Enantiomeric Effect of d-Amino Acid Substitution on the Mechanism of Action of α-Helical Membrane-Active Peptides


Abstract
:1. Introduction
2. Results
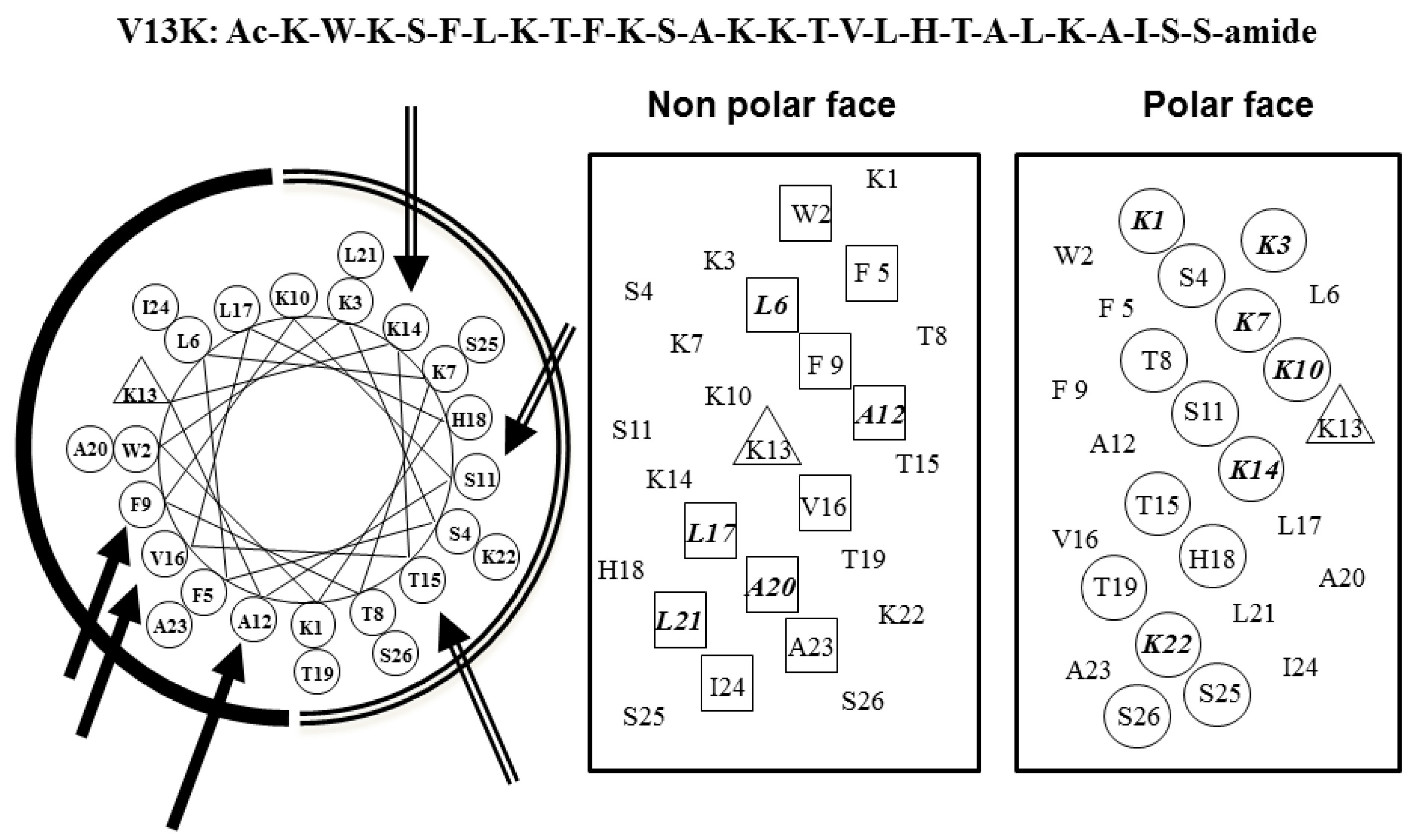
2.1. Peptide Design
2.2. Peptide Secondary Structure
2.3. Peptide Hydrophobicity
2.4. Biological Activity
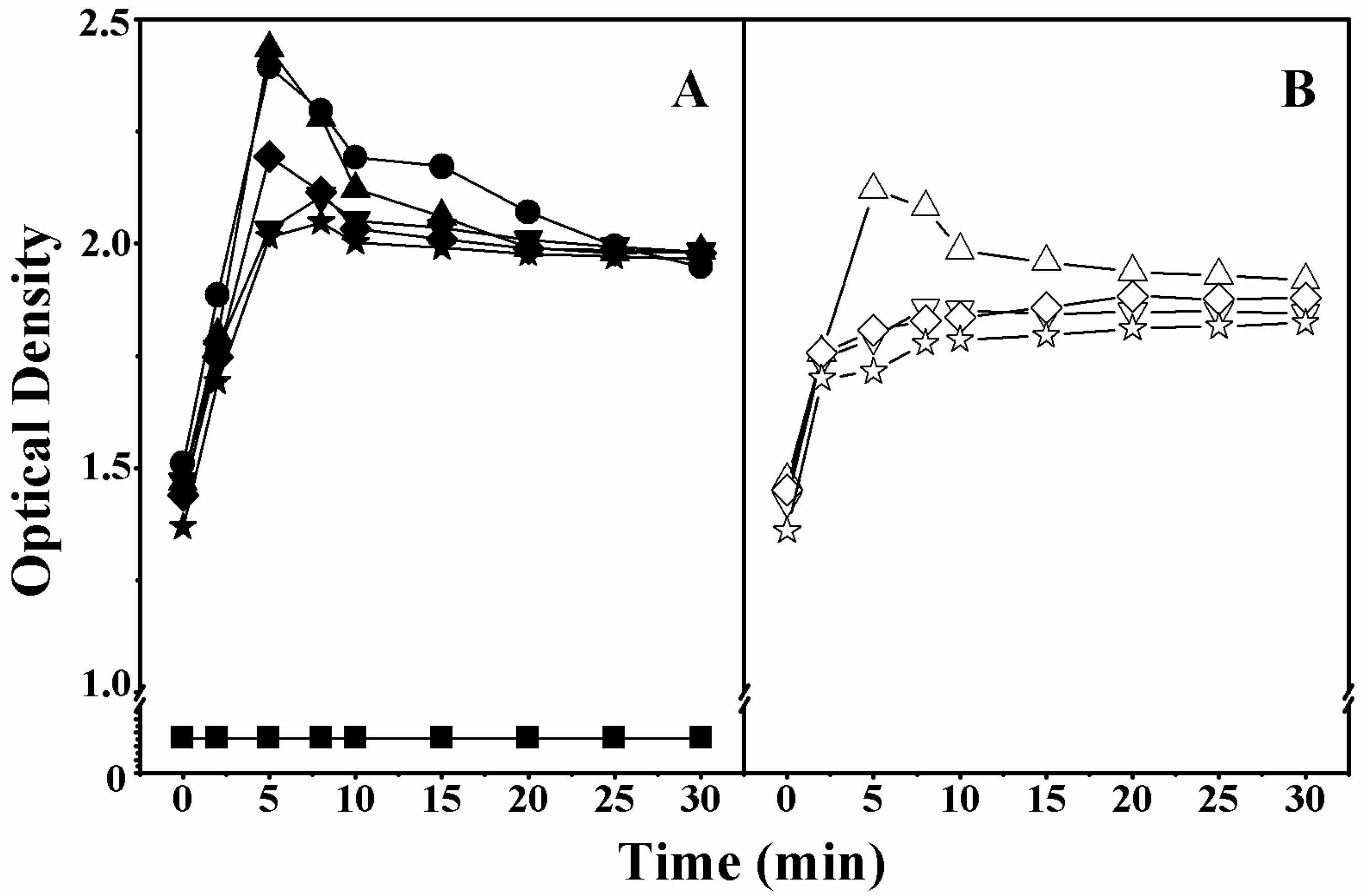
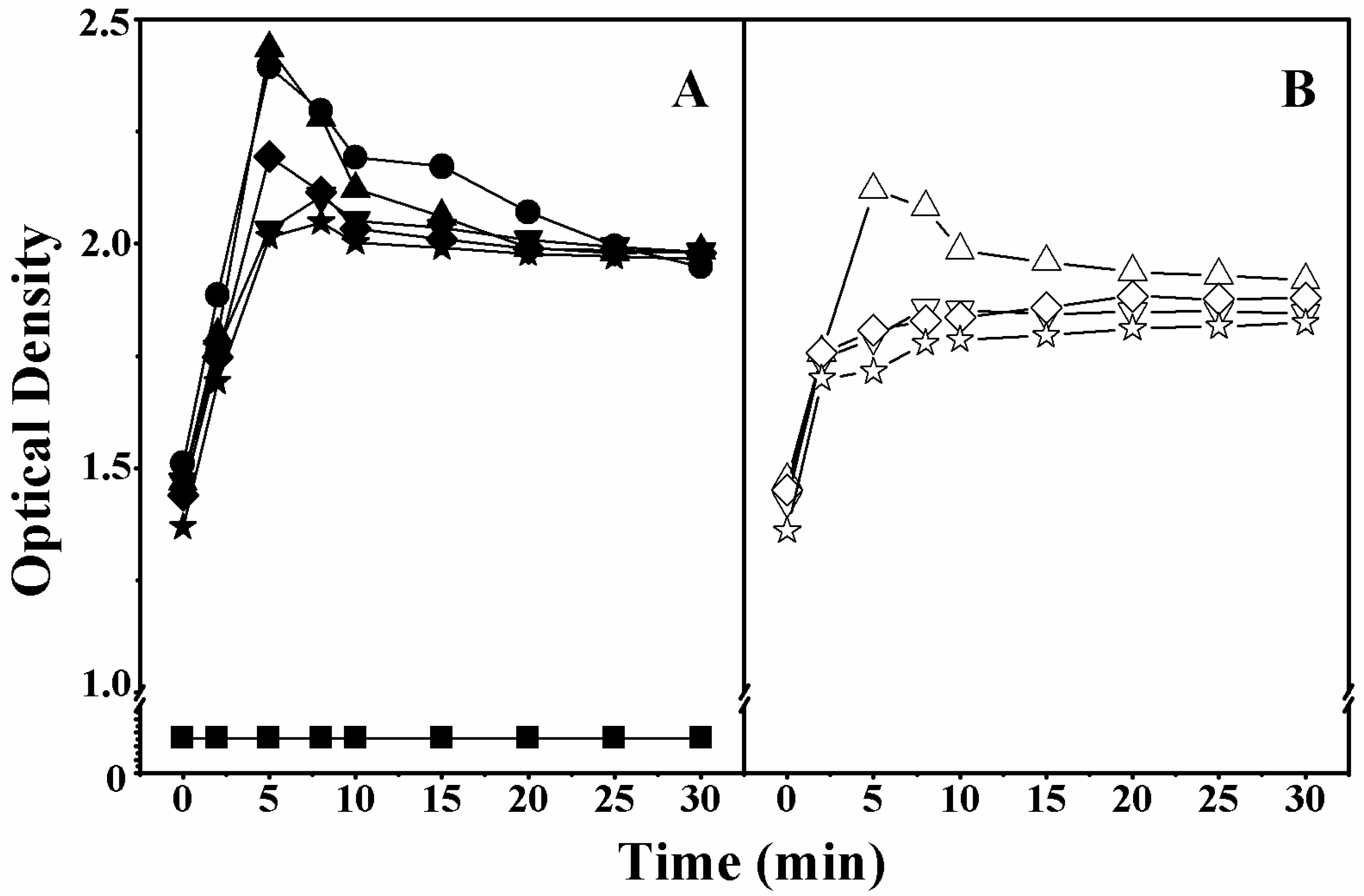
2.5. Cell Membrane Permeabilization
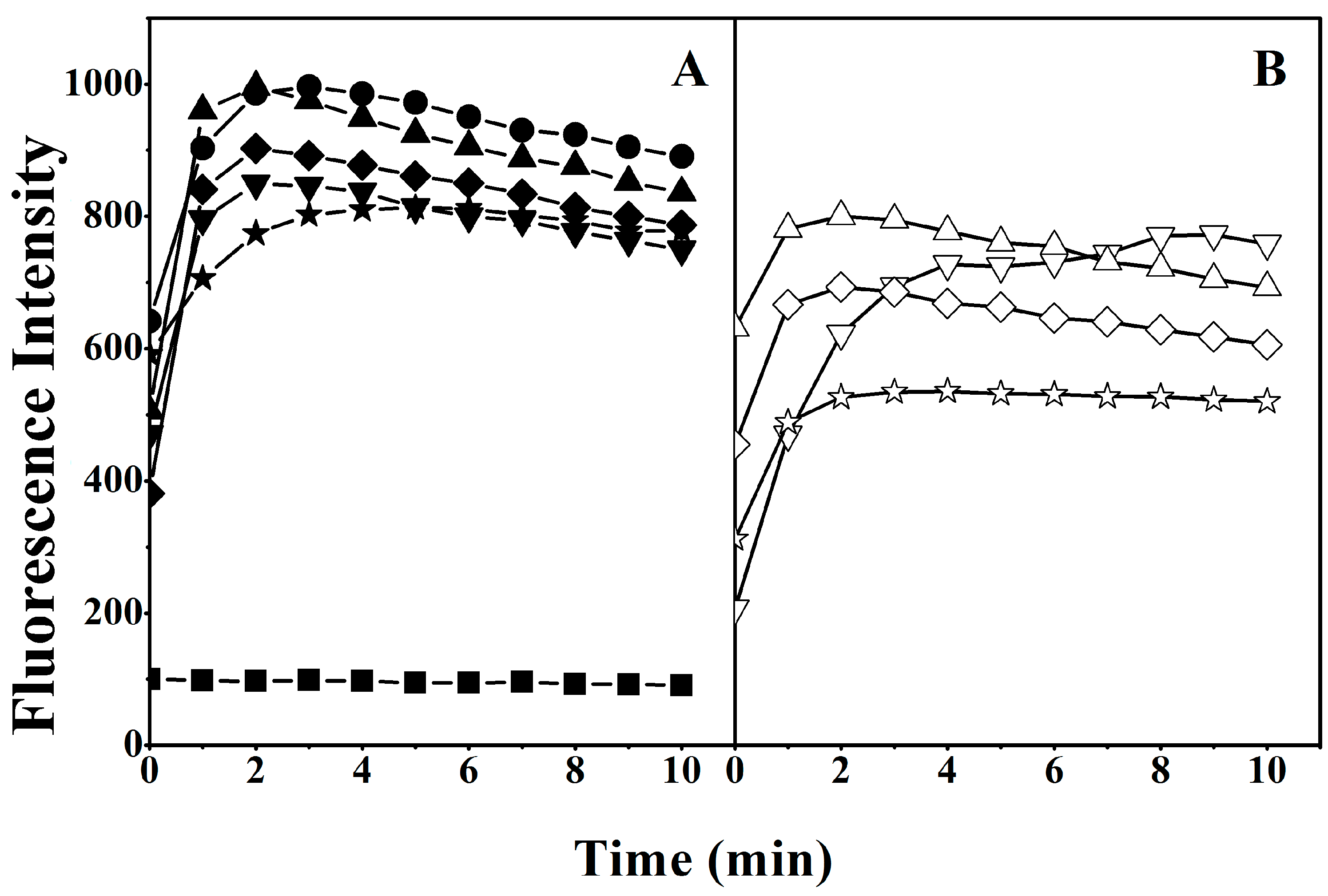
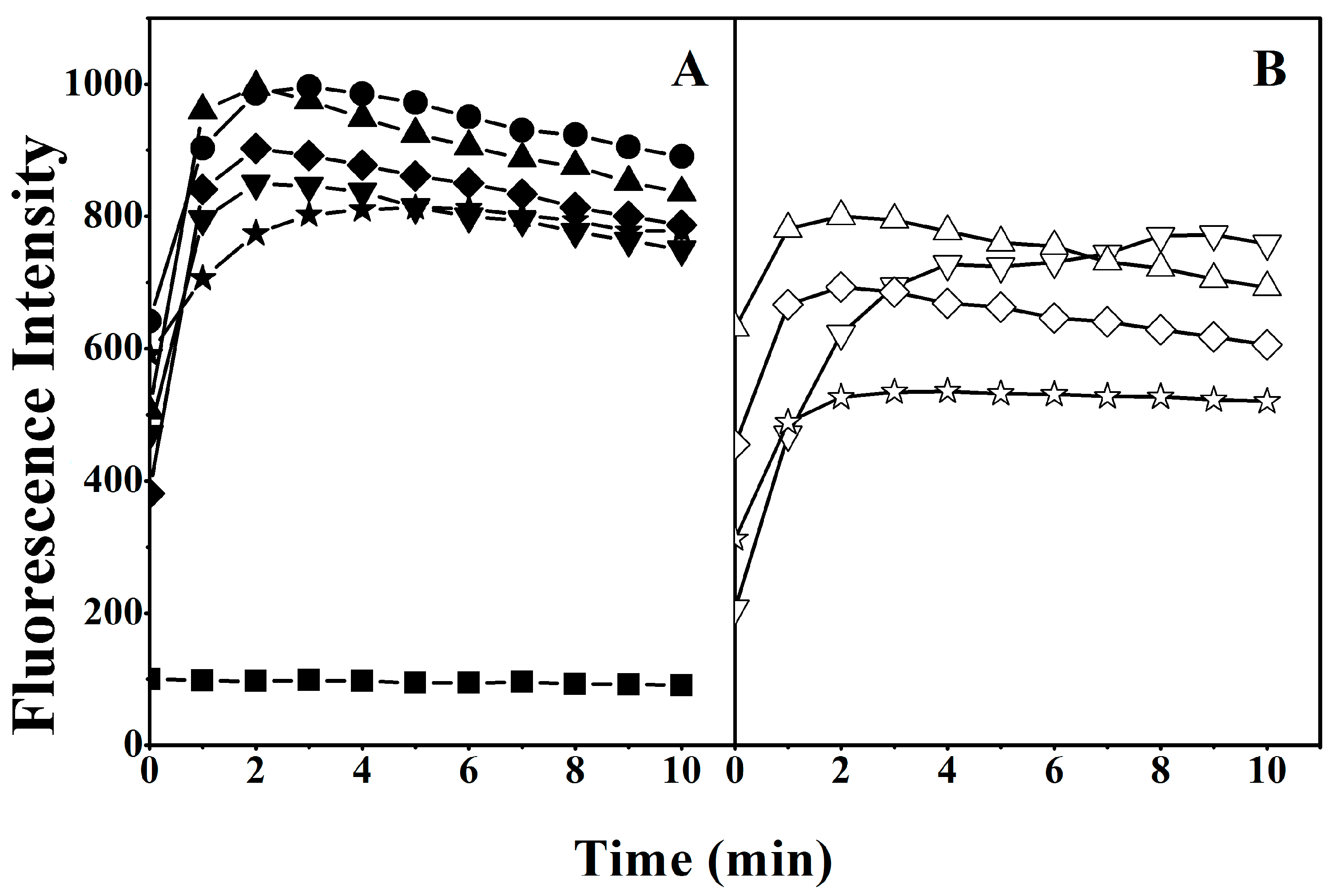
2.6. Interaction of Peptides with Liposome Model Membranes
2.7. AFM Imaging
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Peptide Synthesis and Purification
4.3. Circular Dichroism (CD) Analysis
4.4. Antimicrobial Activity Assay
4.5. Cell Culture and Analysis
4.6. Hemolytic Activity Assay
4.7. Outer Membrane Permeability Assay
4.8. Inner Membrane Permeability Assay
4.9. Preparation of Large Unilamellar Vesicles (LUVs)
4.10. Tryptophan Fluorescence and Quenching Assay
4.11. AFM Imaging
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MAPs | cationic membrane-active peptides |
| AFM | atomic force microscopy |
| KP | potassium phosphate |
| TFE | trifluoroethanol |
| RP-HPLC | reversed-phase HPLC |
| MIC | minimal inhibitory concentration |
| MHC | minimal hemolytic concentration |
| Chol | cholesterol |
| PC | chicken egg phosphatidylcholine |
| PS | porcine brain phosphatidylserine |
| PE | linear dichroism |
| PG | E. coli phosphatidylglycerol |
| SM | porcine brain sphingomyelin |
| TFA | trifluoroacetic acid |
| CD | Circular Dichroism |
| MH | Mueller–Hinton |
| ONPG | o-nitrophenyl-β-d-galactoside |
| NPN | 1-N-phenylnaphthylamine |
| HEPES | 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid |
| MTT | 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide |
| DMSO | dimethyl sulfoxide |
| AAC | Acoustic AC |
References
- Gatti, L.; Zunino, F. Overview of tumor cell chemoresistance mechanisms. Meth. Mol. Med. 2005, 111, 127–148. [Google Scholar]
- Gottesman, M.M. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiang, Q.; Zhang, Q.; Huang, Y.; Su, Z. Overview on the recent study of antimicrobial peptides: Origins, functions, relative mechanisms and application. Peptides 2012, 37, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, S.A.; Gelmon, K.; Mayer, L.D.; Hancock, R.E.; Bally, M.B. In vitro characterization of the anticancer activity of membrane-active cationic peptides. I. Peptide-mediated cytotoxicity and peptide-enhanced cytotoxic activity of doxorubicin against wild-type and p-glycoprotein over-expressing tumor cell lines. Anti-Cancer Drug Des. 2000, 15, 151–160. [Google Scholar]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta 1999, 1462, 55–70. [Google Scholar] [CrossRef]
- Maher, S.; McClean, S. Melittin exhibits necrotic cytotoxicity in gastrointestinal cells which is attenuated by cholesterol. Biochem. Pharmacol. 2008, 75, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhao, G.; Huang, Y.; Cai, M.; Shan, Y.; Wang, H.; Chen, Y. Specificity and mechanism of action of alpha-helical membrane-active peptides interacting with model and biological membranes by single-molecule force spectroscopy. Sci. Rep. 2016, 6, 29145. [Google Scholar] [CrossRef] [PubMed]
- Papo, N.; Shai, Y. New lytic peptides based on the d,l-amphipathic helix motif preferentially kill tumor cells compared to normal cells. Biochemistry 2003, 42, 9346–9354. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Cappiello, F.; Cohen, H.; Casciaro, B.; Luca, V.; Pini, A.; Di, Y.P.; Shai, Y.; Mangoni, M.L. d-Amino acids incorporation in the frog skin-derived peptide esculentin-1a(1-21)NH2 is beneficial for its multiple functions. Amino Acids 2015, 47, 2505–2519. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mant, C.T.; Farmer, S.W.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S. Rational design of alpha-helical antimicrobial peptides with enhanced activities and specificity/therapeutic index. J. Biol. Chem. 2005, 280, 12316–12329. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.B.; Wang, X.F.; Wang, H.Y.; Liu, Y.; Chen, Y. Studies on mechanism of action of anticancer peptides by modulation of hydrophobicity within a defined structural framework. Mol. Cancer Ther. 2011, 10, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; He, L.; Li, G.; Zhai, N.; Jiang, H.; Chen, Y. Role of helicity of alpha-helical antimicrobial peptides to improve specificity. Protein Cell 2014, 5, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mant, C.T.; Hodges, R.S. Determination of stereochemistry stability coefficients of amino acid side-chains in an amphipathic alpha-helix. J. Pept. Res. 2002, 59, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hao, D.; Chen, Y.; Xu, Y.; Tan, J.; Huang, Y.; Li, F.; Chen, Y. Inhibitory effects and mechanisms of physiological conditions on the activity of enantiomeric forms of an alpha-helical antibacterial peptide against bacteria. Peptides 2011, 32, 1488–1495. [Google Scholar] [CrossRef] [PubMed]
- Monera, O.D.; Sereda, T.J.; Zhou, N.E.; Kay, C.M.; Hodges, R.S. Relationship of sidechain hydrophobicity and alpha-helical propensity on the stability of the single-stranded amphipathic alpha-helix. J. Pept. Sci. 1995, 1, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.E.; Monera, O.D.; Kay, C.M.; Hodges, R.S. alpha-Helical propensities of amino acids in the hydrophobic face of an amphipathic alpha-helix. Protein Pept. Lett. 1994, 1, 114–119. [Google Scholar]
- Zhao, J.; Huang, Y.; Liu, D.; Chen, Y. Two hits are better than one: Synergistic anticancer activity of alpha-helical peptides and doxorubicin/epirubicin. Oncotarget 2015, 6, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Cai, M.; Xu, H.; Ding, B.; Hao, X.; Jiang, J.; Sun, Y.; Wang, H. Atomic force microscopy of asymmetric membranes from turtle erythrocytes. Mol. Cells 2014, 37, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Huang, Y.; Chen, Y. Production of an antimicrobial peptide AN5-1 in Escherichia coli and its dual mechanisms against bacteria. Chem. Biol. Drug Des. 2015, 85, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, K.; Dang, W.; Chen, R.; Xie, J.; Zhang, B.; Song, J.; Wang, R. Two hits are better than one: Membrane-active and DNA binding-related double-action mechanism of NK-18, a novel antimicrobial peptide derived from mammalian NK-lysin. Antimicrob. Agents Chemother. 2013, 57, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Huang, Y.; Feng, Q.; Chen, Y. Tryptophan as a probe to study the anticancer mechanism of action and specificity of alpha-helical anticancer peptides. Molecules 2014, 19, 12224–12241. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Gao, Z.; Liu, Y.; Cheng, Y.; Yu, M.; Zhao, L.; Duan, Y.; Liu, Y. Breath ketone testing: A new biomarker for diagnosis and therapeutic monitoring of diabetic ketosis. BioMed. Res. Int. 2014, 2014, 869186. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hao, X.; Liu, D.; Huang, Y.; Chen, Y. In vitro Characterization of the Rapid Cytotoxicity of Anticancer Peptide HPRP-A2 through Membrane Destruction and Intracellular Mechanism against Gastric Cancer Cell Lines. PLoS ONE 2015, 10, e0139578. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Lee, P.Y.; Ho, B.; Ding, J.L.; Lim, C.T. Atomic force microscopy study of the antimicrobial action of Sushi peptides on Gram negative bacteria. Biochim. Biophys. Acta 2007, 1768, 411–418. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Peptide | Amino Acid Sequence a |
|---|---|---|
| Parent | V13K | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide |
| Polar face group | K14D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide |
| S11D/K14D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide | |
| K14D/T15D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide | |
| S11D/K14D/T15D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide | |
| Non-polar face group | A12D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide |
| F9D/A12D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide | |
| A12D/V16D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide | |
| F9D/A12D/V16D | Ac-K-W-K-S-F-L-K-T-F-K-S-A-K-K-T-V-L-H-T-A-L-K-A-I-S-S-amide |
| Peptides a | tR (min) b | Benign c | 50% TFE d | ||
|---|---|---|---|---|---|
| 25 °C | [θ]222 | α-Helix (%) e | [θ]222 | α-Helix (%) e | |
| V13K | 44.48 | −5617.46 | 11.1% | −50,520.3 | 100% |
| K14D | 41.93 | −2223.13 | 4.4% | −38,897.2 | 77.0% |
| S11D/K14D | 40.77 | −3524.85 | 7.0% | −25,931.3 | 51.3% |
| K14D/T15D | 40.41 | −1862.4 | 3.7% | −28,551.5 | 56.5% |
| S11D/K14D/T15D | 39.59 | −2845.85 | 5.6% | −19,537.5 | 38.7% |
| A12D | 41.46 | −4130.9 | 8.2% | −35,785.1 | 70.8% |
| F9D/A12D | 40.04 | −2319.89 | 4.6% | −19,328.6 | 38.3% |
| A12D/V16D | 39.79 | −3435.72 | 6.8% | −23,452.9 | 46.4% |
| F9D/A12D/V16D | 39.06 | −1907.83 | 3.8% | −18,393.1 | 36.4% |
| Peptides | MHC a (μM) | G+ MIC b (μM) | G− MIC b (μM) | ||
|---|---|---|---|---|---|
| S. aureus | S. epidermidis | P. aeruginosa | E. coli | ||
| ATCC25923 | ATCC12228 | ATCC27853 | ATCC25922 | ||
| V13K | 250 | 32 | 4 | 8 | 8 |
| K14D | 125 | 32 | 16 | 16 | 32 |
| S11D/K14D | 125 | 64 | 16 | 32 | 64 |
| K14D/T15D | 250 | 64 | 16 | 32 | 64 |
| S11D/K14D/T15D | 250 | 64 | 16 | 125 | 125 |
| A12D | 125 | 64 | 16 | 16 | 32 |
| F9D/A12D | 250 | 125 | 64 | 32 | 125 |
| A12D/V16D | >500 | 125 | 125 | 32 | 125 |
| F9D/A12D/V16D | >500 | 125 | 125 | 125 | 125 |
| Peptides | IC50 (μM) a | |||
|---|---|---|---|---|
| HeLa | MIA PaCa-2 | HPAC | BxPC-3 | |
| V13K | 16 ± 0.5 | 24.8 ± 0.6 | 7.6 ± 4.0 | 12.5 ± 1.2 |
| K14D | 15.6 ± 4.1 | 17.9 ± 2.3 | 7.2 ± 5.2 | 20.0 ± 0.8 |
| S11D/K14D | 30.6 ± 0.5 | 41.3 ± 2.6 | 25.5 ± 0.5 | 22.4 ± 2.3 |
| K14D/T15D | 31.6 ± 2.1 | 47.1 ± 5.3 | 22.5 ± 0.5 | 24.1 ± 5.5 |
| S11D/K14D/T15D | 41.1 ± 0.5 | 56.8 ± 3.3 | 54.2 ± 1.2 | 45.5 ± 5.6 |
| A12D | 28.8 ± 0.8 | 29.9 ± 4.8 | 15.5 ± 1.7 | 45.1 ± 1.0 |
| F9D/A12D | 59.8 ± 0.5 | 38.8 ± 2.9 | 42.4 ± 3.0 | 125.0 ± 3.7 |
| A12D/V16D | 60.1 ± 1.2 | 46.2 ± 3.0 | 41.4 ± 1.4 | 125.0 ± 5.9 |
| F9D/A12D/V16D | 125.0 ± 3.4 | 57.9 ± 2.8 | 95.6 ± 1.8 | 125.0 ± 1.8 |
| Peptides | HEPES (nm) | PC/PG | PC/Chol | PC/SM/PE/PS/Chol | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Wavelength (nm) | Blue Shift (nm) | Intensity | Wavelength (nm) | Blue Shift (nm) | Intensity | Wavelength (nm) | Blue Shift (nm) | Intensity | ||
| V13K | 353 | 329 | 24 | 459 | 344 | 9 | 486 | 330 | 23 | 582 |
| K14D | 349 | 332 | 17 | 400 | 349 | 0 | 459 | 329 | 20 | 507 |
| S11D/K14D | 347 | 332 | 15 | 346 | 345 | 2 | 414 | 331 | 16 | 363 |
| K14D/T15D | 349 | 336 | 13 | 348 | 349 | 0 | 412 | 337 | 12 | 346 |
| S11D/K14D/T15D | 345 | 339 | 6 | 324 | 343 | 2 | 308 | 335 | 10 | 345 |
| A12D | 351 | 328 | 23 | 386 | 345 | 6 | 464 | 327 | 24 | 413 |
| F9D/A12D | 350 | 335 | 15 | 339 | 348 | 2 | 456 | 331 | 19 | 338 |
| A12D/V16D | 350 | 336 | 14 | 329 | 348 | 2 | 330 | 337 | 13 | 340 |
| F9D/A12D/V16D | 346 | 335 | 11 | 300 | 346 | 0 | 261 | 337 | 9 | 316 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Zhao, G.; Huang, Y.; Cai, M.; Yan, Q.; Wang, H.; Chen, Y. Enantiomeric Effect of d-Amino Acid Substitution on the Mechanism of Action of α-Helical Membrane-Active Peptides. Int. J. Mol. Sci. 2018, 19, 67. https://doi.org/10.3390/ijms19010067
Sun S, Zhao G, Huang Y, Cai M, Yan Q, Wang H, Chen Y. Enantiomeric Effect of d-Amino Acid Substitution on the Mechanism of Action of α-Helical Membrane-Active Peptides. International Journal of Molecular Sciences. 2018; 19(1):67. https://doi.org/10.3390/ijms19010067
Chicago/Turabian StyleSun, Shiyu, Guangxu Zhao, Yibing Huang, Mingjun Cai, Qiuyan Yan, Hongda Wang, and Yuxin Chen. 2018. "Enantiomeric Effect of d-Amino Acid Substitution on the Mechanism of Action of α-Helical Membrane-Active Peptides" International Journal of Molecular Sciences 19, no. 1: 67. https://doi.org/10.3390/ijms19010067
APA StyleSun, S., Zhao, G., Huang, Y., Cai, M., Yan, Q., Wang, H., & Chen, Y. (2018). Enantiomeric Effect of d-Amino Acid Substitution on the Mechanism of Action of α-Helical Membrane-Active Peptides. International Journal of Molecular Sciences, 19(1), 67. https://doi.org/10.3390/ijms19010067