1, FKBP12, CIPK8, TPC1, AVP1 |
| CPK | |
| At5g24430 | CPK1, At2g45300, CDPK1, CPK6, CPK9, At2g41090, CPK19, CPK24, CPK28, CPK32 |
| CDPK1 | CPK1, CPK4, CPK5, CPK6, CPK9, CPK28, CPK29, At5g34430, CPK32, At2g41090 |
| CPK30 | At2g41090, CRK1, CPK6, CPK9, CPK16, CPK18, CPK28, CDPK2, At5g24430, CPK23 |
| CPK9 | At2g41090, ABI2, CRK1, CPK4, At3g49370, At5g24430, CPK23, CPK28, CPK31, CPK32 |
| CPK19 | At2g41090, At5g24430, CRK1, CPK5, CPK9, CPK16, CPK21, CPK28, F5M15.5, |
| CPK7 | CPK1, CPK4, CPK6, CPK9, CPK16, CPK18, CPK28, CPK29, CDPK1, At2g41090 |
| CPK1 | CDPK1, CDPK6, CPK28, CPK6, CPK4, CPK9, At2g41090, CPK32, CPK7, At5g24430 |
| CPK18 | At2g41090, CPK14, CPK19, CPK7, CDPK1, CPK5, CPK13, CPK32, CPK24, CPK30 |
| CDPK6 | At2g41090, ORP2A, CPK1, CPK9, CPK6, CDPK1, OZS1, CPK28, CPK18, CPK16 |
| CPK15 | At2g41090, CPK18, CPK16, CPK28, CPK24, CPK14, At3g49370, At5g24430, CPK30, CPK7 |
| CPK4 | ABF1, CPK28, CPK29, At2g41090, CPK1, CPK32, CPK6, CPK9, RBOHD |
| CPK31 | At2g41090, CPK27, CRK1, At3g49370, CPK16, CPK18, CPK32, CPK25, CPK14, CPK13 |
| CPK32 | CPK28, ABF4, CPK6, CPK4, CDPK1, CPK9, CPK1, At2g41090, CPK9, CPK16 |
| CPK2 | At2g41090, CPK16, CPK19, CPK18, CPK5, At3g49370, At5g24430, CRK1, CPK1, At3g56760 |
| CPK14 | CPK24, CPK16, At2g41090, CPK20, CPK28, CPK18, CPK31, CPK29, CPK15, CPK19 |
| CPK6 | OZS1, CDPK1, CPK9, CPK28, CPK32, At5g24430, CPK29, CPK1, CPK4, CPK5 |
| Reactome Pathway | Functional Pathway | Pfam Pathway | GO Component |
|---|---|---|---|
| CML | |||
| DNA Repair | Calcium ion binding | EF-hand domain | GO:0005623→cell |
| DNA Damage Recognition in GG-NER | Binding | EF-hand domain pair | GO:0005622→intracellular |
| Formation of Incision Complex in GG-NER | Metal ion binding | EF-hand domain pair | GO:0044464→cell part |
| Nucleotide Excision Repair | Ion binding | EF hand | GO:0044424→intracellular part |
| Calmodulin induced events | Calmodulin-dependent protein kinase activity | EF hand | GO:0005634→nucleus |
| Ca-dependent events | Calcium-dependent protein serine/threonine kinase activity | XPC-binding domain | GO:0043229→intracellular organelle |
| CaM pathway | Damaged DNA binding | WD domain, G-beta repeat | GO:0043227→membrane-bounded organelle |
| DAG and IP3 signaling | Calmodulin binding | UBA/TS-N domain | GO:0005737→cytoplasm |
| Activation of CaMK IV | Protein binding | WRKY DNA -binding domain | GO:0043231→intracellular membrane-bounded organelle |
| Opioid Signaling | Hsp90 protein binding | TPR repeat | GO:0080008→Cul4-RING E3 ubiquitin ligase complex |
| G-protein mediated events | Organic cyclic compound binding | Kinase-like | GO:0031461→cullin-RING ubiquitin ligase complex |
| PLC beta mediated events | Heterocyclic compound binding | Tetratricopeptide repeat | GO:1990234→transferase complex |
| Intracellular signaling by second messengers | Proteasome binding | Sulfotransferase family | GO:0005829→cytosol |
| G2/M DNA damage checkpoint | Polyubiquitin modification-dependent protein binding | Ubiquitin family | GO:0005886→plasma membrane |
| G alpha (i) signaling events | Preprotein binding | Hsp90 protein | GO:0044444→cytoplasmic part |
| Signaling by GPCR | Ubiquitin binding | CPSF A subunit region | GO:0071944→cell periphery |
| GPCR downstream signaling | Protein serine/threonine kinase activity | 3-Oxoacyl-[acyl-carrier-protein (ACP)] synthase III | GO:0005773→vacuole |
| Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks | Tetraketide alpha-pyrone synthase activity | Mono-functional DNA-alkylating methyl methanesulfonate N-term | GO:0016020→membrane |
| Neutrophil degranulation | Nucleic acid binding | Secreted protein acidic and rich in cysteine Ca binding region | GO:0032991→protein-containing complex |
| Immune System | Protein-containing complex binding | Tetratricopeptide repeat | GO:0034399→nuclear periphery |
| Post-translational protein modification | DNA binding | Protein kinase domain | GO:0098805→whole membrane |
| Josephin domain DUBs | Anion binding | Protein tyrosine kinase | GO:0005777→peroxisome |
| Metabolism of proteins | Purine ribonucleotide triphosphate binding | Chalcone and stilbene synthases, N-terminal domain | GO:0012505→endomembrane system |
| Dual Incision in GG-NER | Purine ribonucleotide binding | Tetratricopeptide repeat | |
| Signal Transduction | Serine-type endopeptidase inhibitor activity | Histidine kinase-, DNA gyrase B- and HSP90-like ATPase | |
| Neddylation | Drug binding | Sulfotransferase domain | |
| Formation of TC-NER Pre-Incision Complex | Small molecule binding | Tetratricopeptide repeat | |
| Dual incision in TC-NER | Nucleotide binding | Chalcone and stilbene synthases, C-terminal domain | |
| Gap-filling DNA repair synthesis and ligation in TC-NER | ATP binding | NAF domain | |
| HSP90 chaperone cycle for steroid hormone receptors (SHR) | Sulfotransferase activity | Histidine kinase-, DNA gyrase B- and HSP90-like ATPase | |
| N-glycan trimming in the ER and Calnexin/Calreticulin cycle | Transferase activity | Ubiquitin-2 like Rad60 SUMO-like | |
| Recognition of DNA damage by PCNA-containing replication complex | Enzyme binding | Lipopolysaccharide kinase (Kdo/WaaP) family | |
| Cellular responses to external stimuli | Sequence-specific DNA binding | C2H2-type zinc finger | |
| Cellular responses to stress | Transcription regulator activity | ||
| Glycogen breakdown (glycogenolysis) | Catalytic activity, acting on a protein | ||
| Protein methylation | Ras GTPase binding | ||
| Cytosolic sulfonation of small molecules | |||
| Asparagine N-linked glycosylation | |||
| CPK | |||
| Calmodulin induced events | Calmodulin-dependent protein kinase activity | EF-hand domain pair | GO:0005634 nucleus |
| Ca-dependent events | Calcium-dependent protein serine/threonine kinase activity | EF-hand domain pair | GO:0005886 Plasma membrane |
| CaM pathway | Calmodulin binding | EF hand | GO:0016020 membrane |
| DAG and IP3 signaling | Calcium ion binding | EF-hand domain | GO:0043231 Intracellular membrane bound organelle |
| Activation of CaMK IV | Protein binding | EF hand | GO:0005737 cytoplasm |
| G2/M DNA damage checkpoint | ATP binding | Protein kinase domain | GO:0071944 cell periphery |
| Opioid Signaling | Metal ion binding | Protein tyrosine kinase | GO:0005622 intracellular |
| G-protein mediated events | Heterocyclic compound binding | Lipopolysaccharide kinase (Kdo/WaaP) family | GO:0005623 cell |
| PLC beta mediated events | Organic cyclic compound binding | Secreted protein acidic and rich in cysteine Ca binding region | GO:0044464 cell part |
| Intracellular signaling by second messengers | Binding | Kinase-like | GO:0005829 cytosol |
| G alpha (i) signaling events | Catalytic activity | bZIP transcription factor | |
| Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks | Protein phosphatase binding | Basic region leucine zipper | |
| Signaling by GPCR | Peroxidase activity | ||
| GPCR downstream signaling | |||
| Signal Transduction | |||
| CaM | |||
| Neurotransmitter receptors and postsynaptic signal transmission | Calmodulin binding | Cytoskeletal-regulatory complex EF hand | GO:0005622 Intracellular |
| Post NMDA receptor activation events | Calcium ion binding | EF hand | GO:0005623 Cell |
| CREB phosphorylation through the activation of CaMKK | Binding | EF-hand domain pair | GO:0044464 Cell Part |
| Activation of NMDA receptors and postsynaptic events | Protein binding | EF-hand domain pair | GO:0044424 Intracellular part |
| Transmission across Chemical Synapses | Ion binding | EF hand | GO:0043229 Intracellular organelle |
| Neuronal System | Metal ion binding | EF-hand domain | GO:0005737 Cytoplasm |
| Ion channel transport | Protein serine/threonine kinase activity | EF-hand domain | GO:0043227 Membrane bound organelle |
| Opioid Signaling | Purine ribonucleotide binding | Lipopolysaccharide kinase (Kdo/WaaP) family | GO:0005874 Microtubule |
| Calmodulin induced events | Adenyl ribonucleotide binding | Autophagy protein Atg8 ubiquitin like | GO:0015630 Microtubule cytoskeleton |
| Ca-dependent events | Small molecule binding | Ubiquitin-like autophagy protein Apg12 | GO:0043231 Intracellular membrane bound organelle |
| CaM pathway | Anion binding | Protein kinase domain | GO:0005634 Nucleus |
| G-protein mediated events | Drug binding | Protein tyrosine kinase | GO:0044446 Intracellular organelle part |
| PLC beta mediated events | Purine ribonucleoside triphosphate binding | Ion transport protein | GO:0000421 Autophagosome membrane |
| DAG and IP3 signaling | Organic cyclic compound binding | Kinase-like | GO:0005886 Plasma membrane |
| Signal Transduction | Heterocyclic compound binding | GO:0071944 Cell periphery | |
| Activation of CaMK IV | ATP binding | GO:0005776 Autophagosome | |
| G2/M DNA damage checkpoint | Catalytic activity, acting on a protein | GO:0043232 Intracellular non-membrane bound organelle | |
| Intracellular signaling by second messengers | Voltage-gated cation channel activity | GO:0016020 Membrane | |
| G alpha (i) signaling events | Calmodulin-dependent protein kinase activity | GO:0098805 Whole membrane | |
| Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks | Calcium-dependent protein serine/threonine kinase activity | GO:0005774 Vacuolar membrane | |
| Signaling by GPCR | Transcription regulatory region DNA binding | GO:0005773 Vauole | |
| GPCR downstream signaling | Calcium ion transmembrane transporter activity | ||
| Metal ion transmembrane transporter activity | |||
| Identical protein binding | |||
| RNA polymerase II regulatory region sequence-specific DNA binding | |||
| CBL | |||
| NA | Calcium ion binding | NAF domain | GO:0005623 Cell |
| Protein serine/threonine kinase activity | Kinase-like | GO:0044464 Cell part | |
| Ion binding | EF hand | GO:0005886 Plasma membrane | |
| Binding | Protein kinase domain | GO:0005622 Intracellular | |
| Catalytic activity, acting on a protein | Protein tyrosine kinase | GO:0000325 Plant-type vacuole | |
| Drug binding | EF-hand domain pair | GO:0071944 Cell periphery | |
| ATP binding | EF-hand domain pair | GO:0005737 Cytoplasm | |
| Metal ion transmembrane transporter activity | Phosphorylase superfamily | GO:0005774 Vacuolar membrane | |
| Potassium ion transmembrane transporter activity | Ion transport protein | GO:0016020 Membrane | |
| Cation channel activity | EF hand | GO:0005773 Vacuole | |
| Adenosylhomocysteine nucleosidase activity | GO:0009705 Plant-type vacuole membrane | ||
| Methylthioadenosine nucleosidase activity | GO:0043231 Intracellular membrane bound organelle | ||
| Metal ion binding | GO:0005634 Nucleus | ||
| Ion gated channel activity | GO:0044444 Cytoplasmic part | ||
| Sodium ion transmembrane transporter activity | |||
| Catalytic activity | |||
| Transmembrane transporter activity | |||
| Ligand-gated ion channel activity | |||
| Voltage-gated cation channel activity | |||
| Calcium channel activity | |||
| Potassium channel activity | |||
| Organic cyclic compound binding | |||
| Heterocyclic compound binding | |||
| Active transmembrane transporter activity | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohanta, T.K.; Yadav, D.; Khan, A.L.; Hashem, A.; Abd_Allah, E.F.; Al-Harrasi, A. Molecular Players of EF-hand Containing Calcium Signaling Event in Plants. Int. J. Mol. Sci. 2019, 20, 1476. https://doi.org/10.3390/ijms20061476
Mohanta TK, Yadav D, Khan AL, Hashem A, Abd_Allah EF, Al-Harrasi A. Molecular Players of EF-hand Containing Calcium Signaling Event in Plants. International Journal of Molecular Sciences. 2019; 20(6):1476. https://doi.org/10.3390/ijms20061476
Chicago/Turabian StyleMohanta, Tapan Kumar, Dhananjay Yadav, Abdul Latif Khan, Abeer Hashem, Elsayed Fathi Abd_Allah, and Ahmed Al-Harrasi. 2019. "Molecular Players of EF-hand Containing Calcium Signaling Event in Plants" International Journal of Molecular Sciences 20, no. 6: 1476. https://doi.org/10.3390/ijms20061476
APA StyleMohanta, T. K., Yadav, D., Khan, A. L., Hashem, A., Abd_Allah, E. F., & Al-Harrasi, A. (2019). Molecular Players of EF-hand Containing Calcium Signaling Event in Plants. International Journal of Molecular Sciences, 20(6), 1476. https://doi.org/10.3390/ijms20061476