Rhein Induces Oxidative Stress and Apoptosis in Mouse Blastocysts and Has Immunotoxic Effects during Embryonic Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
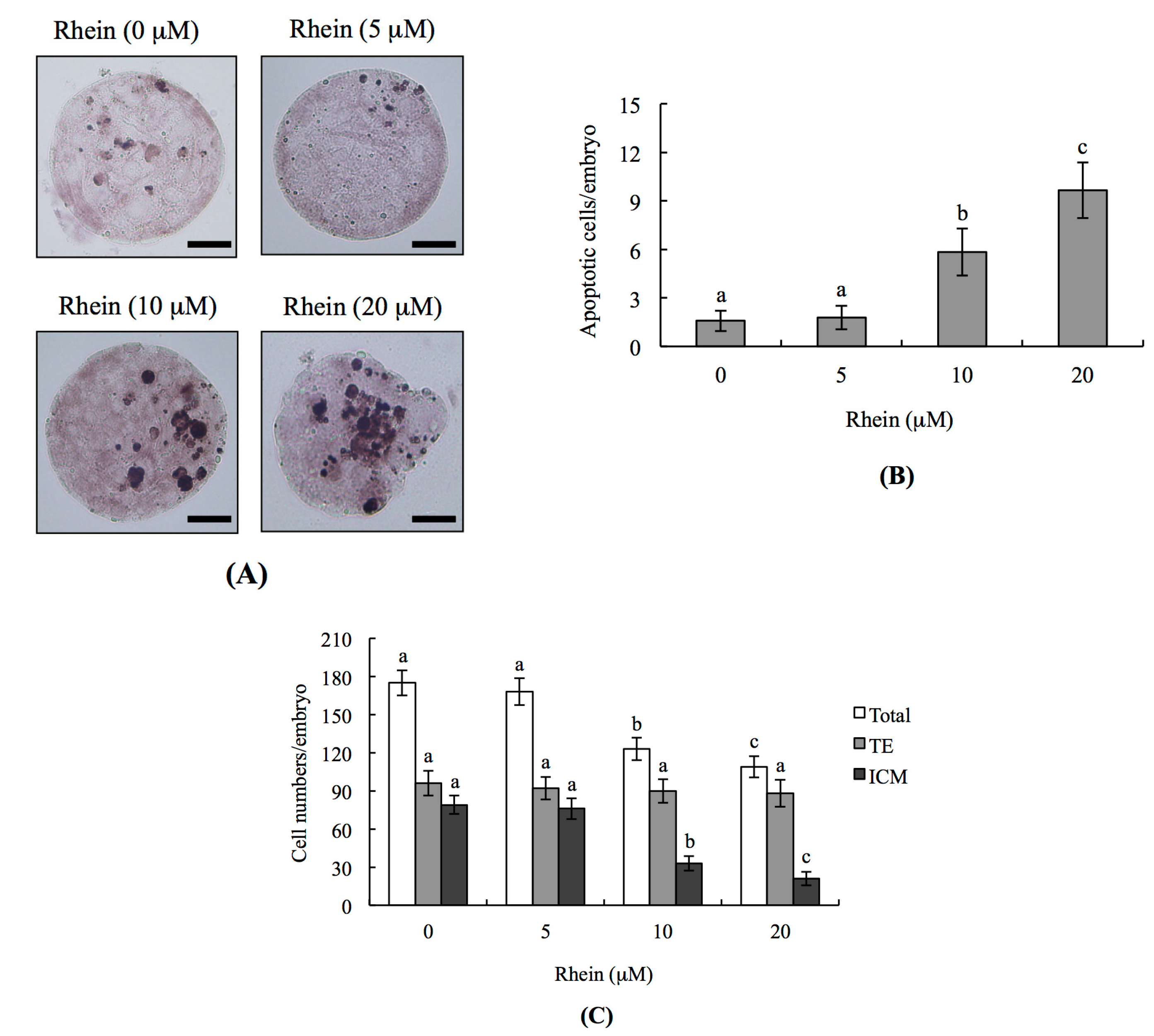
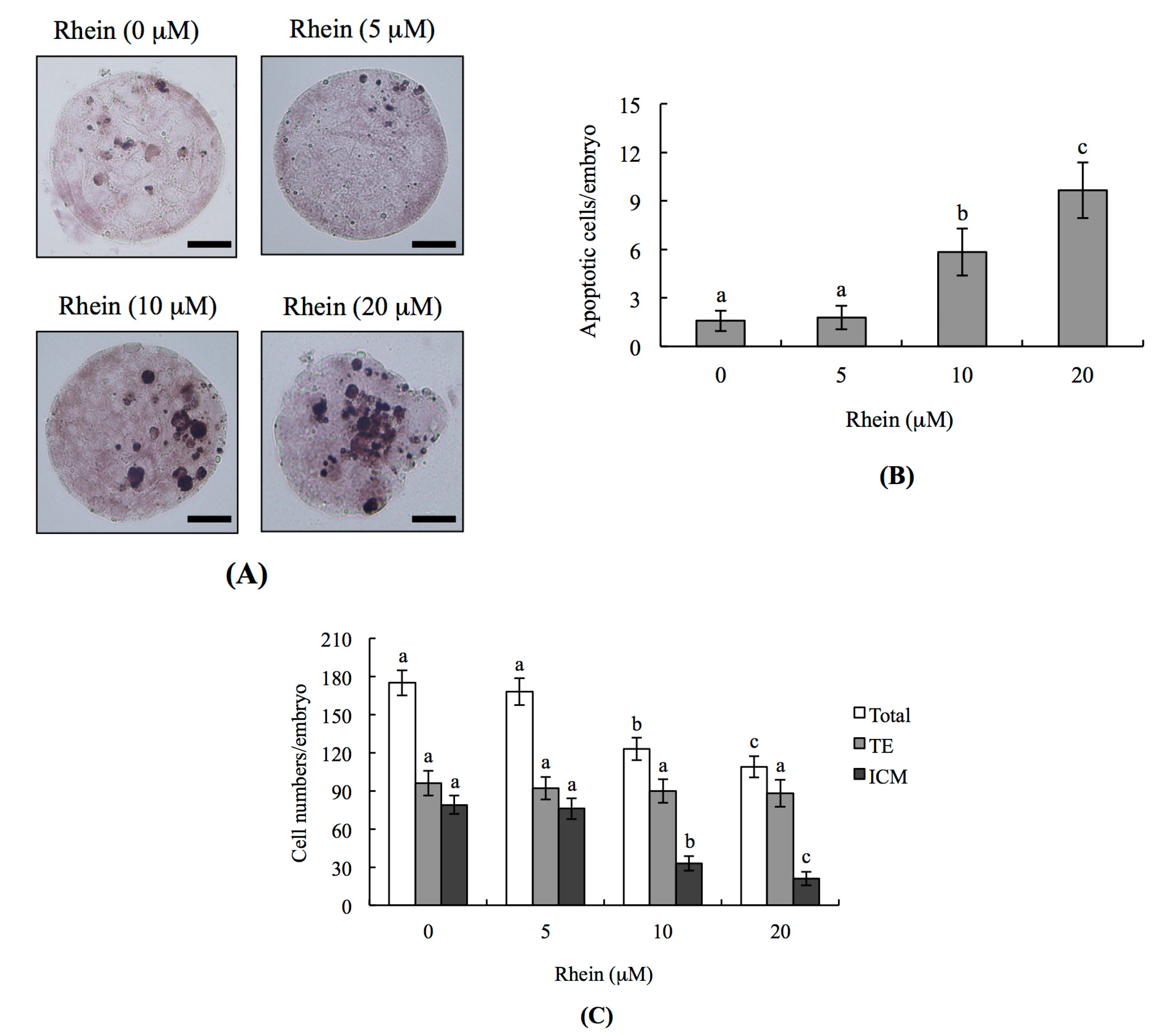
2.1. Effects of Rhein on Mouse Blastocysts
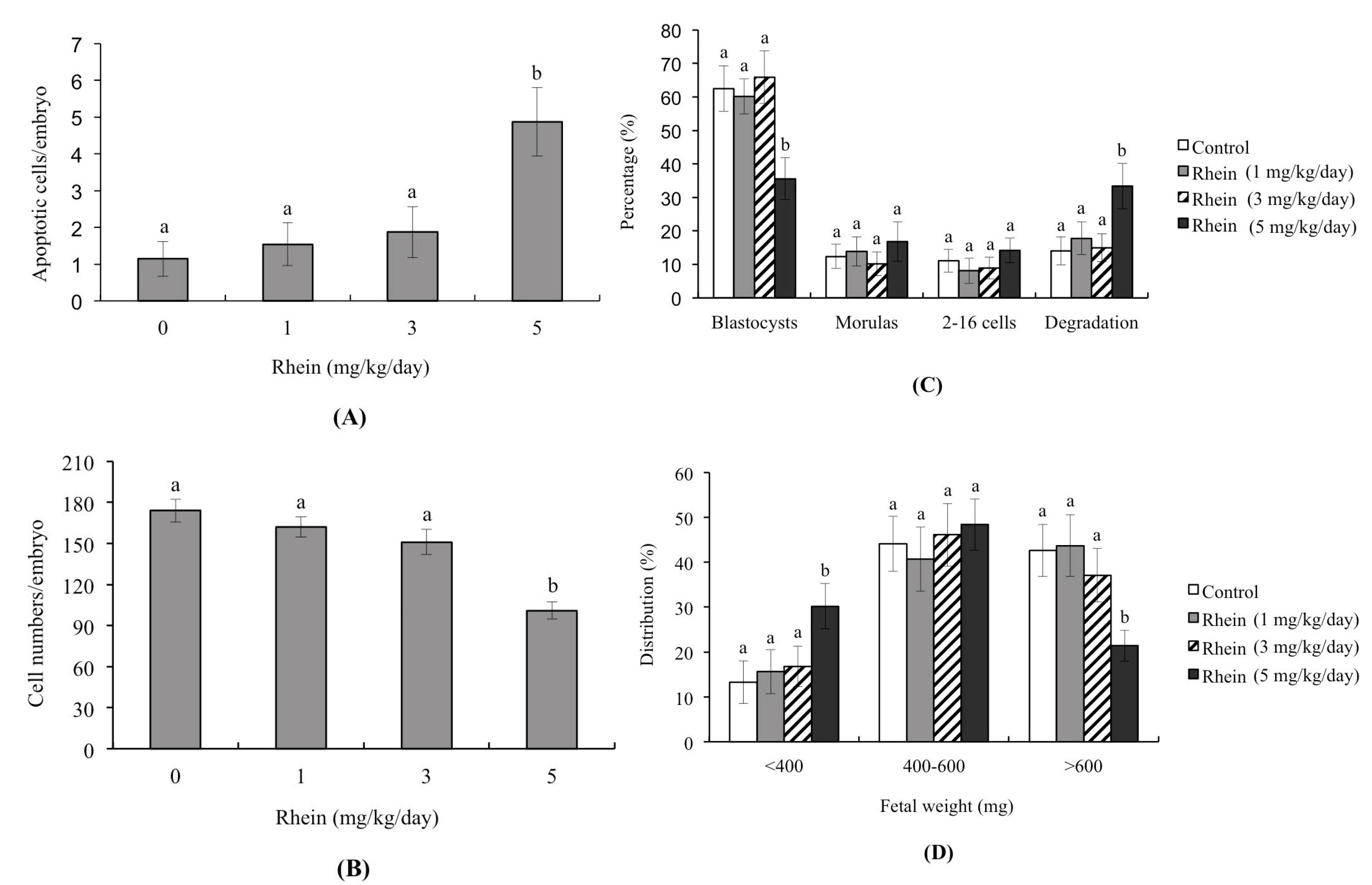
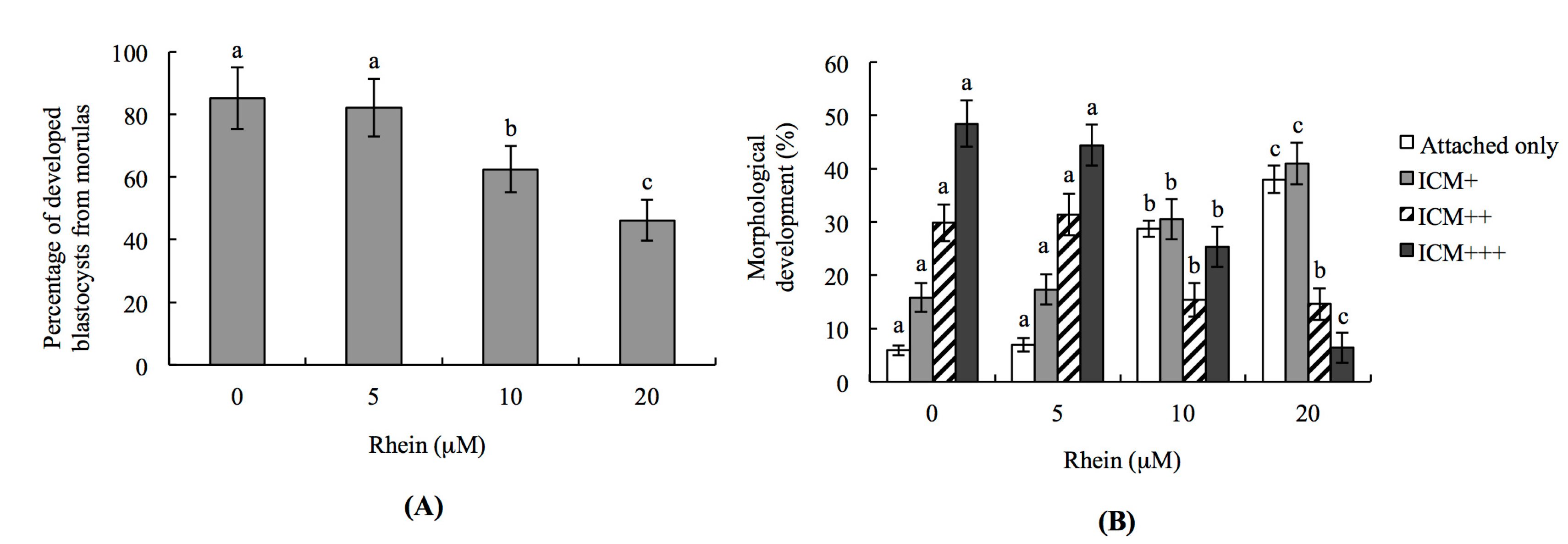
2.2. Impacts of Rhein on Mouse Embryonic Developmental Potential In Vitro
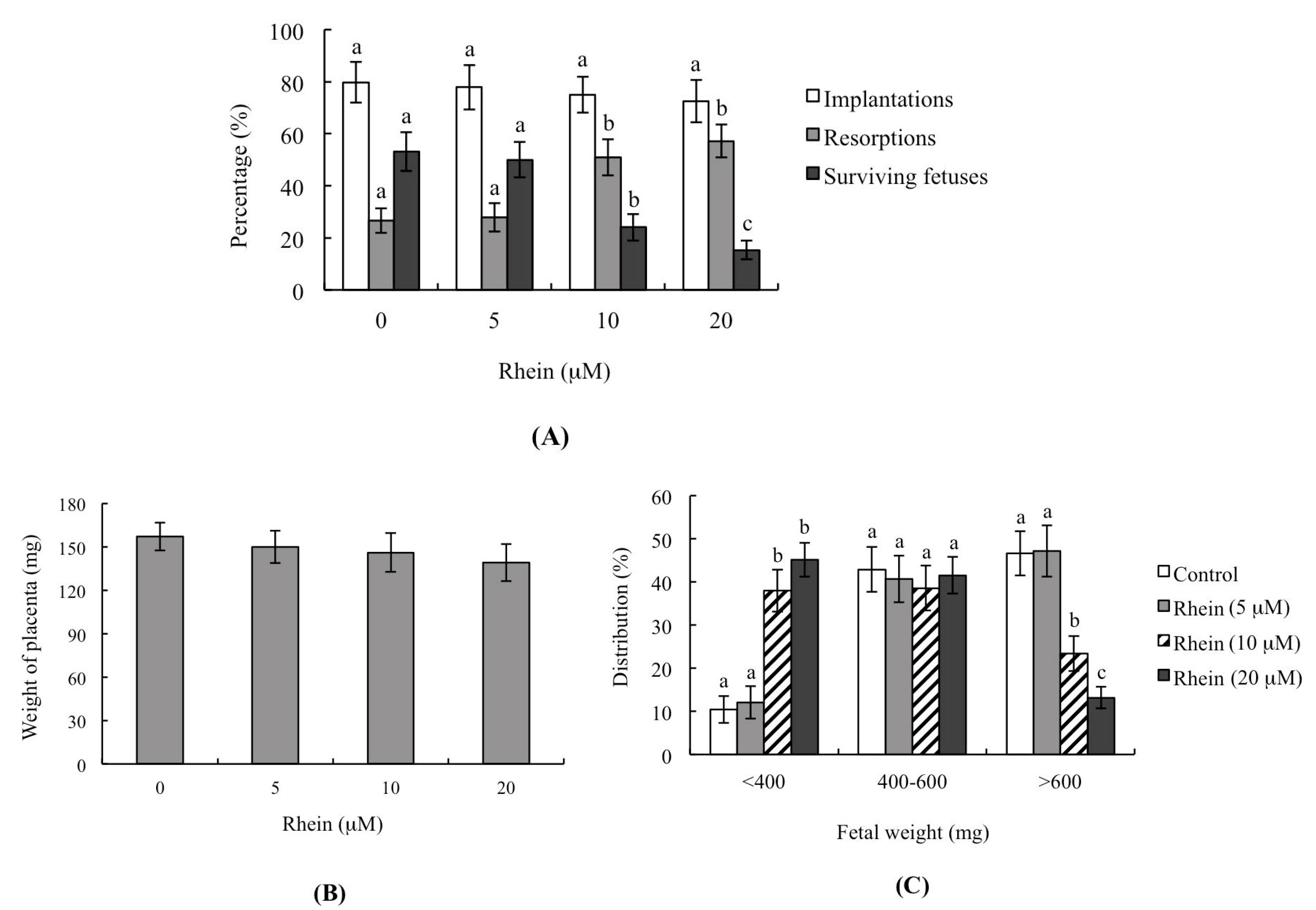
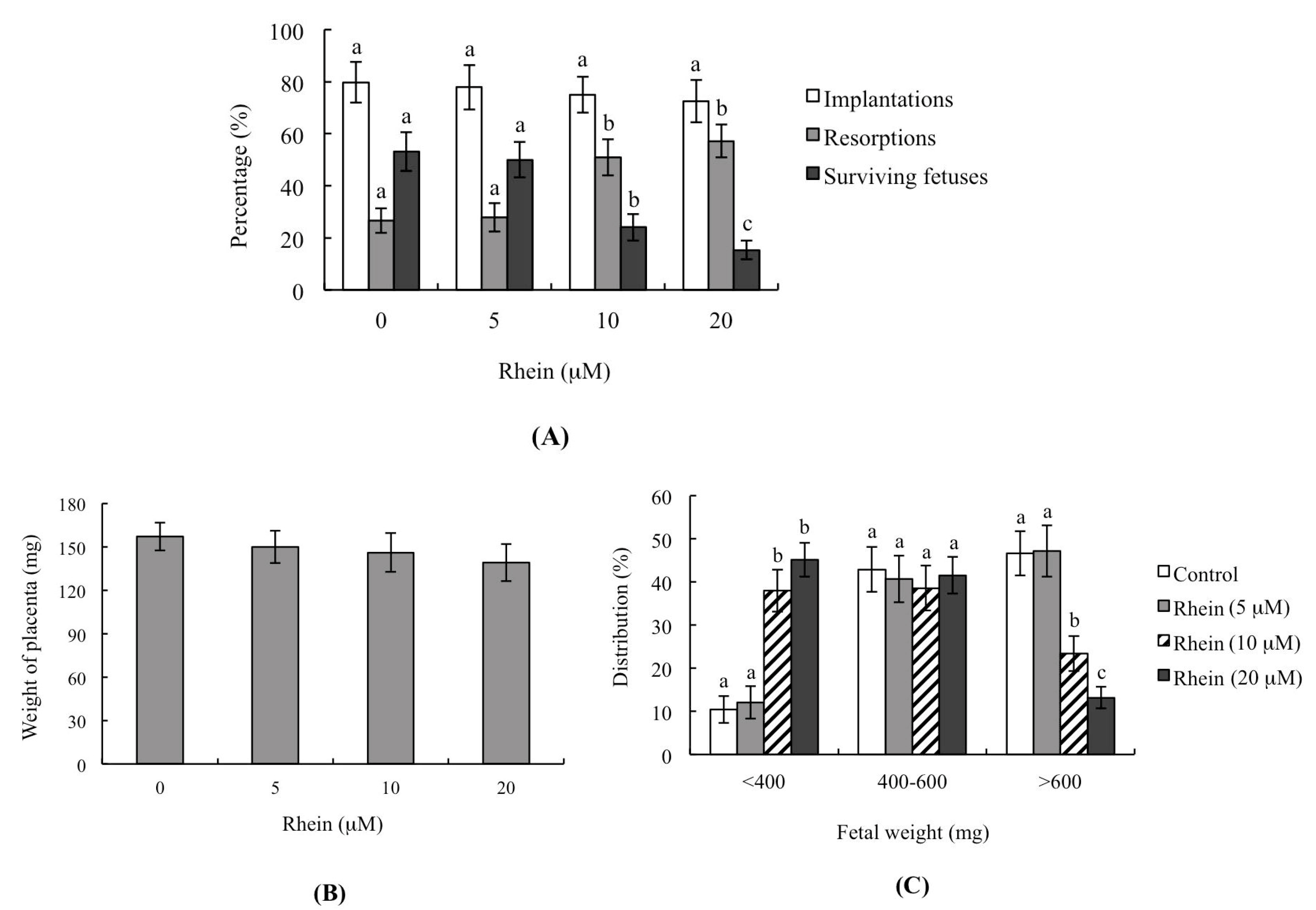
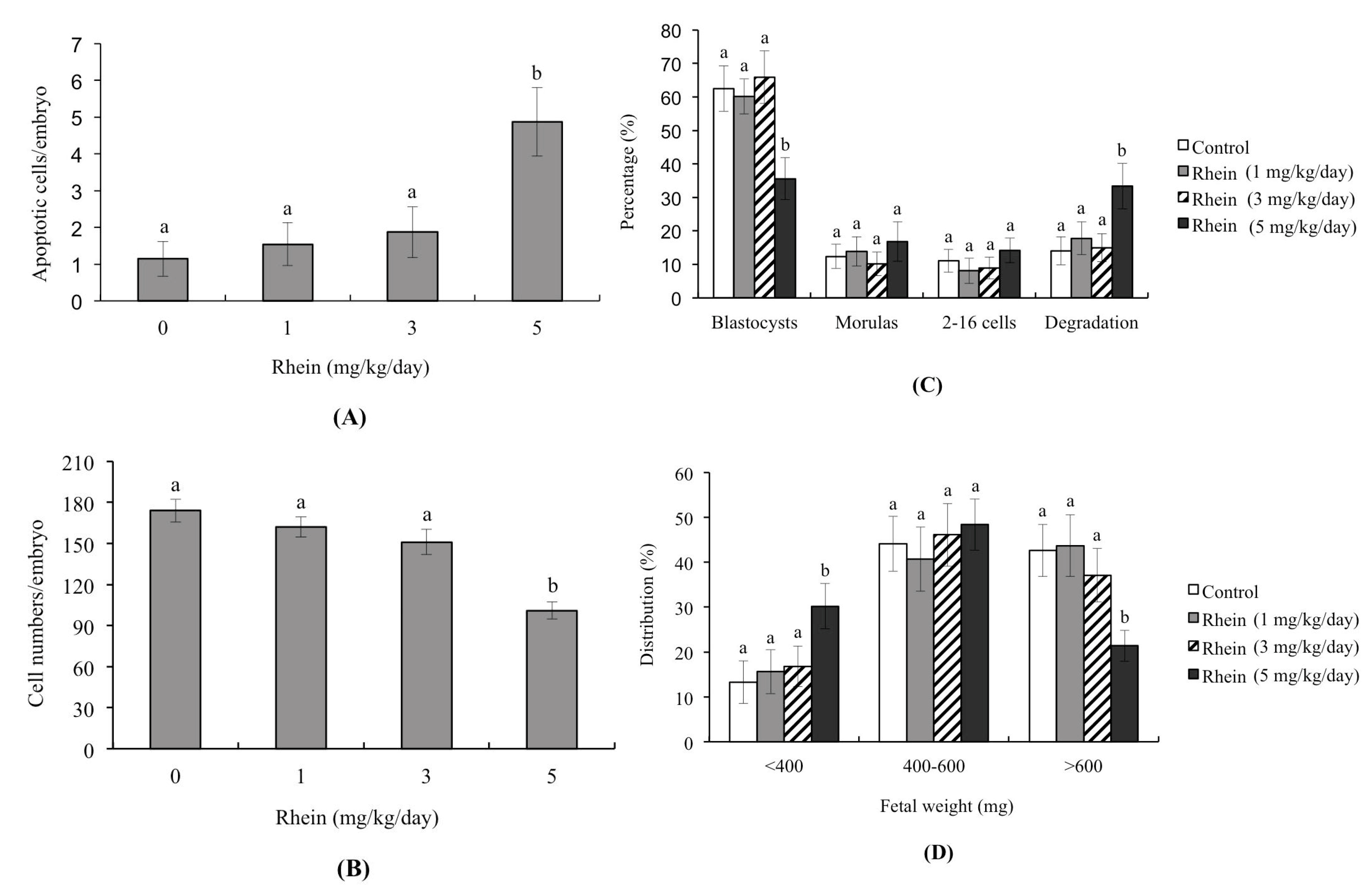
2.3. Effects of Rhein on Blastocyst Developmental Potential In Vivo
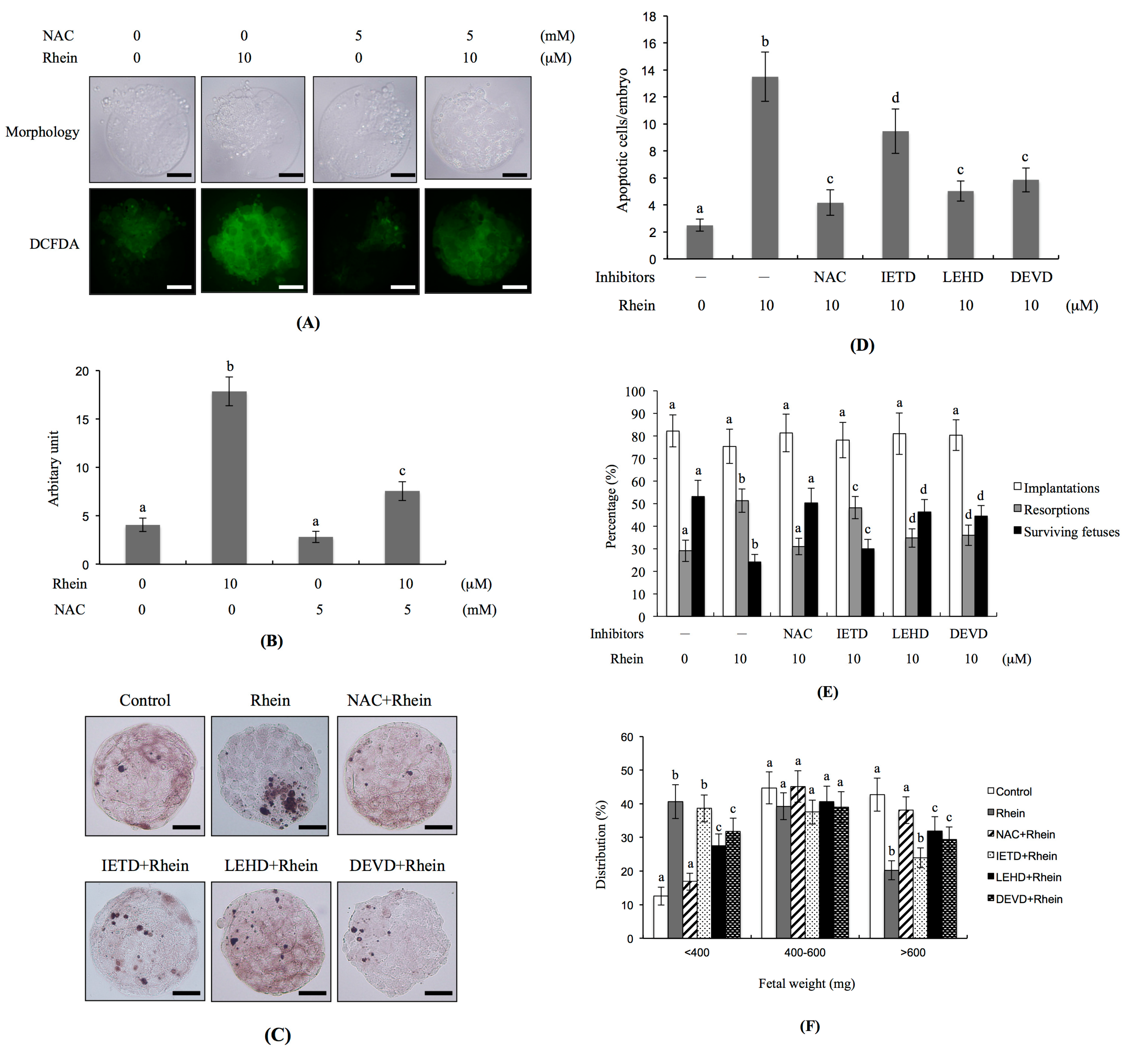
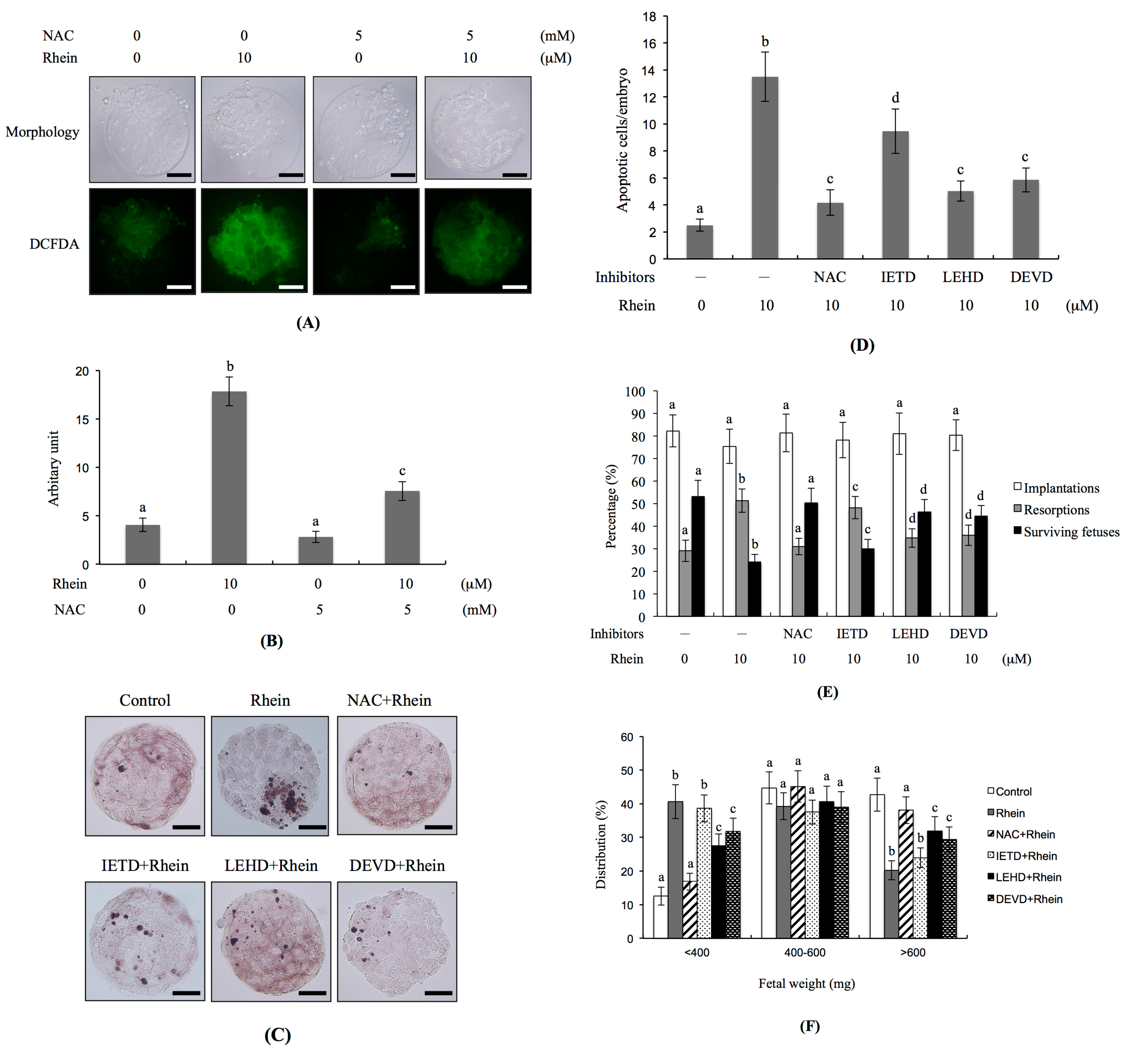
2.4. Effects of Antioxidants and Caspase Inhibitors on the Development of Rhein-Treated Blastocysts
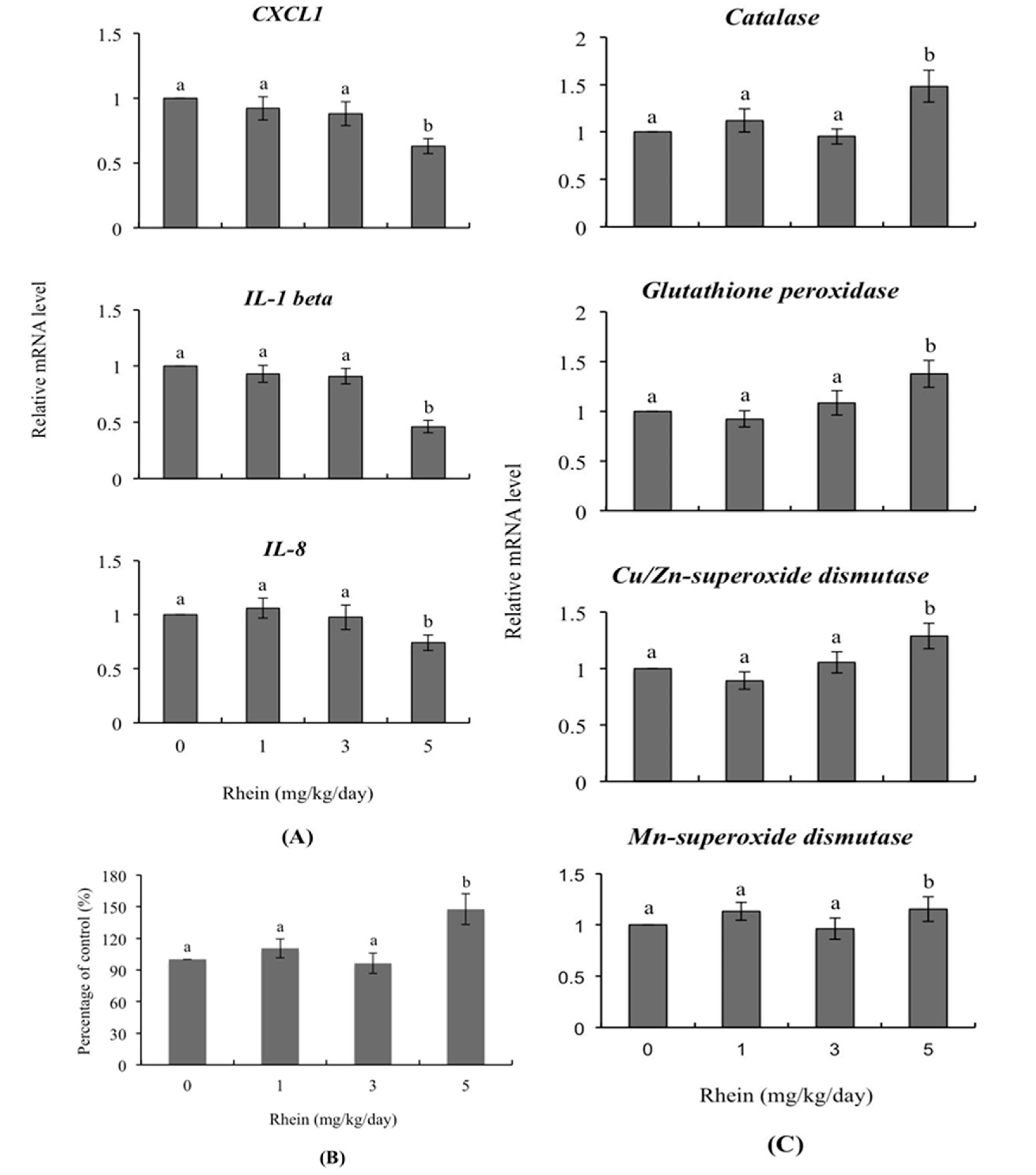
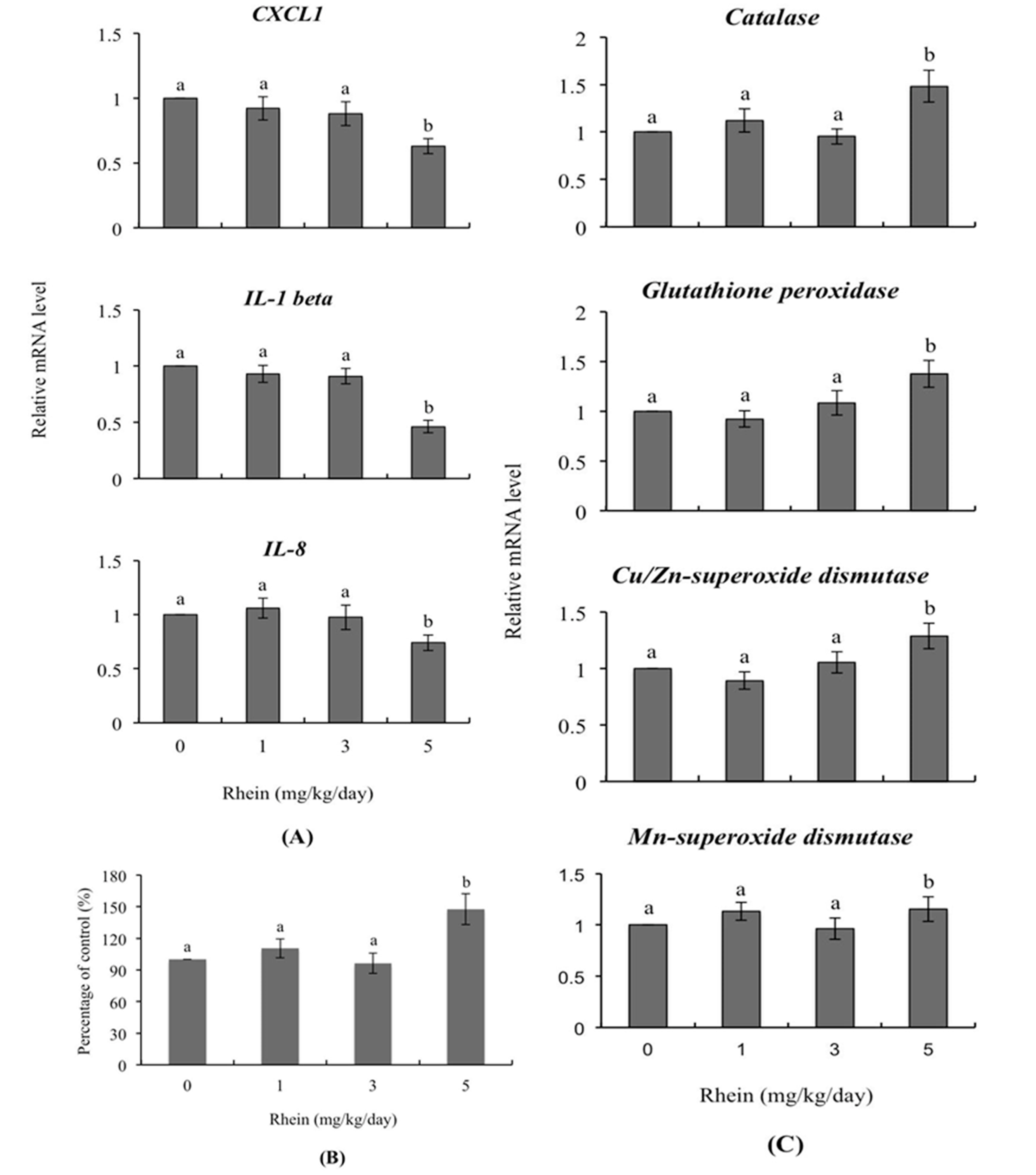
2.5. Transcription of Genes Related to Innate Immunity and Oxidative Stress in Day-Old Mice Born to Dams Intravenously Injected with Rhein during Pregnancy
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Mouse Morula and Blastocyst Collection
4.3. Analysis of Blastocysts Developed from Morulas
4.4. TUNEL Assay of Rhein-Treated Embryos
4.5. Cell Proliferation in Rhein-Treated Embryos
4.6. Embryonic Development via Morphological Analysis
4.7. Blastocyst Development Following Embryo Transfer
4.8. Intravenous Injection of Female Mice with Rhein and Collection of Blastocysts
4.9. Pre-Treatment of Blastocysts with NAC or Caspase Inhibitors
4.10. Gene Expression and ROS Content in Fetal Liver
4.11. Analysis of Gene Expression
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, J.H.; Kim, J.M.; Kim, C. Pharmacokinetic analysis of rhein in Rheum undulatum L. J. Ethnopharmacol. 2003, 84, 5–9. [Google Scholar] [CrossRef]
- Lee, B.H.; Huang, Y.Y.; Duh, P.D.; Wu, S.C. Hepatoprotection of emodin and Polygonum multiflorum against CCl(4)-induced liver injury. Pharm. Biol. 2012, 50, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Nadkarni, J.R.; Vishwakarma, R.A.; Bharate, S.B.; Nivsarkar, M.; Anandjiwala, S. The hydroalcoholic extract of Cassia alata (Linn.) leaves and its major compound rhein exhibits antiallergic activity via mast cell stabilization and lipoxygenase inhibition. J. Ethnopharmacol. 2012, 141, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.K.; Singh, S.S.; Verma, S.; Kumar, S. Antifungal activity of anthraquinone derivatives from Rheum emodi. J. Ethnopharmacol. 2000, 72, 43–46. [Google Scholar] [CrossRef]
- Cyong, J.; Matsumoto, T.; Arakawa, K.; Kiyohara, H.; Yamada, H.; Otsuka, Y. Anti-bacteroides fragilis substance from rhubarb. J. Ethnopharmacol. 1987, 19, 279–283. [Google Scholar] [PubMed]
- Barnard, D.L.; Huffman, J.H.; Morris, J.L.; Wood, S.G.; Hughes, B.G.; Sidwell, R.W. Evaluation of the antiviral activity of anthraquinones, anthrones and anthraquinone derivatives against human cytomegalovirus. Antiviral Res. 1992, 17, 63–77. [Google Scholar] [CrossRef]
- Delpino, A.; Paggi, M.G.; Gentile, P.F.; Castiglione, S.; Bruno, T.; Benass, M.; Floridi, A. Protein synthetic activity and adenylate energy charge in Rhein-treated cultured human glioma cells. Cancer Biochem. Biophys. 1992, 12, 241–252. [Google Scholar] [PubMed]
- Castiglione, S.; Fanciulli, M.; Bruno, T.; Evangelista, M.; del Carlo, C.; Paggi, M.G.; Chersi, A.; Floridi, A. Rhein inhibits glucose uptake in Ehrlich ascites tumor cells by alteration of membrane-associated functions. Anticancer Drugs 1993, 4, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Fujii, M.; Hou, D.X. Rhein induces apoptosis in HL-60 cells via reactive oxygen species-independent mitochondrial death pathway. Arch. Biochem. Biophys. 2003, 418, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Chan, H.L.; Lin, H.Y.; Way, T.D.; Kao, M.C.; Song, M.Z.; Lin, Y.J.; Lin, C.W. Rhein induces apoptosis in human breast cancer cells. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ip, S.W.; Weng, Y.S.; Lin, S.Y.; Mei, D.; Tang, N.Y.; Su, C.C.; Chung, J.G. The role of Ca+2 on rhein-induced apoptosis in human cervical cancer Ca Ski cells. Anticancer Res. 2007, 27, 379–389. [Google Scholar] [PubMed]
- Li, Y.; Xu, Y.; Lei, B.; Wang, W.; Ge, X.; Li, J. Rhein induces apoptosis of human gastric cancer SGC-7901 cells via an intrinsic mitochondrial pathway. Braz. J. Med. Biol. Res. 2012, 45, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Huang, Z.; Chen, G. Rhein induces apoptosis and cell cycle arrest in human hepatocellular carcinoma BEL-7402 cells. Am. J. Chin. Med. 2008, 36, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Bounda, G.A.; Zhou, W.; Wang, D.D.; Yu, F. Rhein elicits in vitro cytotoxicity in primary human liver hl-7702 cells by inducing apoptosis through mitochondria-mediated pathway. Evid. Based Complement. Altern. Med. 2015, 2015, 329831. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Hazardous effects of sanguinarine on maturation of mouse oocytes, fertilization, and fetal development through apoptotic processes. Environ. Toxicol. 2015, 30, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Cytotoxic effects of dillapiole on embryonic development of mouse blastocysts in vitro and in vivo. Int. J. Mol. Sci. 2014, 15, 10751–10765. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Embryonic toxicity of sanguinarine through apoptotic processes in mouse blastocysts. Toxicol. Lett. 2011, 205, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Huang, F.J.; Chan, W.H. Emodin induces embryonic toxicity in mouse blastocysts through apoptosis. Toxicology 2012, 299, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Citrinin induces apoptosis via a mitochondria-dependent pathway and inhibition of survival signals in embryonic stem cells, and causes developmental injury in blastocysts. Biochem. J. 2007, 404, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Ginkgolide B induces apoptosis and developmental injury in mouse embryonic stem cells and blastocysts. Hum. Reprod. 2006, 21, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K. Cell death in the mammalian blastocyst. Mol. Hum. Reprod. 1997, 3, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.; Stark, J.; Winston, R.M. Maintenance of the inner cell mass in human blastocysts from fragmented embryos. Biol. Reprod. 2003, 68, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Byrne, A.T.; Southgate, J.; Brison, D.R.; Leese, H.J. Analysis of apoptosis in the preimplantation bovine embryo using TUNEL. J. Reprod. Fertil. 1999, 117, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Effects of citrinin on maturation of mouse oocytes, fertilization, and fetal development in vitro and in vivo. Toxicol. Lett. 2008, 180, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Cytotoxic effects of 2-bromopropane on embryonic development in mouse blastocysts. Int. J. Mol. Sci. 2010, 11, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chen, Y.; Yu, R.; Zhao, X.; Wang, Q.; Cai, L. Pretilachlor has the potential to induce endocrine disruption, oxidative stress, apoptosis and immunotoxicity during zebrafish embryo development. Environ. Toxicol. Pharmacol. 2016, 42, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wu, S.; Liu, X.; Wang, Y.; An, X.; Cai, L.; Zhao, X. Effect of acetochlor on transcription of genes associated with oxidative stress, apoptosis, immunotoxicity and endocrine disruption in the early life stage of zebrafish. Environ. Toxicol. Pharmacol. 2015, 40, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Niu, L.; Liu, W.; Xu, C. Embryonic exposure to butachlor in zebrafish (Danio rerio): Endocrine disruption, developmental toxicity and immunotoxicity. Ecotoxicol. Environ. Saf. 2013, 89, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Murugesan, P.; Kanagaraj, P.; Yuvaraj, S.; Balasubramanian, K.; Aruldhas, M.M.; Arunakaran, J. The inhibitory effects of polychlorinated biphenyl Aroclor 1254 on Leydig cell LH receptors, steroidogenic enzymes and antioxidant enzymes in adult rats. Reprod. Toxicol. 2005, 20, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; Hsuuw, Y.D.; Lan, K.C.; Kang, H.Y.; Chang, S.Y.; Hsu, Y.C.; Huang, K.E. Adverse effects of retinoic acid on embryo development and the selective expression of retinoic acid receptors in mouse blastocysts. Hum. Reprod. 2006, 21, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Impact of genistein on maturation of mouse oocytes, fertilization, and fetal development. Reprod. Toxicol. 2009, 28, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Shiao, N.H. Cytotoxic effect of CdSe quantum dots on mouse embryonic development. Acta Pharmacol. Sin. 2008, 29, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Shiao, N.H. Effect of citrinin on mouse embryonic development in vitro and in vivo. Reprod. Toxicol. 2007, 24, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Chang, Y.J. Dosage effects of resveratrol on ethanol-induced cell death in the human K562 cell line. Toxicol. Lett. 2006, 161, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Resveratrol protects against 2-bromopropane-induced apoptosis and disruption of embryonic development in blastocysts. Int. J. Mol. Sci. 2011, 12, 4991–5010. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Wu, H.J. Protective effects of curcumin on methylglyoxal-induced oxidative DNA damage and cell injury in human mononuclear cells. Acta Pharmacol. Sin. 2006, 27, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Wu, H.J.; Hsuuw, Y.D. Curcumin inhibits ROS formation and apoptosis in methylglyoxal-treated human hepatoma G2 cells. Ann. N. Y. Acad. Sci. 2005, 1042, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Senathilake, K.S.; Karunanayake, E.H.; Samarakoon, S.R.; Tennekoon, K.H.; de Silva, E.D.; Adhikari, A. Oleanolic acid from antifilarial triterpene saponins of dipterocarpus zeylanicus induces oxidative stress and apoptosis in filarial parasite Setaria digitata in vitro. Exp. Parasitol. 2017, 177, 13–21. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Feng, C.; Vinothkumar, R.; Chen, W.; Dai, X.; Chen, X.; Ye, Q.; Qiu, C.; Zhou, H.; Wang, Y.; et al. Curcumin analog EF24 induces apoptosis via ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Cancer Chemother. Pharmacol. 2016, 78, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Bian, X.L.; Xu, X.L.; Zhu, B.; Yu, B.; Zhai, Q. Role of mitochondrial permeability transition in human hepatocellular carcinoma Hep-G2 cell death induced by rhein. Fitoterapia 2013, 91, 68–73. [Google Scholar] [CrossRef] [PubMed]
- KoraMagazi, A.; Wang, D.; Yousef, B.; Guerram, M.; Yu, F. Rhein triggers apoptosis via induction of endoplasmic reticulum stress, caspase-4 and intracellular calcium in primary human hepatic HL-7702 cells. Biochem. Biophys. Res. Commun. 2016, 473, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.C.; Werb, Z.; Fisher, S.J. Implantation and the placenta: Key pieces of the development puzzle. Science 1994, 266, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Pampfer, S.; de Hertogh, R.; Vanderheyden, I.; Michiels, B.; Vercheval, M. Decreased inner cell mass proportion in blastocysts from diabetic rats. Diabetes 1990, 39, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.M.; Robaire, B.; Hales, B.F. Paternal cyclophosphamide treatment causes postimplantation loss via inner cell mass-specific cell death. Teratology 1992, 45, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H. Ginkgolides induce apoptosis and decrease cell numbers in mouse blastocysts. Biochem. Biophys. Res. Commun. 2005, 338, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and maturation of the immune system in zebrafish, danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- Trede, N.S.; Langenau, D.M.; Traver, D.; Look, A.T.; Zon, L.I. The use of zebrafish to understand immunity. Immunity 2004, 20, 367–379. [Google Scholar] [CrossRef]
- Eder, K.J.; Clifford, M.A.; Hedrick, R.P.; Kohler, H.R.; Werner, I. Expression of immune-regulatory genes in juvenile Chinook salmon following exposure to pesticides and infectious hematopoietic necrosis virus (IHNV). Fish Shellfish Immunol. 2008, 25, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wu, S.; Wu, C.; An, X.; Cai, L.; Zhao, X. Embryonic exposure to carbendazim induces the transcription of genes related to apoptosis, immunotoxicity and endocrine disruption in zebrafish (Danio rerio). Fish Shellfish Immunol. 2014, 41, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zheng, S.; Fu, Z. Embryonic exposure to cypermethrin induces apoptosis and immunotoxicity in zebrafish (Danio rerio). Fish Shellfish Immunol. 2011, 30, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M.; Moser, B.; Clark-Lewis, I. Interleukin-8 and related chemotactic cytokines. Chest 1994, 105, 95S–98S. [Google Scholar] [CrossRef] [PubMed]
- Orrenius, S. Reactive oxygen species in mitochondria-mediated cell death. Drug Metab. Rev. 2007, 39, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Risso-de, F.C.; Orsini, N.; de Sousa, G.; Rahmani, R. Cadmium-induced apoptosis through the mitochondrial pathway in rainbow trout hepatocytes: Involvement of oxidative stress. Aquat. Toxicol. 2004, 69, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.M.; Chang-Liao, W.L.; Chien, C.F.; Lin, L.C.; Tsai, T.H. Effects of polymer molecular weight on relative oral bioavailability of curcumin. Int. J. Nanomedicine 2012, 7, 2957–2966. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.; Handyside, A.H.; Winston, R.M. The human blastocyst: Cell number, death and allocation during late preimplantation development in vitro. Development 1989, 107, 597–604. [Google Scholar] [PubMed]
- Gardner, R.L.; Davies, T.J. Lack of coupling between onset of giant transformation and genome endoreduplication in the mural trophectoderm of the mouse blastocyst. J. Exp. Zool. 1993, 265, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; Wu, T.C.; Tsai, M.Y. Effect of retinoic acid on implantation and post-implantation development of mouse embryos in vitro. Hum. Reprod. 2001, 16, 2171–2176. [Google Scholar] [CrossRef] [PubMed]
- Witschi, E. Characterization of developmental stages. Part II. Rat. In Biology Data Book, 2nd ed.; Federation of American Societies of Experimental Biologies: Bethesda, MD, USA, 1972; pp. 178–180. [Google Scholar]
- Armant, D.R.; Kaplan, H.A.; Lennarz, W.J. Fibronectin and laminin promote in vitro attachment and outgrowth of mouse blastocysts. Dev. Biol. 1986, 116, 519–523. [Google Scholar] [CrossRef]
- Pampfer, S.; Wuu, Y.D.; Vanderheyden, I.; de Hertogh, R. In vitro study of the carry-over effect associated with early diabetic embryopathy in the rat. Diabetologia 1994, 37, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.H.; Shiao, N.H.; Hsuuw, Y.D.; Chan, W.H. Protective effects of resveratrol on ethanol-induced apoptosis in embryonic stem cells and disruption of embryonic development in mouse blastocysts. Toxicology 2007, 242, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.H.; Hester, J.M.; Stone, B.J.; Carrico, K.M.; Spear, B.T.; Fath, -G.A. Nonsurgical embryo transfer device compared with surgery for embryo transfer in mice. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 17–21. [Google Scholar] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-H.; Chan, W.-H. Rhein Induces Oxidative Stress and Apoptosis in Mouse Blastocysts and Has Immunotoxic Effects during Embryonic Development. Int. J. Mol. Sci. 2017, 18, 2018. https://doi.org/10.3390/ijms18092018
Huang C-H, Chan W-H. Rhein Induces Oxidative Stress and Apoptosis in Mouse Blastocysts and Has Immunotoxic Effects during Embryonic Development. International Journal of Molecular Sciences. 2017; 18(9):2018. https://doi.org/10.3390/ijms18092018
Chicago/Turabian StyleHuang, Chien-Hsun, and Wen-Hsiung Chan. 2017. "Rhein Induces Oxidative Stress and Apoptosis in Mouse Blastocysts and Has Immunotoxic Effects during Embryonic Development" International Journal of Molecular Sciences 18, no. 9: 2018. https://doi.org/10.3390/ijms18092018
APA StyleHuang, C.-H., & Chan, W.-H. (2017). Rhein Induces Oxidative Stress and Apoptosis in Mouse Blastocysts and Has Immunotoxic Effects during Embryonic Development. International Journal of Molecular Sciences, 18(9), 2018. https://doi.org/10.3390/ijms18092018