

Impact of Clove Essential Oil on the Intestinal Flora in Mice Infected with Candida albicans

 , , ,
, , ,
Abstract

1. Introduction
2. Results
2.1. Four Components Were Identified by GC-MS in CEO
2.2. CEO Has an Inhibitory Effect on C. albicans
2.3. Effect of CEO on the Relative Conductivity of C. albicans
2.4. Effect of CEO on Protein Damage in C. albicans
2.5. Effects of CEO on Biofilm Formation
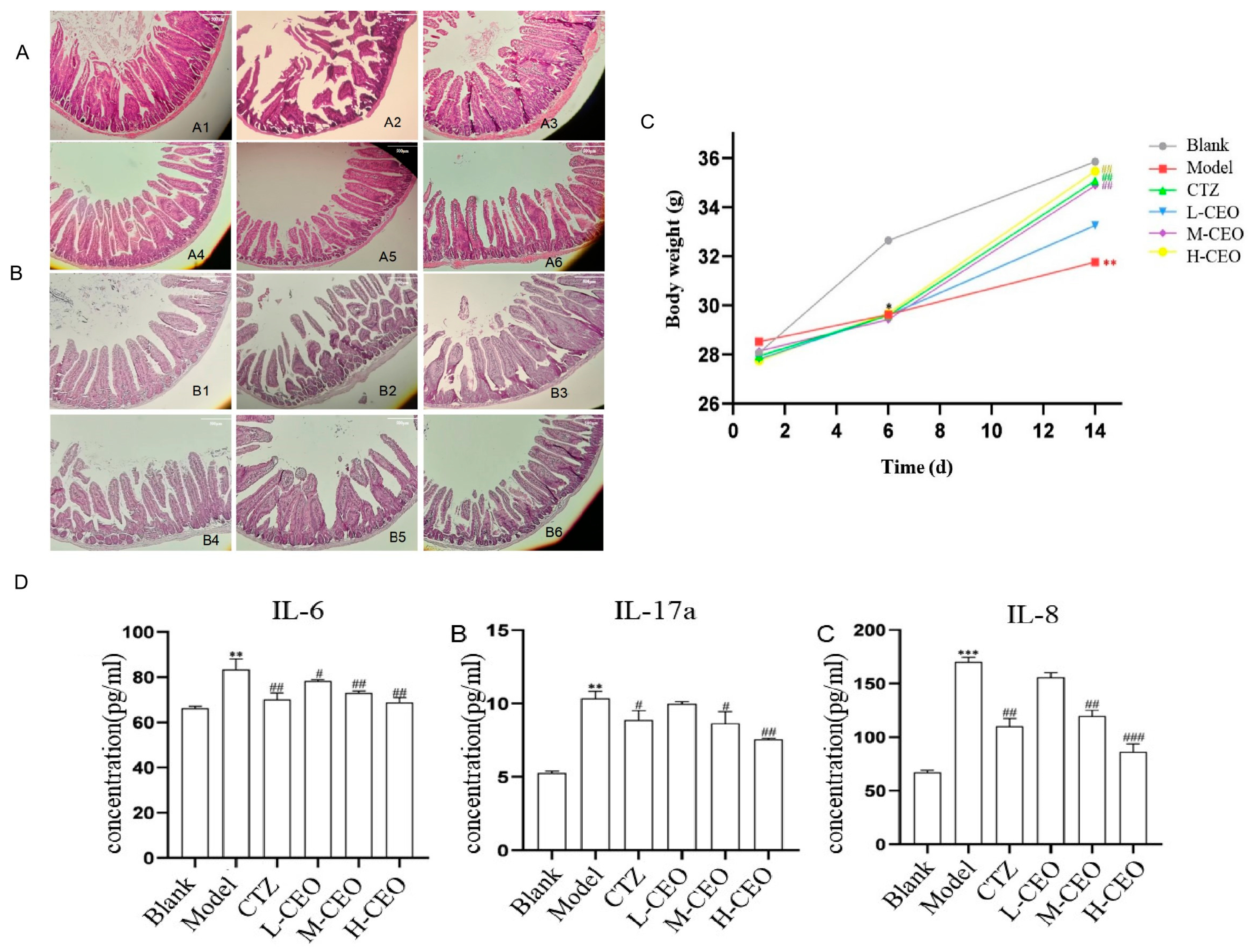
2.6. Effect of CEO on General Signs and Body Weight of Mice
2.7. Effect of CEO on Intestinal Histopathology in Mice
2.8. Effect of CEO on IL-6, IL-17A, and IL-8 in Mouse Intestinal Tissues
2.9. High-Throughput Sequencing of the Mouse Gut Microbiota
2.10. α Diversity Analysis
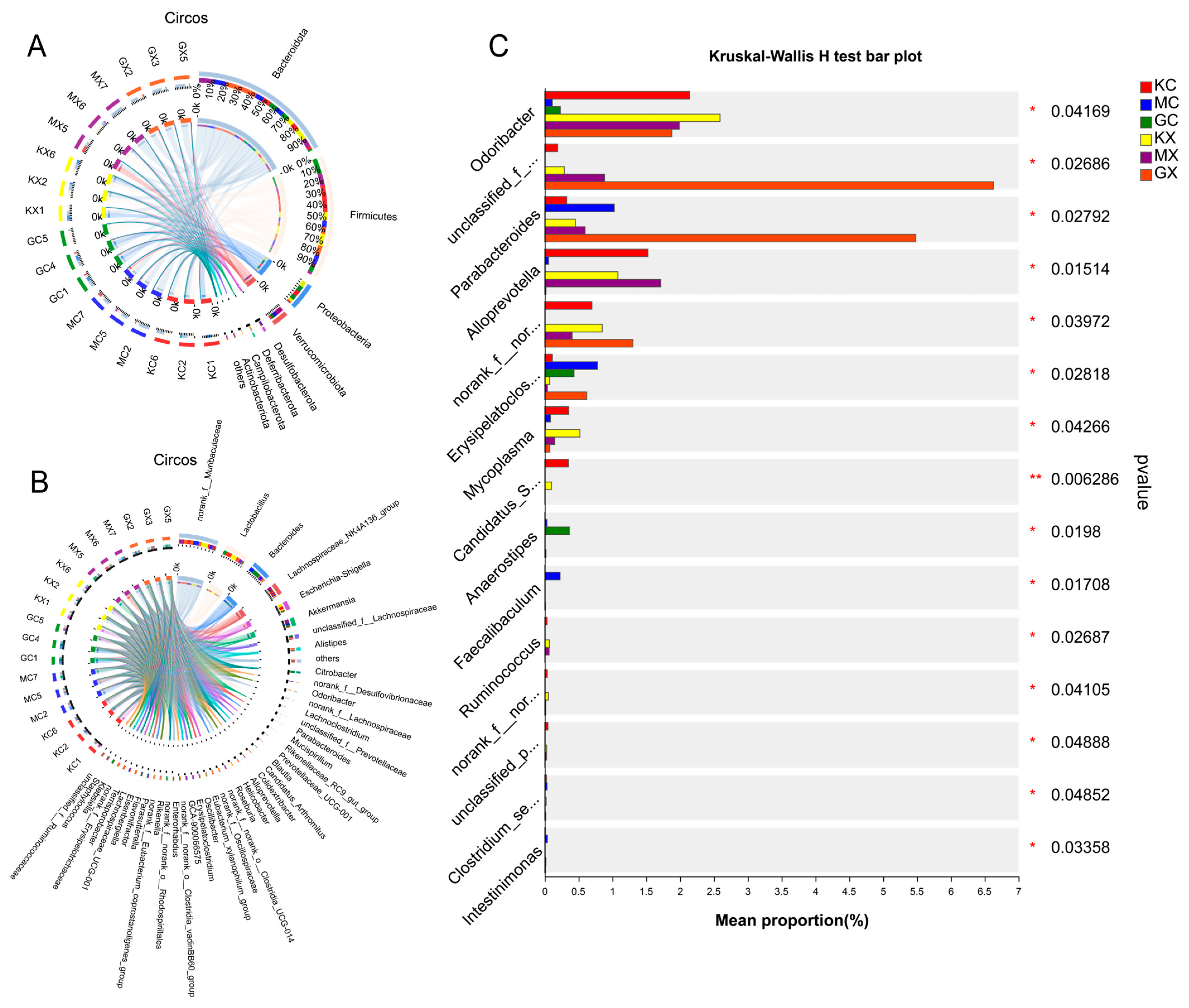
2.11. Analysis of the OTU Species Situation
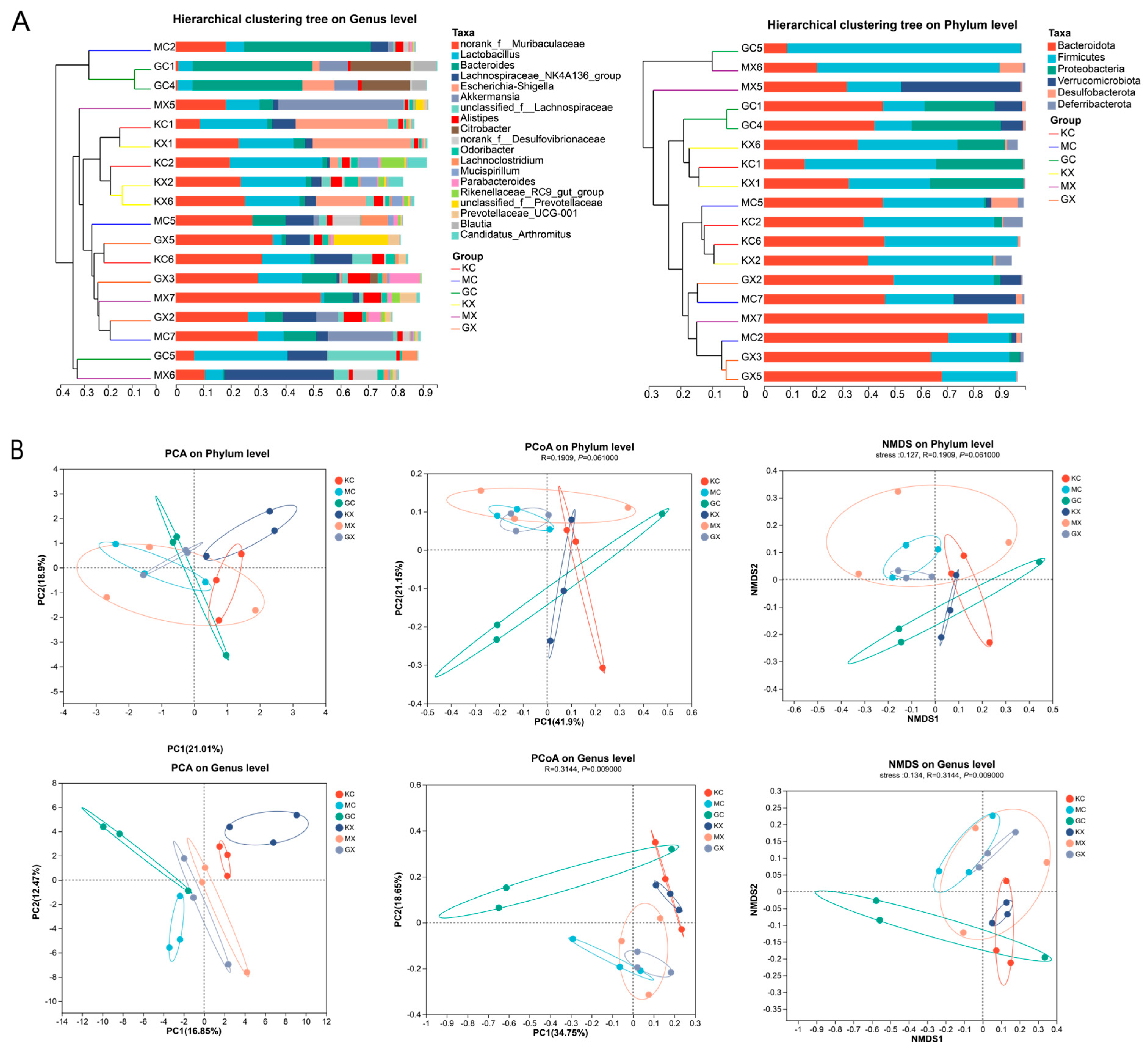
2.12. β Diversity Analysis
2.13. Association of Species Differences in the Mouse Intestinal Microbiota
3. Discussion
4. Materials and Methods
4.1. Strain, Animals, and Chemicals
4.2. GC-MS Analysis
4.3. Anti-C. albicans Assay
4.4. Conductivity Experiment
4.5. Protein Damage Test
4.6. Observation of C. albicans Biofilm Formation Stage by SEM
4.7. Construction of a Mouse Intestinal Model of C. albicans Infection
4.8. IL-6, IL-8, IL-17A Determination
4.9. HE and PAS Staining
4.10. Sequencing of Intestinal Flora of MICE Colonized by C. albicans
4.11. Data Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.H.; Kim, Y.G.; Lee, J. Antibiofilm activity of lawsone against polymicrobial enterohemorrhagic Escherichia coli O157:H7 and Candida albicans by suppression of curli production and hyphal growth. Phytomedicine 2024, 124, 155306. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2017, 16, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Antonia, L.; Michelle, M.; Alexander, S.M.; Mark, S.G.; Bernhard, H. In vitro infection models to study fungal–host interactions. FEMS Microbiol. Rev. 2021, 45, fuab005. [Google Scholar]
- Talapko, J.; Juzbai, M.; Matijevi, T.; Pustijanac, E.; Krlec, I. Candida albicans—The Virulence Factors and Clinical Manifestations of Infection. J. Fungi. 2021, 7, 79. [Google Scholar] [CrossRef]
- Daneshnia, F.; de Almeida Júnior, J.N.; Ilkit, M.; Lombardi, L.; Perry, A.M.; Gao, M.; Nobile, C.; Egger, M.; Perlin, D.; Zhai, B. Worldwide emergence of fluconazole-resistant Candida parapsilosis: Current framework and future research roadmap. Lancet Microbe 2023, 6, 470–480. [Google Scholar] [CrossRef]
- Lertsuphotvanit, N.; Tuntarawongsa, S.; Jitrangsri, K.P.T. Clotrimazole-Loaded Borneol-Based In Situ Forming Gel as Oral Sprays for Oropharyngeal Candidiasis Therapy. Gels 2023, 9, 412. [Google Scholar] [CrossRef]
- Chowdhary, A.; Jain, K.; Chauhan, N. Candida auris Genetics and Emergence. Annu. Rev. Microbiol. 2023, 77, 583602. [Google Scholar] [CrossRef]
- Bassi, R.C.; Boriollo, M.F.G. Amphotericin B, fluconazole, and nystatin as development inhibitors of Candida albicans biofilms on a dental prosthesis reline material: Analytical models invitro. J. Prosthet. Dent. 2022, 127, 320–330. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Carvalhaes, C.G.; Castanheira, M. Susceptibility patterns of amphotericin B, itraconazole, posaconazole, voriconazole and caspofungin for isolates causing invasive mould infections from the SENTRY Antifungal Surveillance Program (2018–2021) and application of single-site epidemiological cutoff values to evaluate amphotericin B activity. Mycoses 2023, 66, 854–868. [Google Scholar]
- Tragiannidis, A.; Gkampeta, A.; Vousvouki, M.; Vasileiou, E.; Groll, A.H. Antifungal agents and the kidney: Pharmacokinetics, clinical nephrotoxicity, and interactions. Expert Opin. Drug Saf. 2021, 20, 1061–1074. [Google Scholar] [CrossRef]
- Rakhshan, A.; Rahmati Kamel, B.; Saffaei, A.; Tavakoli-Ardakani, M. Hepatotoxicity Induced by Azole Antifungal Agents: A Review Study. Iran. J. Pharm. Res. 2023, 22, e130336. [Google Scholar] [CrossRef] [PubMed]
- Draskau, M.K.; Rosenmai, A.K.; Scholze, M.; Pedersen, M.; Boberg, J.; Christiansen, S.; Svingen, T. Human-relevant concentrations of the antifungal drug clotrimazole disrupts maternal and fetal steroid hormone profiles in rats. Toxicol. Appl. Pharm. 2021, 422, 115554. [Google Scholar] [CrossRef]
- Ben Hassine, D.; Kammoun El Euch, S.; Rahmani, R.; Ghazouani, N.; Kane, R.; Abderrabba, M.; Bouajila, J. Clove Buds Essential Oil: The Impact of Grinding on the Chemical Composition and Its Biological Activities Involved in Consumer’s Health Security. BioMed Res. Int. 2021, 2021, 9940591. [Google Scholar] [CrossRef] [PubMed]
- Noazira Wan Adnan, W.; Ulfah Karim, N.; Husna Yusoff, N.A.; Ihwan Zakariah, M.; Hassan, M. Effect of Cymbopogon citratus Essential Oil (EO) on Handling Stress in Giant Freshwater Prawn (Macrobrachium rosenbergii). Pak. J. Biol. Sci. 2021, 24, 13–18. [Google Scholar] [CrossRef]
- Pandey, V.K.; Srivastava, S.; Ashish; Dash, K.K.; Singh, R.; Dar, A.H.; Singh, T.; Farooqui, A.; Shaikh, A.M.; Kovacs, B. Bioactive properties of clove (Syzygium aromaticum) essential oil nanoemulsion: A comprehensive review. Heliyon 2024, 10, e22437. [Google Scholar] [CrossRef] [PubMed]
- Shahina, Z.; Molaeitabari, A.; Sultana, T.; Dahms, T.E.S. Cinnamon Leaf and Clove Essential Oils Are Potent Inhibitors of Candida albicans Virulence Traits. Microorganisms 2022, 10, 1989. [Google Scholar] [CrossRef]
- Kiki, M.J. In Vitro Antiviral Potential, Antioxidant, and Chemical Composition of Clove (Syzygium aromaticum) Essential Oi. Molecules 2023, 28, 2421. [Google Scholar] [CrossRef]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of clove (Syzygium aromaticum) oil and its major components to human skin cells. Cell Prolif. 2006, 39, 241–248. [Google Scholar] [CrossRef]
- Wijewantha, N.; Sane, S.; Eikanger, M.; Antony, R.M.; Potts, R.A.; Lang, L.; Rezvani, K.; Sereda, G. Enhancing anti-tumorigenic efficacy of eugenol in human colon cancer cells using enzyme-responsive nanoparticles. Cancers 2023, 15, 1145. [Google Scholar] [CrossRef]
- Shahina, Z.; Ndlovu, E.; Persaud, O.; Sultana, T.; Dahms, T.E.S. Candida albicans Reactive Oxygen Species (ROS)-Dependent Lethality and ROS-Independent Hyphal and Biofilm Inhibition by Eugenol and Citral. Microbiol. Spectr. 2022, 10, e3122–e3183. [Google Scholar] [CrossRef]
- El-Baz, A.M.; Mosbah, R.A.; Goda, R.M.; Mansour, B.; Sultana, T.; Dahms, T.E.S.; El-Ganiny, A.M. Back to Nature: Combating Candida albicans Biofilm, Phospholipase and Hemolysin Using Plant Essential Oils. Antibiotics 2021, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Farheen, M.; Kukreti, A.; Afzal, O.; Akhter, M.H.; Chitme, H.; Visht, S.; Altamimi, A.S.A.; Alossaimi, M.A.; Alsulami, E.R.; et al. Natural Oils Enhance the Topical Delivery of Ketoconazole by Nanoemulgel for Fungal Infections. ACS Omega 2023, 8, 28233–28248. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso-Ugarte, M.; Plaza-Díaz, J.; Gómez-Llorente, C.; Lucas Gómez, E.; Sabés-Alsina, M.; Gil, Á. In vitro examination of antibacterial and immunomodulatory activities of cinnamon, white thyme, and clove essential oils. J. Funct. Foods 2021, 81, 104436. [Google Scholar] [CrossRef]
- Luise, D.; Correa, F.; Negrini, C.; Virdis, S.; Mazzoni, M.; Dalcanale, S.; Trevisi, P. Blend of natural and natural identical essential oil compounds as a strategy to improve the gut health of weaning pigs. Anim. Int. J. Anim. Biosci. 2023, 17, 101031. [Google Scholar] [CrossRef]
- McGlynn, D.F.; Yee, L.D.; Garraffo, H.M.; Geer, L.Y.; Mak, T.D.; Mirokhin, Y.A.; Tchekhovskoi, D.V.; Jen, C.N.; Goldstein, A.H.; Kearsley, A.J.; et al. New Library-Based Methods for Nontargeted Compound Identification by GC-EI-MS. J. Am. Soc. Mass. Spectr. 2025, 36, 389–399. [Google Scholar] [CrossRef]
- Biernasiuk, A.; Baj, T.; Malm, A. Clove essential oil and its main constituent, eugenol, as potential natural antifungals against Candida spp. alone or in combination with other antimycotics due to synergistic interactions. Molecules 2022, 28, 215. [Google Scholar] [CrossRef]
- Viveiros, M.M.H.; Silva, M.G.; Da Costa, J.G.M.; de Oliveira, A.G.; Rubio, C.; Padovani, C.R.; Rainho, C.A.; Schellini, S.A. Anti-inflammatory effects of α-humulene and β-caryophyllene on pterygium fibroblasts. Int. J. Ophthalmol. 2022, 15, 1903. [Google Scholar] [CrossRef]
- Didehdar, M.; Chegini, Z.; Shariati, A. Eugenol: A novel therapeutic agent for the inhibition of Candida species infection. Front. Pharmacol. 2022, 13, 872127. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Yang, S.; Fu, B.; Chen, Y.; Zhou, M.; Qi, Y.; Xu, N.; Wu, Q.; Hua, Q.; Wu, Y. Antibacterial activity and mechanism of the sesquiterpene δ-cadinene against Listeria monocytogenes. LWT 2024, 203, 116388. [Google Scholar] [CrossRef]
- Li, M.; Zhao, Y.; Wang, Y.; Geng, R.; Fang, J.; Kang, S.G.; Huang, K.; Tong, T. Eugenol, a major component of clove oil, attenuates adiposity, and modulates gut microbiota in high-fat diet-fed mice. Mol. Nutr. Food Res. 2022, 66, 2200387. [Google Scholar] [CrossRef]
- Sangeeta, J.P.; Aishwarya, O.B.; Omkar, D.B.; Madhura, N.B. Anti-biofilm effect of clove oil against Candida albicans: A systematic review. J. Oral Maxillofac. Pathol. 2024, 28, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Chen, M.; Liu, J.; Ge, Y.; Wang, T.; Wu, D.; Yan, G.; Wang, C.; Shao, J. Sodium houttuyfonate attenuates dextran sulfate sodium associated colitis precolonized with Candida albicans through inducing β-glucan exposure. J. Leukoc. Biol. 2021, 110, 927–937. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, B.; Wang, F.; Tang, L.; Lei, Y.; Luo, Y.; Huang, S.; Yang, M.; Wu, L.; Wang, W. Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner. Theranostics 2020, 10, 5225. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Rosati, E.; Saggau, C.; Messner, B.; Chu, H.; Duan, Y.; Hartmann, P.; Wang, Y.; Ma, S.; Huang, W.J.M. Candida albicans-specific Th17 cell-mediated response contributes to alcohol-associated liver disease. Cell Host Microbe 2023, 31, 389–404. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, L.; Yang, L.; Chu, H. The critical role of gut microbiota in obesity. Front. Endocrinol. 2022, 13, 1025706. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Takezawa, K.; Fujita, K.; Matsushita, M.; Motooka, D.; Hatano, K.; Banno, E.; Shimizu, N.; Takao, T.; Takada, S.; Okada, K. The Firmicutes/Bacteroidetes ratio of the human gut microbiota is associated with prostate enlargement. Prostate 2021, 81, 1287–1293. [Google Scholar] [CrossRef]
- Rastogi, S.; Singh, A. Gut microbiome and human health: Exploring how the probiotic genus Lactobacillus modulate immune responses. Front. Pharmacol. 2022, 13, 1042189. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Ortiz-Alvarez De La Campa, M.; Curtis-Joseph, N.; Beekman, C.; Belenky, P. Gut biogeography accentuates sex-related differences in the murine microbiome. Microorganisms 2024, 12, 221. [Google Scholar] [CrossRef]
- NIST/EPA/NIH. NIST Standard Reference Database 1A: NIST/EPA/NIH Mass Spectral Library (NIST 23). 2023. Available online: https://www.nist.gov/srd/nist-standard-reference-database-1a (accessed on 2 April 2025).
- Ma, H.; Zhao, X.; Yang, L.; Su, P.; Fu, P.; Peng, J.; Yang, N.; Guo, G. Antimicrobial Peptide AMP-17 Affects Candida albicans by Disrupting Its Cell Wall and Cell Membrane Integrity. Infect. Drug Resist. 2020, 13, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Pavlin, M.; Kandušer, M.; Reberšek, M.; Pucihar, G.; Hart, F.X.; Magjarevićcacute, R.; Miklavčič, D. Effect of cell electroporation on the conductivity of a cell suspension. Biophys. J. 2005, 88, 4378–4390. [Google Scholar] [CrossRef]
- Li, N.; Gao, C.; Peng, X.; Wang, W.; Luo, M.; Fu, Y.; Zu, Y. Aspidin BB, a phloroglucinol derivative, exerts its antibacterial activity against Staphylococcus aureus by inducing the generation of reactive oxygen species. Res. Microbiol. 2014, 165, 263–272. [Google Scholar] [CrossRef]
- Farkash, Y.; Feldman, M.; Ginsburg, I.; Steinberg, D.; Shalish, M. Green Tea Polyphenols and Padma Hepaten Inhibit Candida albicans Biofilm Formation. Evid.-Based Complement. Altern. Med. 2018, 2018, 1690747. [Google Scholar] [CrossRef] [PubMed]
- Panariello, B.H.D.; Klein, M.I.; Mima, E.G.D.O.; Pavarina, A.C. Fluconazole impacts the extracellular matrix of fluconazole-susceptible and-resistant Candida albicans and Candida glabrata biofilms. J. Oral Microbiol. 2018, 10, 1476644. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.C.; Lupien, A.; Elhaji, Y.; Farag, M.; Marcus, V.; Behr, M.A. Establishment of persistent enteric mycobacterial infection following streptomycin pre-treatment. Gut Pathog. 2023, 15, 46. [Google Scholar] [CrossRef]
- Ying, M.; Yu, Q.; Zheng, B.; Wang, H.; Wang, J.; Chen, S.; Nie, S.; Xie, M. Cultured Cordyceps sinensis polysaccharides modulate intestinal mucosal immunity and gut microbiota in cyclophosphamide-treated mice. Carbohydr. Polym. 2020, 235, 115957. [Google Scholar] [CrossRef]
- Kim, H.; Hong, J.Y.; Lee, J.; Yeo, C.; Jeon, W.; Lee, Y.J.; Ha, I. Immune-boosting effect of Yookgong-dan against cyclophosphamide-induced immunosuppression in mice. Heliyon 2024, 10, e24033. [Google Scholar] [CrossRef]
- Koh, A.Y. Murine Models of Candida Gastrointestinal Colonization and Dissemination. Eukaryot. Cell 2013, 12, 1416–1422. [Google Scholar] [CrossRef]
- Petrocelli, G.; Farabegoli, F.; Valerii, M.C.; Giovannini, C.; Sardo, A.; Spisni, E. Molecules present in plant essential oils for prevention and treatment of colorectal cancer (CRC). Molecules 2021, 26, 885. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CAS | Retention Time/min | Calculated RI | NIST23 RI Value | Peak Area % |
|---|---|---|---|---|---|
| Eugenol | 97-53-0 | 20.812 | 1349 | 1357 ± 3 | 81.52 |
| Caryophyllene | 87-44-5 | 22.309 | 1422 | 1419 ± 3 | 14.45 |
| α-Humulene | 6753-98-6 | 23.181 | 1459 | 1454 ± 3 | 3.43 |
| δ-cadinene | 483-76-1 | 25.300 | 1519 | 1524 ± 2 | 0.60 |
| Time | Group | Concentration mg/(kg·d) | CFU/104 |
|---|---|---|---|
| 5 d | High-dose group | 300 | 1.53 ± 0.26 |
| Medium-dose group | 200 | 1.69 ± 0.37 | |
| Low-dose group | 100 | 1.83 ± 0.33 | |
| Clotrimazole group | 20 | 1.48 ± 0.21 | |
| Model group | — | 3.13 ± 0.19 | |
| Control group | — | 0 ± 0 |
| Index | Group | SE | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Blank Group | Model Group | L-CEO Group | M-CEO Group | H-CEO Group | CZT Group | |||
| Chao | 386.62 ± 9.99 Aa | 135.61 ± 22.31 Bb | 371.24 ± 16.80 Cc | 380.16 ± 20.41 Cc | 420.14 ± 0.72 Cd | 428.51 ± 0.72 Cd | 27.56 | <0.01 |
| ACE | 384.44 ± 8.18 Aa | 127.93 ± 11.87 Bb | 369.45 ± 7.45 Cc | 377.68 ± 22.90 Cc | 419.99 ± 12.20 Cd | 422.51 ± 12.20 Cd | 27.55 | <0.01 |
| Simpso | 0.16 ± 0.01 Aa | 0.44 ± 0.03 Bb | 0.06 ± 0.01 Cc | 0.13 ± 0.02 Cd | 0.09 ± 0.01 Ce | 0.08 ± 0.01 Ce | 0.44 | <0.01 |
| Shanno | 2.88 ± 0.55 Aa | 1.49 ± 0.37 Ab | 3.22 ± 0.56 Ac | 3.12 ± 0.74 Ac | 3.35 ± 0.52 Ac | 3.30 ± 0.52 Ac | 0.22 | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Guo, Y.; Chen, S.; Zhang, J.; Wang, J.; Wang, F.; Luo, J.; Yan, L. Impact of Clove Essential Oil on the Intestinal Flora in Mice Infected with Candida albicans. Molecules 2025, 30, 2372. https://doi.org/10.3390/molecules30112372
Gao Y, Guo Y, Chen S, Zhang J, Wang J, Wang F, Luo J, Yan L. Impact of Clove Essential Oil on the Intestinal Flora in Mice Infected with Candida albicans. Molecules. 2025; 30(11):2372. https://doi.org/10.3390/molecules30112372
Chicago/Turabian StyleGao, Yuan, Yuyang Guo, Shurong Chen, Jianmei Zhang, Jinhe Wang, Fuling Wang, Jianghan Luo, and Lijun Yan. 2025. "Impact of Clove Essential Oil on the Intestinal Flora in Mice Infected with Candida albicans" Molecules 30, no. 11: 2372. https://doi.org/10.3390/molecules30112372
APA StyleGao, Y., Guo, Y., Chen, S., Zhang, J., Wang, J., Wang, F., Luo, J., & Yan, L. (2025). Impact of Clove Essential Oil on the Intestinal Flora in Mice Infected with Candida albicans. Molecules, 30(11), 2372. https://doi.org/10.3390/molecules30112372