Efficient and Stable Delivery of Multiple Genes to Fish Cells by a Modified Recombinant Baculovirus System


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Construction of Recombinant Baculovirus Containing Dual-Shuttle Promoters Following a pr/pf Cassette
2.2. Fast and Precise Titer Determination Based on Fluorescence Assay
2.3. Efficiently Stable Gene Delivery into Fish Cells by Recombinant Baculovirus
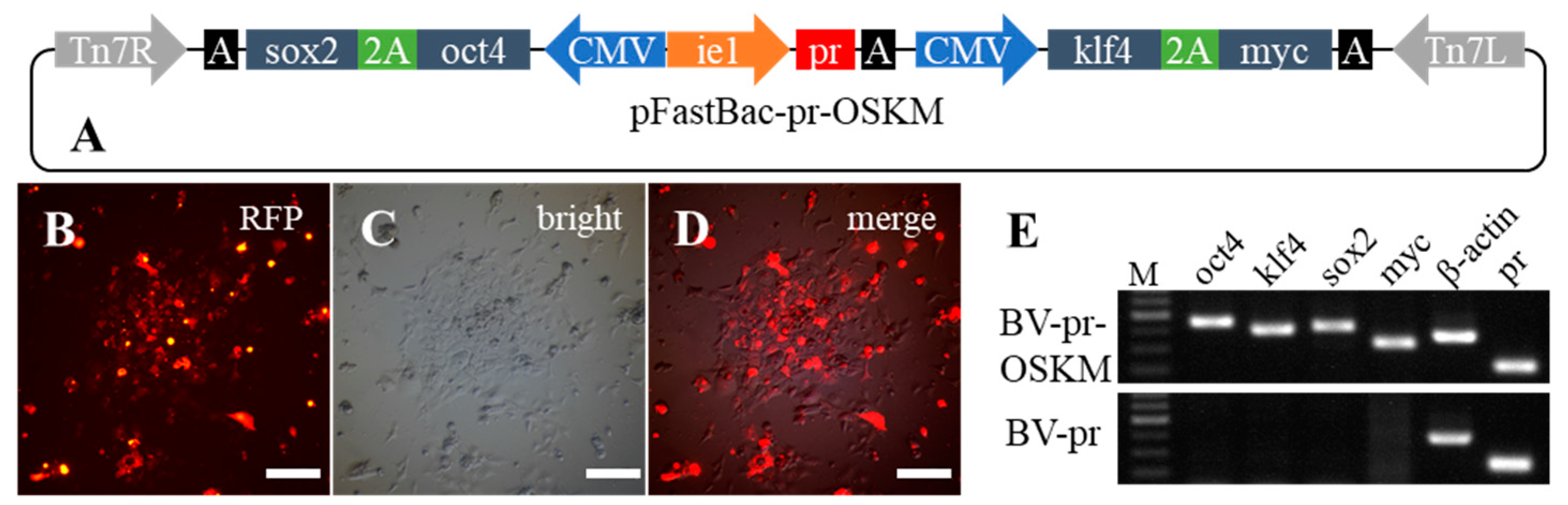
2.4. The Capacity of Stable Delivery of Multiple Genes into Fish Cells by Recombinant Baculovirus
2.5. The Robust Promoter Activity of WSSV ie1 in Fish Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid and Recombinant Baculoviral Vectors
4.3. Titer Determination
4.4. Transduction of Fish Cells
4.5. Fluorescence-Positive Cell Counting
4.6. Western Blot
4.7. Colony Formation Assay
4.8. PCR Assay
4.9. Microinjection of Medaka Embryos
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Luckow, V.A.; Summers, M.D. Trends in the development of baculovirus expression vectors. Nat. Biotechnol. 1988, 6, 47–55. [Google Scholar] [CrossRef]
- Slepushkin, V.A.; Katz, J.M.; Black, R.A.; Gamble, W.C.; Rota, P.A.; Cox, N.J. Protection of mice against influenza A virus challenge by vaccination with baculovirus-expressed M2 protein. Vaccine 1995, 13, 1399–1402. [Google Scholar] [CrossRef]
- Hofmann, C.; Sandig, V.; Jennings, G.; Rudolph, M.; Schlag, P.; Strauss, M. Efficient gene transfer into human hepatocytes by baculovirus vectors. Proc. Natl. Acad. Sci. USA 1995, 92, 10099–10103. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lin, C.Y.; Chen, G.Y.; Hu, Y.C. Baculovirus as a gene delivery vector: Recent understandings of molecular alterations in transduced cells and latest applications. Biotechnol. Adv. 2011, 29, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Du, J.; Chen, T.; Yi, M.; Li, M.; Wang, S.; Li, C.M.; Hong, Y. Establishment of medakafish as a model for stem cell-based gene therapy: Efficient gene delivery and potential chromosomal integration by baculoviral vectors. Exp. Cell Res. 2009, 315, 2322–2331. [Google Scholar] [CrossRef] [PubMed]
- Wagle, M.; Jesuthasan, S. Baculovirus-mediated gene expression in zebrafish. Mar. Biotechnol. 2003, 5, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Leisy, D.J.; Lewis, T.D.; Leong, J.A.; Rohrmann, G.F. Transduction of cultured fish cells with recombinant baculoviruses. J. Gen. Virol. 2003, 84, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Cao, S.; Cui, X.; Xiong, C.; Wang, M.; Lu, Y.; Wang, W.; Ye, J.; Liu, X. Efficient gene delivery into fish cells by an improved recombinant baculovirus. J. Virol. Methods 2011, 173, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wang, Y.; Li, N.; Peng, W.P.; Sun, Y.; Tong, G.Z.; Qiu, H.J. Efficient gene delivery into mammalian cells mediated by a recombinant baculovirus containing a whispovirus ie1 promoter, a novel shuttle promoter between insect cells and mammalian cells. J. Biotechnol. 2007, 131, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Knebel, D.; Doerfler, W. Activation of an insect baculovirus promoter in mammalian cells by adenovirus functions. Virus Res. 1987, 8, 317–326. [Google Scholar] [CrossRef]
- Ping, W.; Ge, J.; Li, S.; Zhou, H.; Wang, K.; Feng, Y.; Lou, Z. Baculovirus-mediated gene expression in chicken primary cells. Avian. Dis. 2006, 50, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xiang, J.; Zhou, G.; Ron, T.B.; Tong, H.I.; Kang, W.; Sun, S.; Lu, Y. The Pacific White Shrimp beta-actin Promoter: Functional Properties and the Potential Application for Transduction System Using Recombinant Baculovirus. Mar. Biotechnol. 2016, 18, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Takata, Y.; Kishine, H.; Sone, T.; Andoh, T.; Nozaki, M.; Poderycki, M.; Chesnut, J.D.; Imamoto, F. Generation of iPS Cells Using a BacMam Multigene Expression System. Cell Struct. Funct. 2011, 36, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Robles, V.; Marti, M.; Izpisua Belmonte, J.C. Study of pluripotency markers in zebrafish embryos and transient embryonic stem cell cultures. Zebrafish 2011, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Moreland, R.B.; Langevin, G.L.; Singer, R.H.; Garcea, R.L.; Hereford, L.M. Amino acid sequences that determine the nuclear localization of yeast histone 2B. Mol. Cell Biol. 1987, 7, 4048–4057. [Google Scholar] [CrossRef] [PubMed]
- Romoren, K.; Thu, B.J.; Bols, N.C.; Evensen, O. Transfection efficiency and cytotoxicity of cationic liposomes in salmonid cell lines of hepatocyte and macrophage origin. Biochim. Biophys. Acta. 2004, 1663, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Encinas, P.; Carbajosa, S.; Cuesta, A.; Chaves-Pozo, E.; Tafalla, C.; Estepa, A.; Coll, J.M. Transfection improvements of fish cell lines by using deacylated polyethylenimine of selected molecular weights. Fish Shellfish Immunol. 2009, 26, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Hink, W.F.; Vail, P.V. Plaque Assay for Titration of Alfalfa Looper Nuclear Polyhedrosis-Virus in a Cabbage-Looper (Tn-368) Cell Line. J. Invertebr. Pathol. 1973, 22, 168–174. [Google Scholar] [CrossRef]
- Shen, C.F.; Meghrous, J.; Kamen, A. Quantitation of baculovirus particles by flow cytometry. J. Virol. Methods 2002, 105, 321–330. [Google Scholar] [CrossRef]
- Kitts, P.A.; Green, G. An immunological assay for determination of baculovirus titers in 48 hours. Anal. Biochem. 1999, 268, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Roldao, A.; Oliveira, R.; Carrondo, M.J.; Alves, P.M. Error assessment in recombinant baculovirus titration: Evaluation of different methods. J. Virol. Methods 2009, 159, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Hitchman, R.B.; Siaterli, E.A.; Nixon, C.P.; King, L.A. Quantitative real-time PCR for rapid and accurate titration of recombinant baculovirus particles. Biotechnol. Bioeng. 2007, 96, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Janakiraman, V.; Forrest, W.F.; Chow, B.; Seshagiri, S. A rapid method for estimation of baculovirus titer based on viable cell size. J. Virol. Methods 2006, 132, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Liu, T.; Pan, J.; Miao, P.; Zhang, C. Rapid baculovirus titration assay based on viable cell side scatter (SSC). Anal. Chim. Acta. 2015, 879, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; Gotoh, T.; Bentley, W.E. Simplification of titer determination for recombinant baculovirus by green fluorescent protein marker. Biotechniques 1997, 23, 782–784. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, R.; Esposito, D. A rapid method for titrating baculovirus stocks using the Sf-9 Easy Titer cell line. Biotechniques 2009, 47, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Wu, H.H.; Chen, C.P.; Chern, S.R.; Hwang, S.M.; Huang, S.F.; Lo, W.H.; Chen, G.Y.; Hu, Y.C. Biosafety assessment of human mesenchymal stem cells engineered by hybrid baculovirus vectors. Mol. Pharm. 2011, 8, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Cheshenko, N.; Krougliak, N.; Eisensmith, R.; Krougliak, V. A novel system for the production of fully deleted adenovirus vectors that does not require helper adenovirus. Gene Ther. 2001, 8, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, J.P.; Land, H. Advanced mammalian gene transfer: High titre retroviral vectors with multiple drug selection markers and a complementary helper-free packaging cell line. Nucleic Acids Res. 1990, 18, 3587–3596. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Shima, A.; Kawakami, N. Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage. Proc. Natl. Acad. Sci. USA 2000, 97, 11403–11408. [Google Scholar] [CrossRef] [PubMed]
- Ivics, Z.; Hackett, P.; Plasterk, R.; Izsvák, Z. Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 1997, 91, 501–510. [Google Scholar] [CrossRef]
- He, F.; Ho, Y.; Yu, L.; Kwang, J. WSSV ie1 promoter is more efficient than CMV promoter to express H5 hemagglutinin from influenza virus in baculovirus as a chicken vaccine. BMC Microbiol. 2008, 8, 238. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Wang, Y.Z.; Pan, Q.H.; Wang, Q.; Yuan, J.F.; Chen, T.S. Establishment of a cell line from the kidney of black carp and its susceptibility to spring viremia of carp virus. J. Fish Dis. 2018, 41, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Winkler, C.; Schartl, M. Production of medakafish chimeras from a stable embryonic stem cell line. Proc. Natl. Acad. Sci. USA 1998, 95, 3679–3684. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Liu, T.; Zhao, H.; Xu, H.; Wang, W.; Liu, R.; Chen, T.; Deng, J.; Gui, J. Establishment of a normal medakafish spermatogonial cell line capable of sperm production in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 8011–8016. [Google Scholar] [CrossRef] [PubMed]
- Driever, W.; Rangini, Z. Characterization of a cell line derived from zebrafish (Brachydanio rerio) embryos. In Vitro Cell Dev. Biol. Anim. 1993, 29A, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Hong, N.; Lu, W.; Zeng, H.; Song, J.; Hong, Y. Fusion gene vectors allowing for simultaneous drug selection, cell labeling, and reporter assay in vitro and in vivo. Anal. Chem. 2012, 84, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chang, Y.S.; Wang, C.H.; Kou, G.H.; Lo, C.F. Microarray and RT-PCR screening for white spot syndrome virus immediate-early genes in cycloheximide-treated shrimp. Virology 2005, 334, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chang, Y.S.; Wang, A.H.; Kou, G.H.; Lo, C.F. White spot syndrome virus annexes a shrimp STAT to enhance expression of the immediate-early gene ie1. J. Virol. 2007, 81, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.L.; Luke, G.; Mehrotra, A.; Li, X.; Hughes, L.E.; Gani, D.; Ryan, M.D. Analysis of the aphthovirus 2A/2B polyprotein ′cleavage′ mechanism indicates not a proteolytic reaction, but a novel translational effect: A putative ribosomal ′skip′. J. Gen. Virol. 2001, 82, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoint. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Fang, J.; Chen, T.; Pan, Q.; Wang, Q. Generation of albino medaka (Oryzias latipes) by CRISPR/Cas9. J. Exp. Zool. B Mol. Dev. Evol. 2018, 330, 242–246. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Fang, J.; Pan, Q.; Wang, Y.; Xue, T.; Li, L.; Chen, T. Efficient and Stable Delivery of Multiple Genes to Fish Cells by a Modified Recombinant Baculovirus System. Int. J. Mol. Sci. 2018, 19, 3767. https://doi.org/10.3390/ijms19123767
Wang Q, Fang J, Pan Q, Wang Y, Xue T, Li L, Chen T. Efficient and Stable Delivery of Multiple Genes to Fish Cells by a Modified Recombinant Baculovirus System. International Journal of Molecular Sciences. 2018; 19(12):3767. https://doi.org/10.3390/ijms19123767
Chicago/Turabian StyleWang, Qian, Jian Fang, Qihua Pan, Yizhou Wang, Ting Xue, Lingyu Li, and Tiansheng Chen. 2018. "Efficient and Stable Delivery of Multiple Genes to Fish Cells by a Modified Recombinant Baculovirus System" International Journal of Molecular Sciences 19, no. 12: 3767. https://doi.org/10.3390/ijms19123767
APA StyleWang, Q., Fang, J., Pan, Q., Wang, Y., Xue, T., Li, L., & Chen, T. (2018). Efficient and Stable Delivery of Multiple Genes to Fish Cells by a Modified Recombinant Baculovirus System. International Journal of Molecular Sciences, 19(12), 3767. https://doi.org/10.3390/ijms19123767