
Calcium Signaling in Interstitial Cells: Focus on Telocytes

 ,
,  ,
, 

Abstract
:1. Introduction
1.1. Definition and Nomenclature for Interstitial Cells
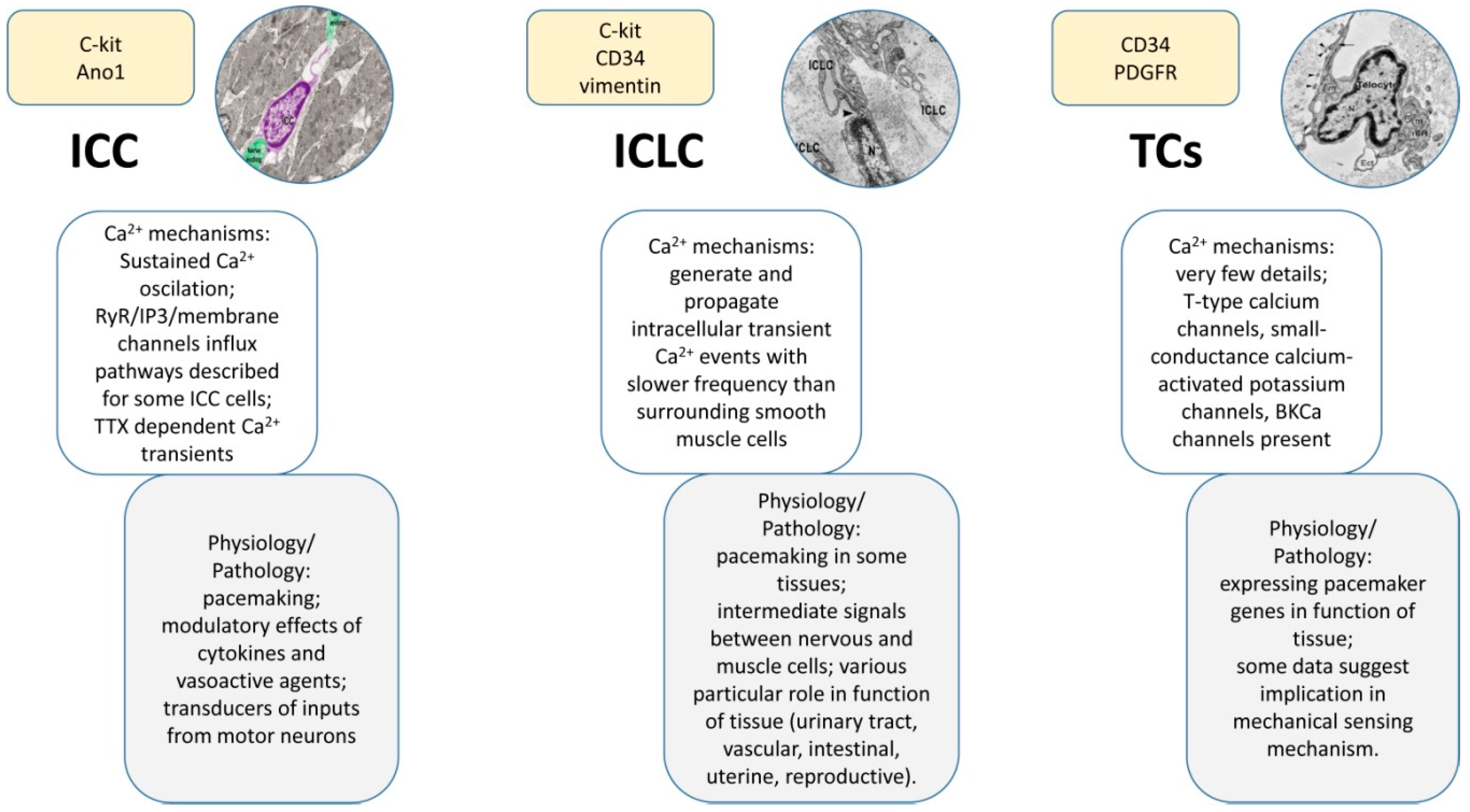
1.2. Interstitial Cells of Cajal (ICCs)
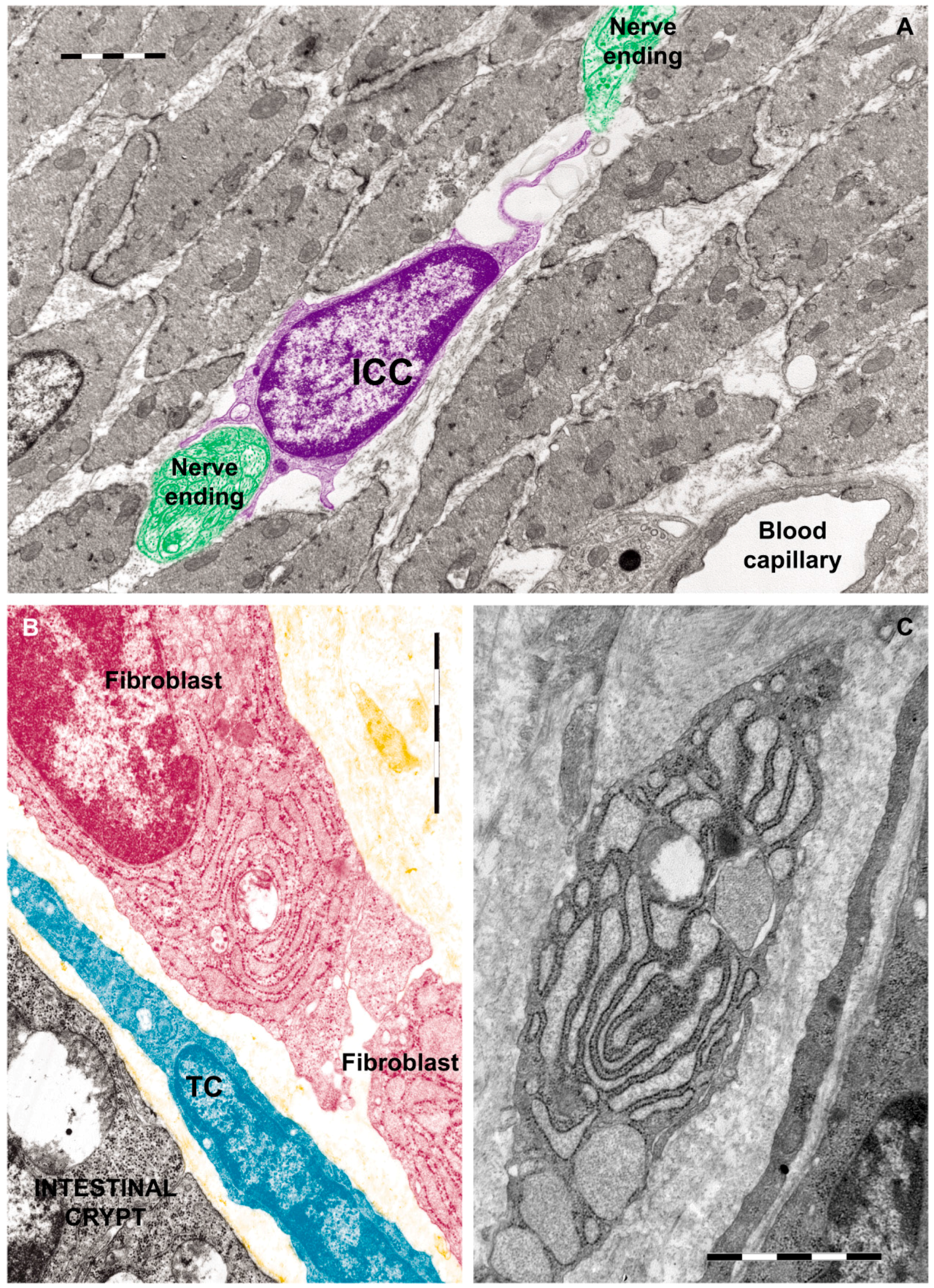
1.2.1. Ultrastructural Features of ICCs
1.2.2. Immunophenotype of ICCs
1.2.3. Roles of ICCs
1.3. Interstitial Cajal-Like Cells (ICLCs)
1.4. TCs
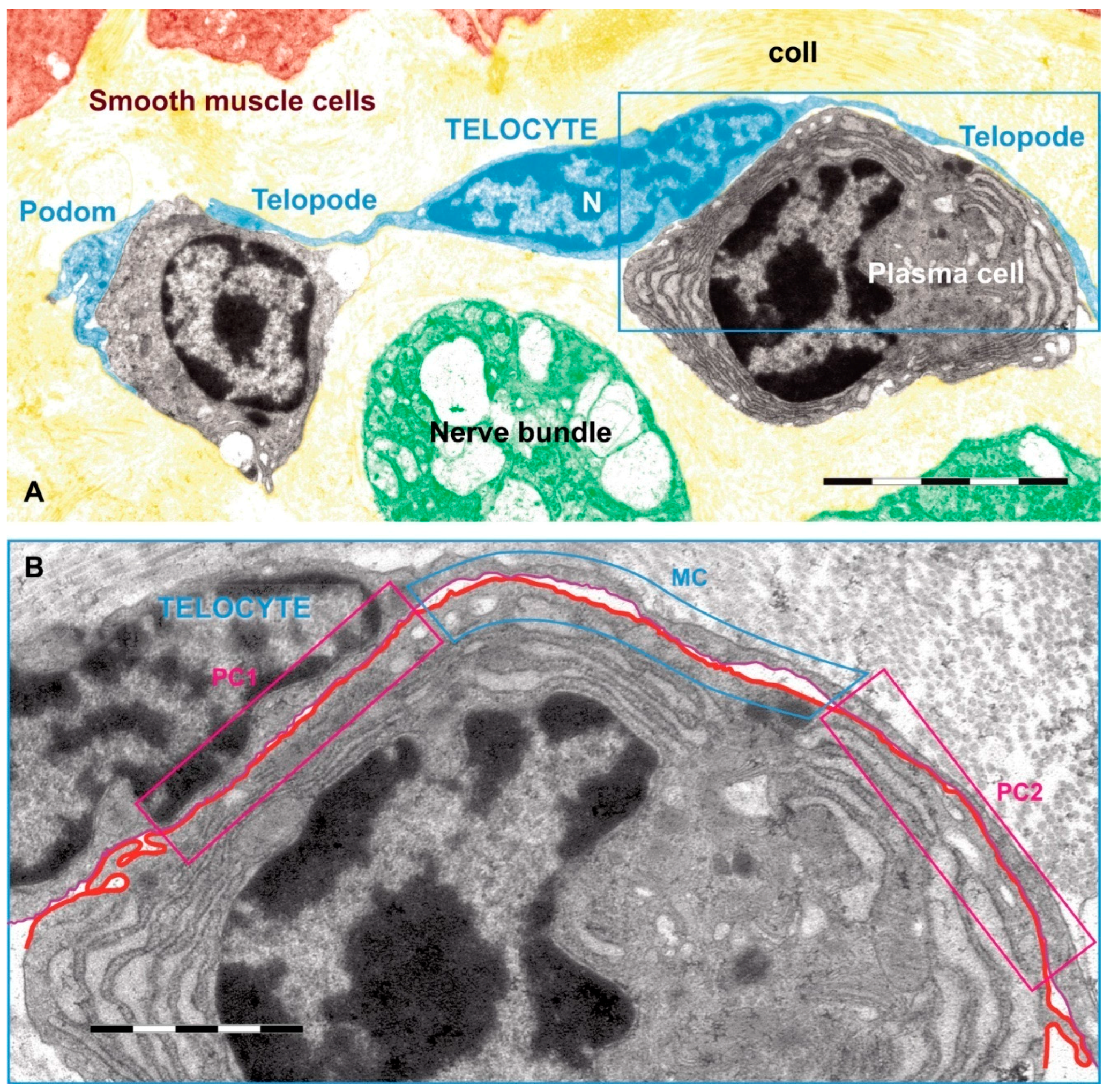
1.4.1. Discovery, Definition and Ultrastructural Features
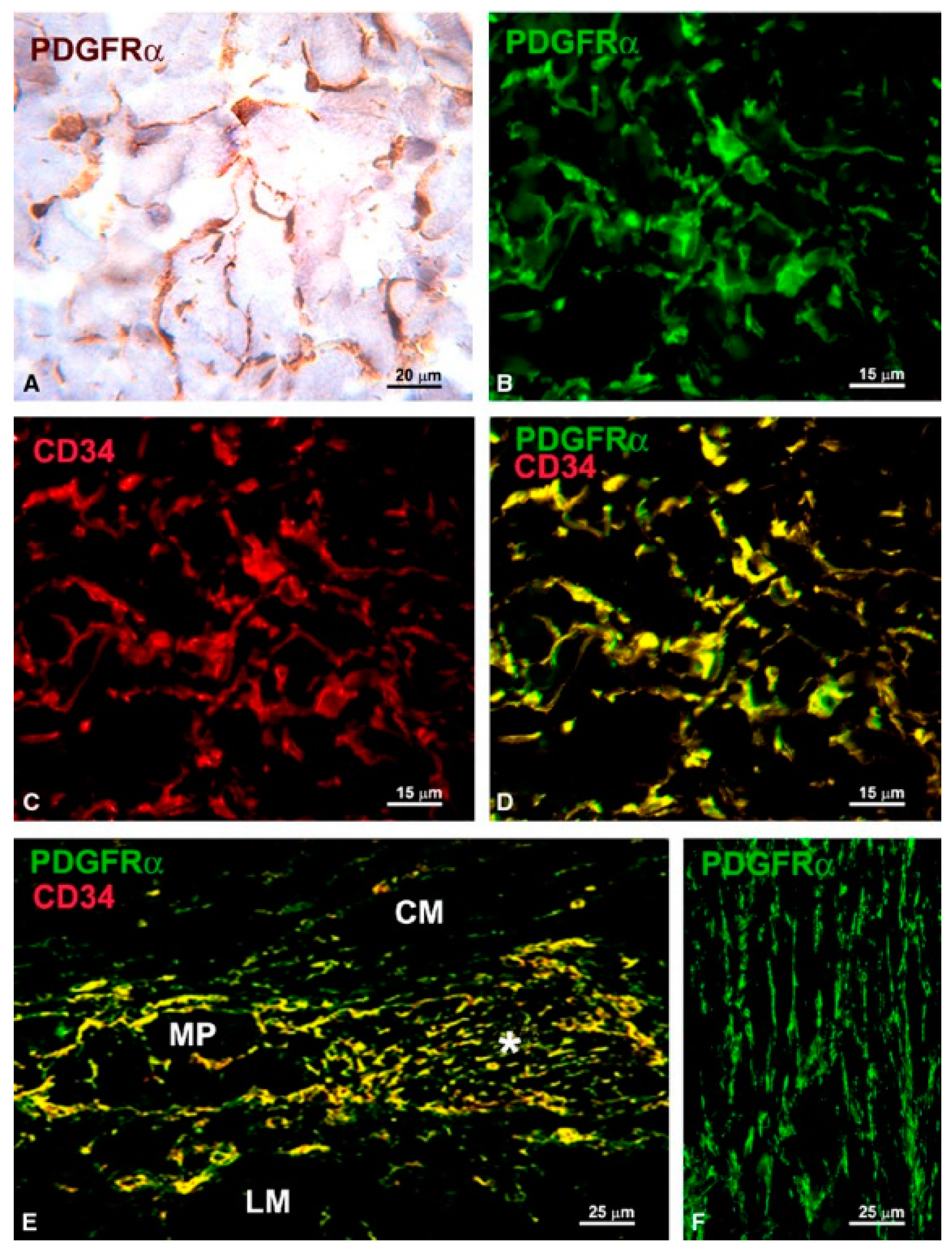
1.4.2. Immunophenotype of TCs
1.4.3. Roles of TCs
2. Calcium Signaling in Interstitial Cells
2.1. Main Calcium Signaling Pathways in Interstitial Cells
2.2. Ca2+ Oscillations in Interstitial Cells
2.3. Cytokines and Vasoactive agents Modulate Calcium Signaling in Interstitial Cells
3. Physiology and Alterations of Calcium Signaling in Interstitial Cells
3.1. Physiological Role of Calcium Signaling in Interstitial Cells
3.1.1. Calcium Signaling in Gastrointestinal Interstitial Cells
3.1.2. Calcium Signaling in the Interstitial Cells of the Urinary Tract
3.1.3. Calcium Signaling in Interstitial Cells of the Female Reproductive System
3.1.4. Calcium Signaling in Interstitial Cells of the Male Reproductive System
3.1.5. Calcium Signaling in Interstitial Cells from the Vascular System
3.2. Pathological Role of Calcium Signaling in Interstitial Cells
3.2.1. Modulation of Calcium Signaling Pathways in Interstitial Cells as a Therapeutic Strategy against Aortic Valve Calcification and Aortic Stenosis
3.2.2. Interstitial Cells Dysfunction during Intestinal Inflammation
4. Calcium Signaling in TCs
4.1. Physiological Role of Calcium Signaling in TCs
4.1.1. Contribution of Calcium Signaling in TCs to the Uterine Physiology
4.1.2. Contribution of Calcium Signaling in TCs to the Cardiac Physiology
4.1.3. Contribution of Calcium Signaling in TCs to the Urinary Physiology
4.2. Pathological Role of Calcium Signaling in TCs
4.2.1. Contribution of Calcium Signaling in TCs to the Immune Response
4.2.2. Correlations between Elements that Modulate TCs Migration and Various Pathologies
4.2.3. TCs and Possible Roles of Calcium Metabolism in Uterine Pathologies
4.2.4. TCs and Possible Roles of Calcium Metabolism in Cardiac Pathologies
5. Concluding Remarks and Perspectives
Acknowledgments
Conflicts of Interest
References
- Vannucchi, M.G.; Traini, C. Interstitial cells of cajal and telocytes in the gut: Twins, related or simply neighbor cells? Biomol. Concepts 2016, 7, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Mongkoldhumrongkul, N.; Yacoub, M.; Chester, A. Valve endothelial cells—Not just any old endothelial cells. Curr. Vasc. Pharmacol. 2016, 14, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, B.; Le Hir, M. The renal cortical interstitium: Morphological and functional aspects. Histochem. Cell Biol. 2008, 130, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Landreh, L.; Weisser, J.; Izzo, G.; Colon, E.; Svechnikova, I.; Soder, O. Origin, development and regulation of human leydig cells. Horm. Res. Paediatr. 2010, 73, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Komuro, T.; Tokui, K.; Zhou, D.S. Identification of the interstitial cells of cajal. Histol. Histopathol. 1996, 11, 769–786. [Google Scholar] [PubMed]
- Cajal, S.R. Les preuves objectives de lúnite anatomiquedes cellules nerveuses. Trav. Lab. Rech. Biol. Univ. Madrid. 1934, 29, 1–137. [Google Scholar]
- Thuneberg, L. One hundred years of interstitial cells of cajal. Microsc. Res. Tech. 1999, 47, 223–238. [Google Scholar] [CrossRef]
- Faussone, P.M.; Cortesini, C.; Romagnoli, P. Ultrastructure of the tunica muscularis of the cardial portion of the human esophagus and stomach, with special reference to the so-called cajal’s interstitial cells. Arch. Ital. Anat. Embriol. 1977, 82, 157–177. [Google Scholar]
- Faussone-Pellegrini, M.S.; Thuneberg, L. Guide to the identification of interstitial cells of cajal. Microsc. Res. Tech. 1999, 47, 248–266. [Google Scholar] [CrossRef]
- Komuro, T. Comparative morphology of interstitial cells of cajal: Ultrastructural characterization. Microsc. Res. Tech. 1999, 47, 267–285. [Google Scholar] [CrossRef]
- Christensen, J. A commentary on the morphological identification of interstitial cells of Cajal in the gut. J. Auton. Nerv. Syst. 1992, 37, 75–88. [Google Scholar] [CrossRef]
- Epperson, A.; Hatton, W.J.; Callaghan, B.; Doherty, P.; Walker, R.L.; Sanders, K.M.; Ward, S.M.; Horowitz, B. Molecular markers expressed in cultured and freshly isolated interstitial cells of cajal. Am. J. Physiol. Cell Physiol. 2000, 279, C529–C539. [Google Scholar] [PubMed]
- Grider, J.R. Focus on “Molecular markers expressed in cultured and freshly isolated interstitial cells of Cajal”. Am. J. Physiol. Cell Physiol. 2000, 279, C284–C285. [Google Scholar] [PubMed]
- Takeda, M.; Takayama, I.; Terada, N.; Baba, T.; Ward, S.M.; Ohno, S.; Fujino, M.A. Immunoelectron-microscopic study of Kit-expressing cells in the jejunum of wildtype and Ws/Ws rats. Cell Tissue Res. 2001, 304, 21–30. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, K.; Nojyo, Y. Wsh/Wsh c-Kit mutant mice possess interstitial cells of Cajal in the deep muscular plexus layer of the small intestine. Neurosci. Lett. 2009, 459, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ordog, T.; Chen, J.; Young, D.L.; Bardsley, M.R.; Redelman, D.; Ward, S.M.; Sanders, K.M. Differential gene expression in functional classes of interstitial cells of Cajal in murine small intestine. Physiol. Genom. 2007, 31, 492–509. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, P.J.; Gibbons, S.J.; Bardsley, M.R.; Lorincz, A.; Pozo, M.J.; Pasricha, P.J.; van de Rijn, M.; West, R.B.; Sarr, M.G.; Kendrick, M.L.; et al. Ano1 is a selective marker of interstitial cells of cajal in the human and mouse gastrointestinal tract. Am. J. Physiol. 2009, 296, G1370–G1381. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M. A case for interstitial cells of cajal as pacemakers and mediators of neurotransmission in the gastrointestinal tract. Gastroenterology 1996, 111, 492–515. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.M.; Ordog, T.; Koh, S.D.; Baker, S.A.; Jun, J.Y.; Amberg, G.; Monaghan, K.; Sanders, K.M. Pacemaking in interstitial cells of cajal depends upon calcium handling by endoplasmic reticulum and mitochondria. J. Physiol. 2000, 525, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Hirst, G.D.; Bramich, N.J.; Teramoto, N.; Suzuki, H.; Edwards, F.R. Regenerative component of slow waves in the guinea-pig gastric antrum involves a delayed increase in [Ca2+]i and Cl− channels. J. Physiol. 2002, 540, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Edwards, F.R.; Hirst, G.D. An electrical analysis of slow wave propagation in the guinea-pig gastric antrum. J. Physiol. 2006, 571, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Pezzone, M.A.; Watkins, S.C.; Alber, S.M.; King, W.E.; de Groat, W.C.; Chancellor, M.B.; Fraser, M.O. Identification of c-Kit-positive cells in the mouse ureter: The interstitial cells of Cajal of the urinary tract. Am. J. Physiol. 2003, 284, F925–F929. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A.; El-Sibai, O.; Shafik, A.A.; Shafik, I. Identification of interstitial cells of Cajal in human urinary bladder: Concept of vesical pacemaker. Urology 2004, 64, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Turunc, T.; Bayazit, Y.; Doran, F.; Bal, N.; Doran, S. Effects of vas deferens obstruction on cajal-like cells in rats. Urol. Int. 2009, 83, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.J.; Klemm, M.F. Interstitial cell of cajal-like cells in the upper urinary tract. J. Cell. Mol. Med. 2005, 9, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.J.; Tonta, M.A.; Zoltkowski, B.Z.; Meeker, W.F.; Wendt, I.; Parkington, H.C. Pyeloureteric peristalsis: Role of atypical smooth muscle cells and interstitial cells of cajal-like cells as pacemakers. J. Physiol. 2006, 576, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Arena, S.; Nicotina, P.A.; Mucciardi, G.; Gali, A.; Magno, C. Pacemakers in the upper urinary tract. Neurourol. Urodyn. 2013, 32, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Feeney, M.M.; Rosenblum, N.D. Urinary tract pacemaker cells: Current knowledge and insights from nonrenal pacemaker cells provide a basis for future discovery. Pediatr. Nephrol. 2014, 29, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Ciontea, S.M.; Radu, E.; Regalia, T.; Ceafalan, L.; Cretoiu, D.; Gherghiceanu, M.; Braga, R.I.; Malincenco, M.; Zagrean, L.; Hinescu, M.E.; et al. C-kit immunopositive interstitial cells (cajal-type) in human myometrium. J. Cell. Mol. Med. 2005, 9, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Hinescu, M.E.; Ionescu, N.; Ciontea, S.M.; Cretoiu, D.; Ardelean, C. Interstitial cells of cajal in pancreas. J. Cell. Mol. Med. 2005, 9, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A.; El-Sibai, O.; Shafik, I.; Shafik, A.A. Immunohistochemical identification of the pacemaker cajal cells in the normal human vagina. Arch. Gynecol. Obstetr. 2005, 272, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Ciontea, S.M.; Cretoiu, D.; Hinescu, M.E.; Radu, E.; Ionescu, N.; Ceausu, M.; Gherghiceanu, M.; Braga, R.I.; Vasilescu, F.; et al. Novel type of interstitial cell (cajal-like) in human fallopian tube. J. Cell. Mol. Med. 2005, 9, 479–523. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A.; Shafik, I.; el-Sibai, O. Identification of c-kit-positive cells in the human prostate: The interstitial cells of cajal. Arch. Androl. 2005, 51, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Radu, E.; Regalia, T.; Ceafalan, L.; Andrei, F.; Cretoiu, D.; Popescu, L.M. Cajal-type cells from human mammary gland stroma: Phenotype characteristics in cell culture. J. Cell. Mol. Med. 2005, 9, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Pucovsky, V.; Harhun, M.I.; Povstyan, O.V.; Gordienko, D.V.; Moss, R.F.; Bolton, T.B. Close relation of arterial icc-like cells to the contractile phenotype of vascular smooth muscle cell. J. Cell. Mol. Med. 2007, 11, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Flores, L.; Gutierrez, R.; Garcia, M.P.; Saez, F.J.; Diaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. Cd34+ stromal cells/fibroblasts/fibrocytes/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role in pathology. Histol. Histopathol. 2014, 29, 831–870. [Google Scholar] [PubMed]
- Diaz-Flores, L.; Gutierrez, R.; Garcia, M.P.; Gonzalez, M.; Saez, F.J.; Aparicio, F.; Diaz-Flores, L., Jr.; Madrid, J.F. Human resident CD34+ stromal cells/telocytes have progenitor capacity and are a source of alphasma+ cells during repair. Histol. Histopathol. 2015, 30, 615–627. [Google Scholar] [PubMed]
- Vannucchi, M.G.; Traini, C.; Guasti, D.; Del Popolo, G.; Faussone-Pellegrini, M.S. Telocytes subtypes in human urinary bladder. J. Cell. Mol. Med. 2014, 18, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express pdgfralpha in the human gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Kito, Y.; Hwang, S.J.; Ward, S.M. Regulation of gastrointestinal smooth muscle function by interstitial cells. Physiology 2016, 31, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from interstitial cells of cajal (icc), via interstitial cajal-like cells (iclc) to telocytes. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, G.; Williams, O.; Cretoiu, D.; Ciontea, S.M. Myometrial interstitial cells and the coordination of myometrial contractility. J. Cell. Mol. Med. 2009, 13, 4268–4282. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Cretoiu, S.M.; Simionescu, A.A.; Popescu, L.M. Telocytes, a distinct type of cell among the stromal cells present in the lamina propria of jejunum. Histol. Histopathol. 2012, 27, 1067–1078. [Google Scholar] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Popescu, L.M. Human myometrium—The ultrastructural 3d network of telocytes. J. Cell. Mol. Med. 2012, 16, 2844–2849. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and their extracellular vesicles-evidence and hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cretoiu, D.; Cretoiu, S.M.; Wang, X. Telocytes and lung disease. Histol. Histopathol. 2016, 31, 1303–1314. [Google Scholar] [PubMed]
- Zheng, M.; Sun, X.; Zhang, M.; Qian, M.; Zheng, Y.; Li, M.; Cretoiu, S.M.; Chen, C.; Chen, L.; Cretoiu, D.; et al. Variations of chromosomes 2 and 3 gene expression profiles among pulmonary telocytes, pneumocytes, airway cells, mesenchymal stem cells and lymphocytes. J. Cell. Mol. Med. 2014, 18, 2044–2060. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zheng, M.; Zhang, M.; Qian, M.; Zheng, Y.; Li, M.; Cretoiu, D.; Chen, C.; Chen, L.; Popescu, L.M.; et al. Differences in the expression of chromosome 1 genes between lung telocytes and other cells: Mesenchymal stem cells, fibroblasts, alveolar type ii cells, airway epithelial cells and lymphocytes. J. Cell. Mol. Med. 2014, 18, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cretoiu, D.; Zheng, M.; Qian, M.; Zhang, M.; Cretoiu, S.M.; Chen, L.; Fang, H.; Popescu, L.M.; Wang, X. Comparison of chromosome 4 gene expression profile between lung telocytes and other local cell types. J. Cell. Mol. Med. 2016, 20, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Cretoiu, D.; Yan, G.; Cretoiu, S.M.; Popescu, L.M.; Fang, H.; Wang, X. Protein profiling of human lung telocytes and microvascular endothelial cells using itraq quantitative proteomics. J. Cell. Mol. Med. 2014, 18, 1035–1059. [Google Scholar] [CrossRef] [PubMed]
- Albulescu, R.; Tanase, C.; Codrici, E.; Popescu, D.I.; Cretoiu, S.M.; Popescu, L.M. The secretome of myocardial telocytes modulates the activity of cardiac stem cells. J. Cell. Mol. Med. 2015, 19, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Cismasiu, V.B.; Radu, E.; Popescu, L.M. Mir-193 expression differentiates telocytes from other stromal cells. J. Cell. Mol. Med. 2011, 15, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, M.; Qian, M.; Wang, L.; Cismasiu, V.B.; Bai, C.; Popescu, L.M.; Wang, X. Genetic comparison of mouse lung telocytes with mesenchymal stem cells and fibroblasts. J. Cell. Mol. Med. 2013, 17, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, S.M. Telocytes heterogeneity: From cellular morphology to functional evidence. Semin. Cell Dev. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Iino, S.; Horiguchi, K. Interstitial cells of cajal are involved in neurotransmission in the gastrointestinal tract. Acta Histochem. Cytochem. 2006, 39, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Lavin, S. Interstitial cells of cajal and purinergic signalling. Auton Neurosci. 2002, 97, 68–72. [Google Scholar] [CrossRef]
- Kim, B.J.; Park, K.J.; Kim, H.W.; Choi, S.; Jun, J.Y.; Chang, I.Y.; Jeon, J.H.; So, I.; Kim, S.J. Identification of trpm7 channels in human intestinal interstitial cells of cajal. World J. Gastroenterol. 2009, 15, 5799–5804. [Google Scholar] [CrossRef] [PubMed]
- Johnston, L.; Sergeant, G.P.; Hollywood, M.A.; Thornbury, K.D.; McHale, N.G. Calcium oscillations in interstitial cells of the rabbit urethra. J. Physiol. 2005, 565, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Calcium signal transduction and cellular control mechanisms. Biochim. Biophys. Acta 2004, 1742, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Inositol trisphosphate and calcium signalling mechanisms. Biochim. Biophys. Acta 2009, 1793, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Smooth muscle cell calcium activation mechanisms. J. Physiol. 2008, 586, 5047–5061. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Koh, S.D.; Ward, S.M. Interstitial cells of cajal as pacemakers in the gastrointestinal tract. Ann. Rev. Physiol. 2006, 68, 307–343. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.A.; Drumm, B.T.; Saur, D.; Hennig, G.W.; Ward, S.M.; Sanders, K.M. Spontaneous Ca(2+) transients in interstitial cells of cajal located within the deep muscular plexus of the murine small intestine. J. Physiol. 2016, 594, 3317–3338. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.H.; Sung, T.S.; O’Driscoll, K.; Koh, S.D.; Sanders, K.M. Intracellular Ca(2+) release from endoplasmic reticulum regulates slow wave currents and pacemaker activity of interstitial cells of cajal. Am. J. Physiol. 2015, 308, C608–C620. [Google Scholar] [CrossRef] [PubMed]
- Beyder, A.; Farrugia, G. Targeting ion channels for the treatment of gastrointestinal motility disorders. Therap. Adv. Gastroenterol. 2012, 5, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Linsdell, P. Evidence that extracellular anions interact with a site outside the cftr chloride channel pore to modify channel properties. Can. J. Physiol. Pharmacol. 2009, 87, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, D.; Ceafalan, L.C.; Popescu, L.M. Isolated human uterine telocytes: Immunocytochemistry and electrophysiology of t-type calcium channels. Histochem. Cell Biol. 2015, 143, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Hashitani, H.; Lang, R.J. Functions of icc-like cells in the urinary tract and male genital organs. J. Cell. Mol. Med. 2010, 14, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.J.; Hashitani, H.; Tonta, M.A.; Bourke, J.L.; Parkington, H.C.; Suzuki, H. Spontaneous electrical and Ca2+ signals in the mouse renal pelvis that drive pyeloureteric peristalsis. Clin. Exp. Pharmacol. Physiol. 2010, 37, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Drumm, B.T.; Koh, S.D.; Andersson, K.E.; Ward, S.M. Calcium signalling in cajal-like interstitial cells of the lower urinary tract. Nat. Rev. Urol. 2014, 11, 555–564. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, K.D.; Gurney, A.M. Kit positive cells in the guinea pig bladder. J. Urol. 2002, 168, 832–836. [Google Scholar] [CrossRef]
- Hashitani, H.; Yanai, Y.; Suzuki, H. Role of interstitial cells and gap junctions in the transmission of spontaneous Ca2+ signals in detrusor smooth muscles of the guinea-pig urinary bladder. J. Physiol. 2004, 559, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, G.P.; Hollywood, M.A.; McHale, N.G.; Thornbury, K.D. Ca2+ signalling in urethral interstitial cells of cajal. J. Physiol. 2006, 576, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.; Shigemasa, Y.; Exintaris, B.; Lang, R.J.; Hashitani, H. Spontaneous Ca2+ signaling of interstitial cells in the guinea pig prostate. J. Urol. 2011, 186, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; He, P.; Zhong, X.; Wang, Q.; Li, L.; Song, B. Identification of t-type calcium channels in the interstitial cells of cajal in rat bladder. Urology 2012, 80, e1381–e1387. [Google Scholar] [CrossRef] [PubMed]
- Nivala, M.; Ko, C.Y.; Nivala, M.; Weiss, J.N.; Qu, Z. The emergence of subcellular pacemaker sites for calcium waves and oscillations. J. Physiol. 2013, 591, 5305–5320. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.; Wellman, G.C. Setting the pace for gi motility: Ryanodine receptors and ip3 receptors within interstitial cells of cajal. Focus on “intracellular Ca2+ release from endoplasmic reticulum regulates slow wave currents and pacemaker activity of interstitial cells of cajal”. Am. J. Physiol. 2015, 308, C606–C607. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.D.; Gibbons, S.J.; Saravanaperumal, S.A.; Du, P.; Hennig, G.W.; Eisenman, S.T.; Mazzone, A.; Hayashi, Y.; Cao, C.; Stoltz, G.J.; et al. Ano1, a Ca2+-activated Cl-channel, coordinates contractility in mouse intestine by Ca2+ transient coordination between interstitial cells of cajal. J. Physiol. 2014, 592, 4051–4068. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Huang, L.; Cheng, W.; Li, X.; Lu, J.; Lin, L.; Si, X. Roles of interleukin-9 in the growth and cholecystokinin-induced intracellular calcium signaling of cultured interstitial cells of cajal. PLoS ONE 2014, 9, e95898. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhu, Y.; Khan, W.I.; Van Snick, J.; Huizinga, J.D. Il-9 enhances growth of icc, maintains network structure and strengthens rhythmicity of contraction in culture. J. Cell. Mol. Med. 2006, 10, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Zarate, N.; Soderholm, J.D.; Bourgeois, J.M.; Liu, L.W.; Huizinga, J.D. Ultrastructural injury to interstitial cells of cajal and communication with mast cells in crohn’s disease. Neurogastroenterol. Motility 2007, 19, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Fullerton, D.A.; Ao, L.; Zheng, D.; Zhao, K.S.; Meng, X. Bmp-2 and TGF-beta1 mediate biglycan-induced pro-osteogenic reprogramming in aortic valve interstitial cells. J. Mol. Med. 2015, 93, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Seya, K.; Daitoku, K.; Motomura, S.; Fukuda, I.; Furukawa, K. Tumor necrosis factor-alpha accelerates the calcification of human aortic valve interstitial cells obtained from patients with calcific aortic valve stenosis via the bmp2-dlx5 pathway. J. Pharmacol. Exp. Therap. 2011, 337, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Hafizi, S.; Taylor, P.M.; Chester, A.H.; Allen, S.P.; Yacoub, M.H. Mitogenic and secretory responses of human valve interstitial cells to vasoactive agents. J. Heart Valve Dis. 2000, 9, 454–458. [Google Scholar] [PubMed]
- Choi, S.; Park, D.Y.; Yeum, C.H.; Chang, I.Y.; You, H.J.; Park, C.G.; Kim, M.Y.; Kong, I.D.; So, I.; Kim, K.W.; et al. Bradykinin modulates pacemaker currents through bradykinin b2 receptors in cultured interstitial cells of cajal from the murine small intestine. Br. J. Pharmacol. 2006, 148, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Kwon, Y.K.; Kim, E.; So, I. Effects of histamine on cultured interstitial cells of cajal in murine small intestine. Korean J. Physiol. Pharmacol. 2013, 17, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Rumessen, J.J. Identification of interstitial cells of cajal. Significance for studies of human small intestine and colon. Dan. Med. Bull. 1994, 41, 275–293. [Google Scholar] [PubMed]
- Park, K.J.; Hennig, G.W.; Lee, H.T.; Spencer, N.J.; Ward, S.M.; Smith, T.K.; Sanders, K.M. Spatial and temporal mapping of pacemaker activity in interstitial cells of cajal in mouse ileum in situ. Am. J. Physiol. 2006, 290, C1411–C1427. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Huang, X.; Liu, D.H.; Lu, H.L.; Kim, Y.C.; Xu, W.X. Involvement of actin microfilament in regulation of pacemaking activity increased by hypotonic stress in cultured iccs of murine intestine. Physiol. Res. 2015, 64, 397–405. [Google Scholar] [PubMed]
- Torihashi, S.; Fujimoto, T.; Trost, C.; Nakayama, S. Calcium oscillation linked to pacemaking of interstitial cells of cajal: Requirement of calcium influx and localization of trp4 in caveolae. J. Biol. Chem. 2002, 277, 19191–19197. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, A. Furthering the debate on the role of interstitial cells of cajal in enteric inhibitory neuromuscular neurotransmission. Am. J. Physiol. 2016, 311, C479–C481. [Google Scholar] [CrossRef] [PubMed]
- McHale, N.G.; Hollywood, M.A.; Sergeant, G.P.; Shafei, M.; Thornbury, K.T.; Ward, S.M. Organization and function of icc in the urinary tract. J. Physiol. 2006, 576, 689–694. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, K.D. Interstitial cells of cajal in the urinary tract. Handbook Exp. Pharmacol. 2011, 233–254. [Google Scholar]
- Wu, C.; Sui, G.P.; Fry, C.H. Purinergic regulation of guinea pig suburothelial myofibroblasts. J. Physiol. 2004, 559, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.M.; McGeown, J.G.; McMurray, G.; McCloskey, K.D. Functional innervation of guinea-pig bladder interstitial cells of cajal subtypes: Neurogenic stimulation evokes in situ calcium transients. PLoS ONE 2013, 8, e53423. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.J.; Zoltkowski, B.Z.; Hammer, J.M.; Meeker, W.F.; Wendt, I. Electrical characterization of interstitial cells of cajal-like cells and smooth muscle cells isolated from the mouse ureteropelvic junction. J. Urol. 2007, 177, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, Y.J.; Ciray, N.; Olovsson, M.; Ulmsten, U.; Gylfe, E. Oxytocin-induced oscillations of cytoplasmic Ca2+ in human myometrial cells. Acta Obstet. Gynecol Scand. 2000, 79, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Young, R.C.; Zhang, P. The mechanism of propagation of intracellular calcium waves in cultured human uterine myocytes. Am. J. Obstet. Gynecol. 2001, 184, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Loftus, F.C.; Richardson, M.J.; Shmygol, A. Single-cell mechanics and calcium signalling in organotypic slices of human myometrium. J. Biomech. 2015, 48, 1620–1624. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.E.; Ramsey, K.H.; Schripsema, J.H.; Sanders, K.M.; Ward, S.M. Time-dependent disruption of oviduct pacemaker cells by chlamydia infection in mice. Biol. Reproduct. 2010, 83, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Matzkin, M.E.; Lauf, S.; Spinnler, K.; Rossi, S.P.; Kohn, F.M.; Kunz, L.; Calandra, R.S.; Frungieri, M.B.; Mayerhofer, A. The Ca2+-activated, large conductance K+-channel (bkca) is a player in the lh/hcg signaling cascade in testicular leydig cells. Mol. Cell. Endocrinol. 2013, 367, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.R.; Reis, R.I.; Aguiar, J.F.; Varanda, W.A. Luteinizing hormone (lh) acts through pka and pkc to modulate t-type calcium currents and intracellular calcium transients in mice leydig cells. Cell Calcium 2011, 49, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, J.U.; Kim, C.; Min, C.K. Inhibitory actions of mibefradil on steroidogenesis in mouse leydig cells: Involvement of Ca(2+) entry via the t-type Ca(2+) channel. Asian J. Androl. 2010, 12, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Pardyak, L.; Kaminska, A.; Galas, J.; Ptak, A.; Bilinska, B.; Kotula-Balak, M. Primary and tumor mouse leydig cells exposed to polychlorinated naphthalenes mixture: Effect on estrogen related-receptors expression, intracellular calcium level and sex hormones secretion. Tissue Cell 2016, 48, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Abdou, H.S.; Villeneuve, G.; Tremblay, J.J. The calcium signaling pathway regulates leydig cell steroidogenesis through a transcriptional cascade involving the nuclear receptor nr4a1 and the steroidogenic acute regulatory protein. Endocrinology 2013, 154, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Abdou, H.S.; Robert, N.M.; Tremblay, J.J. Calcium-dependent nr4a1 expression in mouse leydig cells requires distinct ap1/cre and mef2 elements. J. Mol. Endocrinol. 2016, 56, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.I. Mitochondrial Ca2+ handling is crucial for generation of rhythmical Ca2+ waves in vascular interstitial cells from rabbit portal vein. Cell Calcium 2015, 58, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.I.; Gordienko, D.V.; Povstyan, O.V.; Moss, R.F.; Bolton, T.B. Function of interstitial cells of cajal in the rabbit portal vein. Circ. Res. 2004, 95, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.; Gordienko, D.; Kryshtal, D.; Pucovsky, V.; Bolton, T. Role of intracellular stores in the regulation of rhythmical [Ca2+]i changes in interstitial cells of cajal from rabbit portal vein. Cell Calcium 2006, 40, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Bolton, T.B.; Gordienko, D.V.; Povstyan, O.V.; Harhun, M.I.; Pucovsky, V. Smooth muscle cells and interstitial cells of blood vessels. Cell Calcium 2004, 35, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Pucovsky, V.; Bolton, T.B. Localisation, function and composition of primary Ca2+ spark discharge region in isolated smooth muscle cells from guinea-pig mesenteric arteries. Cell Calcium 2006, 39, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.I.; Pucovsky, V.; Povstyan, O.V.; Gordienko, D.V.; Bolton, T.B. Interstitial cells in the vasculature. J. Cell. Mol. Med. 2005, 9, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.I.; Szewczyk, K.; Laux, H.; Prestwich, S.A.; Gordienko, D.V.; Moss, R.F.; Bolton, T.B. Interstitial cells from rat middle cerebral artery belong to smooth muscle cell type. J. Cell. Mol. Med. 2009, 13, 4532–4539. [Google Scholar] [CrossRef] [PubMed]
- Huggins, C.L.; Povstyan, O.V.; Harhun, M.I. Characterization of transcriptional and posttranscriptional properties of native and cultured phenotypically modulated vascular smooth muscle cells. Cell Tissue Res. 2013, 352, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Harhun, M.I.; Huggins, C.L.; Ratnasingham, K.; Raje, D.; Moss, R.F.; Szewczyk, K.; Vasilikostas, G.; Greenwood, I.A.; Khong, T.K.; Wan, A.; et al. Resident phenotypically modulated vascular smooth muscle cells in healthy human arteries. J. Cell. Mol. Med. 2012, 16, 2802–2812. [Google Scholar] [CrossRef] [PubMed]
- Gordienko, D.V.; Greenwood, I.A.; Bolton, T.B. Direct visualization of sarcoplasmic reticulum regions discharging Ca(2+)sparks in vascular myocytes. Cell Calcium 2001, 29, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhao, R.; Yang, Y.; Wang, H.; Shao, Y.; Kong, X. Comparative study of human aortic and mitral valve interstitial cell gene expression and cellular function. Genomics 2013, 101, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Rabkin-Aikawa, E.; Farber, M.; Aikawa, M.; Schoen, F.J. Dynamic and reversible changes of interstitial cell phenotype during remodeling of cardiac valves. J. Heart Valve Dis. 2004, 13, 841–847. [Google Scholar] [PubMed]
- Masjedi, S.; Amarnath, A.; Baily, K.M.; Ferdous, Z. Comparison of calcification potential of valvular interstitial cells isolated from individual aortic valve cusps. Cardiovasc. Pathol. 2016, 25, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Thuneberg, L.; Kluppel, M.; Malysz, J.; Mikkelsen, H.B.; Bernstein, A. W/kit gene required for interstitial cells of cajal and for intestinal pacemaker activity. Nature 1995, 373, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Marin, A.; Radu, B.M.; Popescu, L.M. Telocytes: Ultrastructural, immunohistochemical and electrophysiological characteristics in human myometrium. Reproduction 2013, 145, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Campeanu, R.A.; Radu, B.M.; Cretoiu, S.M.; Banciu, D.D.; Banciu, A.; Cretoiu, D.; Popescu, L.M. Near-infrared low-level laser stimulation of telocytes from human myometrium. Lasers Med. Sci. 2014, 29, 1867–1874. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Shim, W.; Lu, J.; Lim, S.Y.; Ong, B.H.; Lim, T.S.; Liew, R.; Chua, Y.L.; Wong, P. Electrophysiology of human cardiac atrial and ventricular telocytes. J. Cell. Mol. Med. 2014, 18, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.S.; Meng, X.; Song, R.; Fullerton, D.; Jaggers, J. Rapamycin decreases the osteogenic response in aortic valve interstitial cells through the stat3 pathway. Ann. Thoracic Surg. 2016, 102, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Helas, S.; Goettsch, C.; Schoppet, M.; Zeitz, U.; Hempel, U.; Morawietz, H.; Kostenuik, P.J.; Erben, R.G.; Hofbauer, L.C. Inhibition of receptor activator of NF-κB ligand by denosumab attenuates vascular calcium deposition in mice. Am. J. Pathol. 2009, 175, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Lerman, D.A.; Prasad, S.; Alotti, N. Denosumab could be a potential inhibitor of valvular interstitial cells calcification in vitro. Int. J. Cardiovasc. Res. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Der, T.; Bercik, P.; Donnelly, G.; Jackson, T.; Berezin, I.; Collins, S.M.; Huizinga, J.D. Interstitial cells of cajal and inflammation-induced motor dysfunction in the mouse small intestine. Gastroenterology 2000, 119, 1590–1599. [Google Scholar] [CrossRef] [PubMed]
- Kaji, N.; Horiguchi, K.; Iino, S.; Nakayama, S.; Ohwada, T.; Otani, Y.; Firman; Murata, T.; Sanders, K.M.; Ozaki, H.; et al. Nitric oxide-induced oxidative stress impairs pacemaker function of murine interstitial cells of cajal during inflammation. Pharmacol. Res. 2016, 111, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Cretoiu, S.M. Telocytes in the reproductive organs: Current understanding and future challenges. Semin. Cell Dev. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Peri, L.E.; Koh, B.H.; Ward, G.K.; Bayguinov, Y.; Hwang, S.J.; Gould, T.W.; Mullan, C.J.; Sanders, K.M.; Ward, S.M. A novel class of interstitial cells in the mouse and monkey female reproductive tracts. Biol. Reproduct. 2015, 92, 102. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.; Gomez-Pinilla, P.J.; Pozo, M.J.; Cima, R.R.; Dozois, E.J.; Larson, D.W.; Ordog, T.; Gibbons, S.J.; Farrugia, G. Immunoreactivity for ano1 detects depletion of kit-positive interstitial cells of cajal in patients with slow transit constipation. Neurogastroenterol. Motil. 2011, 23, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wei, L.; Zhong, C.; Fu, S.; Bei, Y.; Huica, R.I.; Wang, F.; Xiao, J. Cardiac telocytes are double positive for cd34/pdgfr-alpha. J. Cell. Mol. Med. 2015, 19, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zhang, S.; Li, Y.G.; Wang, Y. Isolation, culture, purification and ultrastructural investigation of cardiac telocytes. Mol. Med. Rep. 2016, 14, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Shynlova, O.; Tsui, P.; Jaffer, S.; Lye, S.J. Integration of endocrine and mechanical signals in the regulation of myometrial functions during pregnancy and labour. Eur. J. Obs. Gynecol. Reproduct. Biol. 2009, 144, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Roatesi, I.; Radu, B.M.; Cretoiu, D.; Cretoiu, S.M. Uterine telocytes: A review of current knowledge. Biol. Reproduct. 2015, 93, 10. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, S.T.; Svalo, J.; Nielsen, K.; Larsen, T.; Jorgensen, J.C.; Bouchelouche, P. Immunolocalization and expression of small-conductance calcium-activated potassium channels in human myometrium. J. Cell. Mol. Med. 2012, 16, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, S.T.; Larsen, T.; Joergensen, J.C.; Bouchelouche, P.N. Relaxant effect of a novel calcium-activated potassium channel modulator on human myometrial spontaneous contractility in vitro. Acta Physiol. 2012, 205, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Li, C.; Lu, Z.; Li, H.; Guo, Z. Multiple immunophenotypes of cardiac telocytes. Exp. Cell Res. 2015, 338, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Bei, Y.; Zhou, Q.; Fu, S.; Lv, D.; Chen, P.; Chen, Y.; Wang, F.; Xiao, J. Cardiac telocytes and fibroblasts in primary culture: Different morphologies and immunophenotypes. PLoS ONE 2015, 10, e0115991. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.H.; Wu, Y.; Gao, Z.; Anderson, M.E.; Dalziel, J.E.; Meredith, A.L. Bk channels regulate sinoatrial node firing rate and cardiac pacing in vivo. Am. J. Physiol. 2014, 307, H1327–H1338. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Jiang, X.J.; Su, L.; Shen, Z.J.; Yang, X.J. In vitro morphology, viability and cytokine secretion of uterine telocyte-activated mouse peritoneal macrophages. J. Cell. Mol. Med. 2015, 19, 2741–2750. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Gherghiceanu, M.; Simionescu, O. Telocyte dynamics in psoriasis. J. Cell. Mol. Med. 2015, 19, 1504–1519. [Google Scholar] [CrossRef] [PubMed]
- Karvonen, S.L.; Korkiamaki, T.; Yla-Outinen, H.; Nissinen, M.; Teerikangas, H.; Pummi, K.; Karvonen, J.; Peltonen, J. Psoriasis and altered calcium metabolism: Downregulated capacitative calcium influx and defective calcium-mediated cell signaling in cultured psoriatic keratinocytes. J. Investig. Dermatol. 2000, 114, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Steinckwich, N.; Myers, P.; Janardhan, K.S.; Flagler, N.D.; King, D.; Petranka, J.G.; Putney, J.W. Role of the store-operated calcium entry protein, stim1, in neutrophil chemotaxis and infiltration into a murine model of psoriasis-inflamed skin. FASEB J. 2015, 29, 3003–3013. [Google Scholar] [CrossRef] [PubMed]
- Staniak, H.L.; Bittencourt, M.S.; de Souza Santos, I.; Sharovsky, R.; Sabbag, C.; Goulart, A.C.; Lotufo, P.A.; Bensenor, I.M. Association between psoriasis and coronary calcium score. Atherosclerosis 2014, 237, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Qadim, H.H.; Goforoushan, F.; Nejad, S.B.; Goldust, M. Studying the calcium serum level in patients suffering from psoriasis. Pak. J. Biol. Sci. 2013, 16, 291–294. [Google Scholar] [PubMed]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kreymann, G.; Grosser, S.; Buggisch, P.; Gottschall, C.; Matthaei, S.; Greten, H. Oxygen consumption and resting metabolic rate in sepsis, sepsis syndrome, and septic shock. Crit. Care Med. 1993, 21, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R.; Bacigalupe, L.D.; Schmitt, B.; Hoppeler, H. Allometric scaling of maximal metabolic rate in mammals: Muscle aerobic capacity as determinant factor. Respir. Physiol. Neurobiol. 2004, 140, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Valiani, V.; Corbett, D.B.; Knaggs, J.D.; Manini, T.M. Metabolic rate and perceived exertion of walking in older adults with idiopathic chronic fatigue. J. Gerontol. A 2016, 71, 1444–1450. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.K.; Gayatri, K.; Jammula, S.; Kota, S.K.; Krishna, S.V.; Meher, L.K.; Modi, K.D. Endocrinology of parturition. Ind. J. Endocrinol. Metab. 2013, 17, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Oguri, H.; Yamada, R.; Maeda, N.; Kohsaki, S.; Fukaya, T. Preoperative evaluation of pelvic masses with combined 18f-fluorodeoxyglucose positron emission tomography and computed tomography. Int. J. Gynaecol. Obstet. 2008, 102, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, U.K.; Svensson, S.P. Inhibition of Ca2+/calmodulin-dependent protein kinase or epidermal growth factor receptor tyrosine kinase abolishes lysophosphatidic acid-mediated DNA-synthesis in human myometrial smooth muscle cells. Cell Biol. Int. 2003, 27, 341–347. [Google Scholar] [CrossRef]
- Wimalasundera, R.C.; Thom, S.A.; Regan, L.; Hughes, A.D. Effects of vasoactive agents on intracellular calcium and force in myometrial and subcutaneous resistance arteries isolated from preeclamptic, pregnant, and nonpregnant woman. Am. J. Obstet. Gynecol. 2005, 192, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. The role of cardiac fibroblasts in post-myocardial heart tissue repair. Exp. Mol. Pathol. 2016, 101, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Gherghiceanu, M.; Hinescu, M.E.; Cretoiu, D.; Ceafalan, L.; Regalia, T.; Popescu, A.C.; Ardeleanu, C.; Mandache, E. Insights into the interstitium of ventricular myocardium: Interstitial cajal-like cells (ICLC). J. Cell. Mol. Med. 2006, 10, 429–458. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anatomical Localization of the Interstitial Cells | Subtype of Interstitial Cells | Calcium Signaling Mechanisms | Contribution to the Pacemaking Activity | Reference |
|---|---|---|---|---|
| Gastrointestinal system | ICCs |
| Yes | [63,90] |
| Urinary system | ICCs |
| Yes/No (Depending on the segment of the urinary tract) | [74,93,96] |
| ICLCs |
| Yes | [71,97] | |
| Reproductive system | ICCs |
| Yes | [101] |
| ICLCs |
| Yes | [29,43] | |
| TCs |
| No | [68,121,122,123] | |
| Leydig cells |
| N/A | [102,103,104,106,107] | |
| Cardiovascular system | Vascular interstitial cells |
| Yes | [110,113,117] |
| TCs |
| Probably yes | [124] | |
| Valve interstitial cells |
| N/A | [120] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radu, B.M.; Banciu, A.; Banciu, D.D.; Radu, M.; Cretoiu, D.; Cretoiu, S.M. Calcium Signaling in Interstitial Cells: Focus on Telocytes. Int. J. Mol. Sci. 2017, 18, 397. https://doi.org/10.3390/ijms18020397
Radu BM, Banciu A, Banciu DD, Radu M, Cretoiu D, Cretoiu SM. Calcium Signaling in Interstitial Cells: Focus on Telocytes. International Journal of Molecular Sciences. 2017; 18(2):397. https://doi.org/10.3390/ijms18020397
Chicago/Turabian StyleRadu, Beatrice Mihaela, Adela Banciu, Daniel Dumitru Banciu, Mihai Radu, Dragos Cretoiu, and Sanda Maria Cretoiu. 2017. "Calcium Signaling in Interstitial Cells: Focus on Telocytes" International Journal of Molecular Sciences 18, no. 2: 397. https://doi.org/10.3390/ijms18020397
APA StyleRadu, B. M., Banciu, A., Banciu, D. D., Radu, M., Cretoiu, D., & Cretoiu, S. M. (2017). Calcium Signaling in Interstitial Cells: Focus on Telocytes. International Journal of Molecular Sciences, 18(2), 397. https://doi.org/10.3390/ijms18020397