
Matrine Restores Colistin Efficacy Against mcr-1-Carrying Escherichia coli

Abstract
1. Introduction
2. Results and Discussion
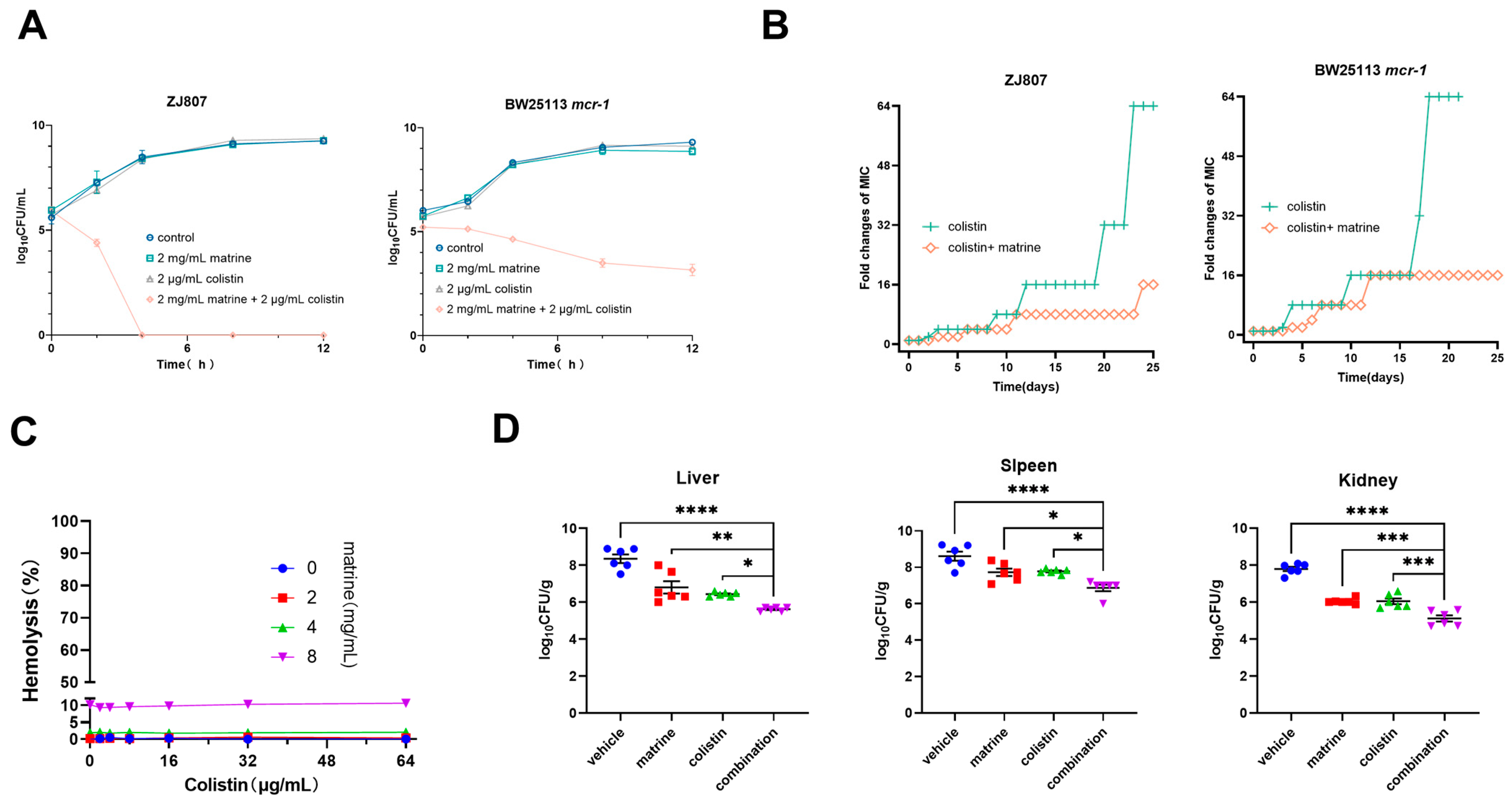
2.1. Synergistic Activity of Matrine with Colistin
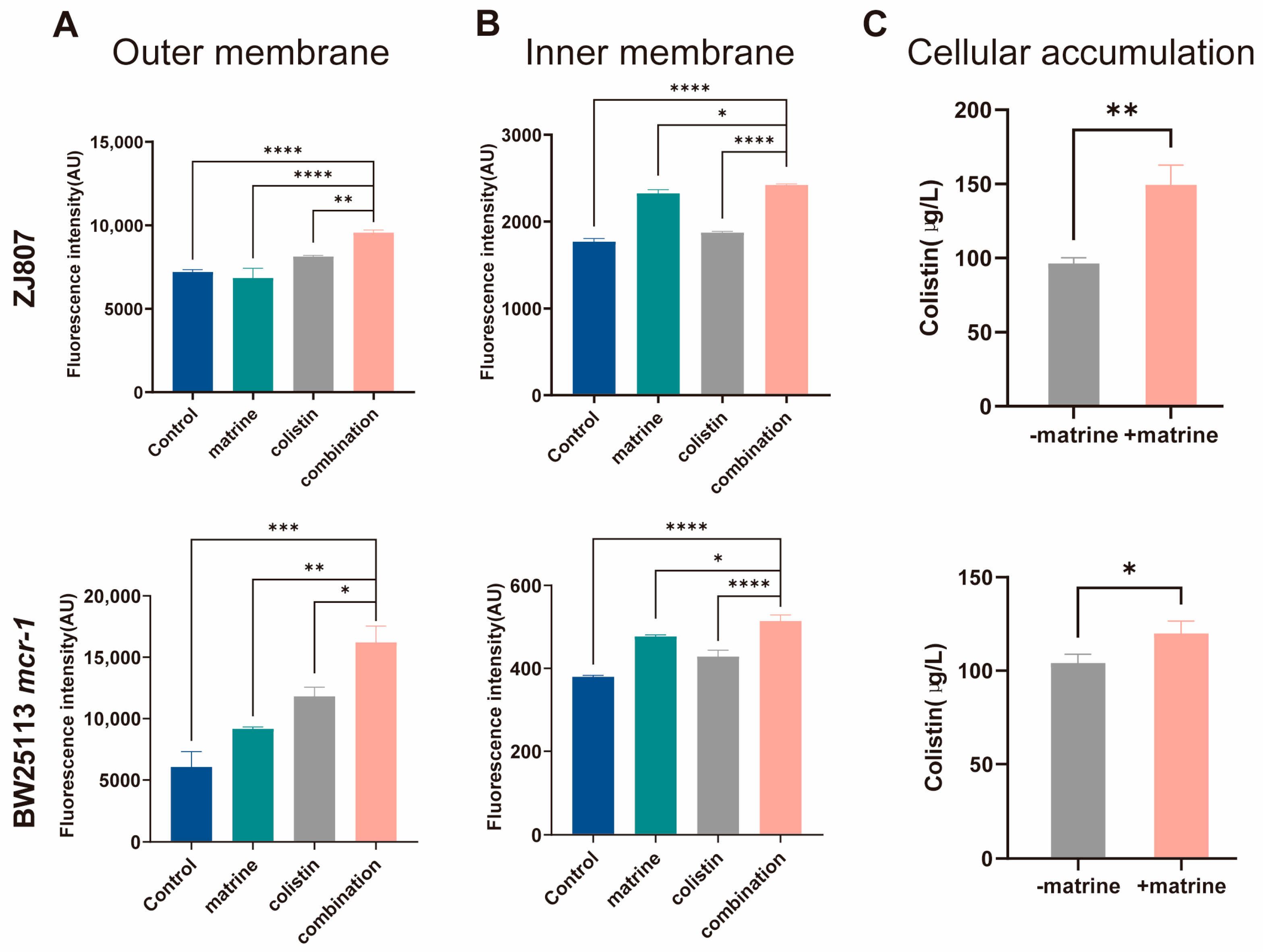
2.2. Matrine Enhances Colistin-Mediated Membrane Damage and Intracellular Drug Accumulation
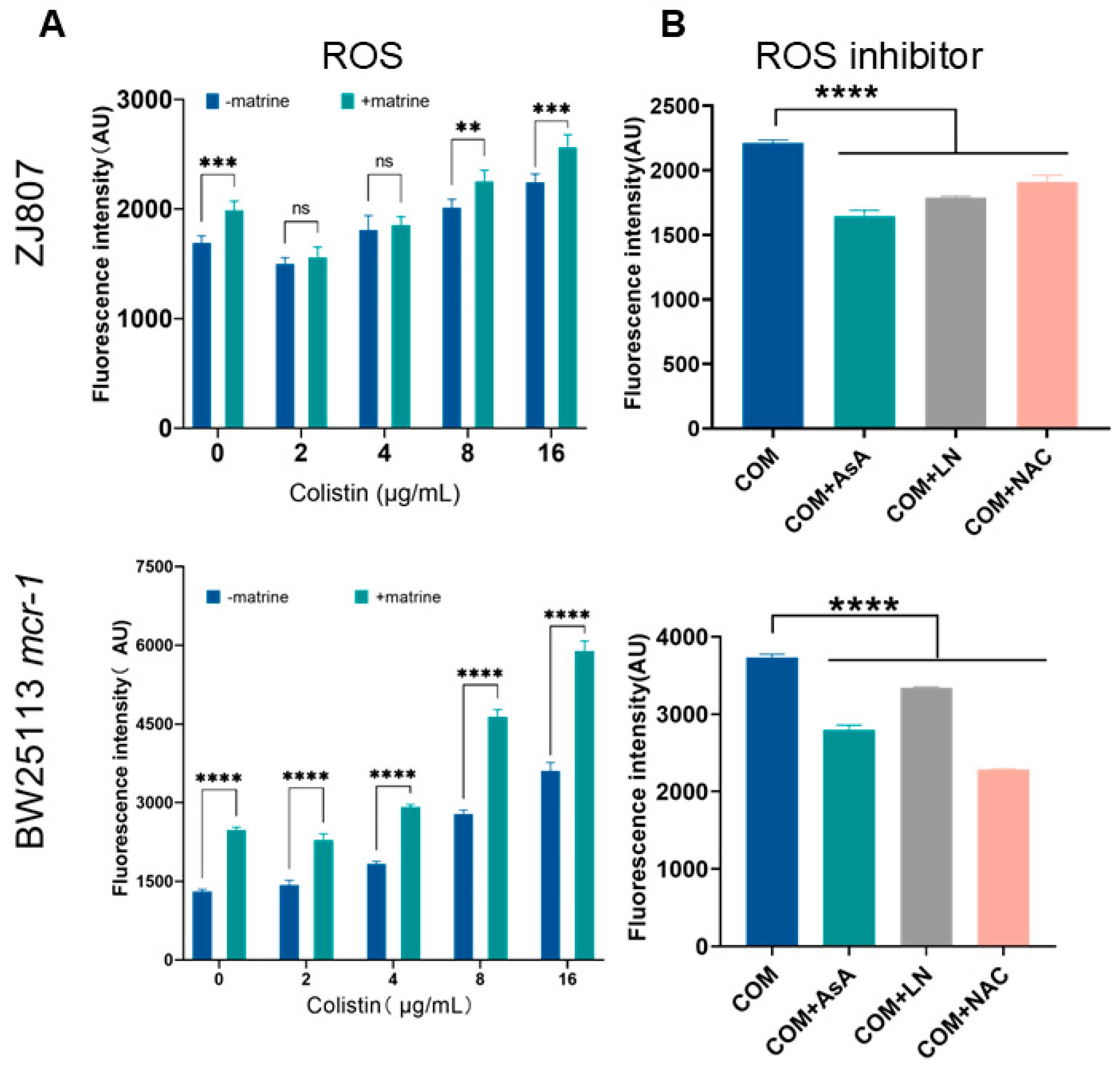
2.3. Colistin-Matrine Combination Promotes Oxidative Damage
3. Materials and Methods
3.1. Bacteria and Reagents
3.2. MIC Assay and Antimicrobial Combination Test
3.3. Time-Kill Curves
3.4. Resistance Development Assay
3.5. Assessment of Matrine on Colistin-Induced Hemolysis
3.6. Mouse Infection Assays
3.7. Outer Membrane Integrity
3.8. Inner Membrane Integrity
3.9. Intracellular Colistin Accumulation
3.10. Total ROS
3.11. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Doi, Y.; Murray, G.L.; Peleg, A.Y. Acinetobacter baumannii: Evolution of antimicrobial resistance-treatment options. Semin. Respir. Crit. Care. Med. 2015, 36, 85–98. [Google Scholar] [PubMed]
- Liu, Y.; Wang, Y.; Walsh, T.R.; Yi, L.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet. Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes. Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Liu, Y.-Y.; Shen, Y.-B.; Yang, J.; Walsh, T.R.; Wang, Y.; Shen, J. Plasmid-mediated colistin-resistance genes: mcr. Trends. Microbiol. 2023, 32, 365–378. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.-G.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and its role in the era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes. Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Li, B.; Ke, B.; Zhao, X.; Guo, Y.; Wang, W.; Wang, X.; Zhu, H. Antimicrobial resistance profile of mcr-1 positive clinical isolates of Escherichia coli in China from 2013 to 2016. Front. Microbiol. 2018, 9, 2514. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, Y.; Feng, Y. Towards understanding MCR-like colistin resistance. Trends. Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef]
- Dhanda, G.; Acharya, Y.; Haldar, J. Antibiotic adjuvants: A versatile approach to combat antibiotic resistance. ACS. Omega. 2023, 8, 10757–10783. [Google Scholar] [CrossRef]
- Zhou, J.; Qian, Y.; Lang, Y.; Zhang, Y.; Tao, X.; Moya, B.; Sayed, A.R.M.; Landersdorfer, C.B.; Shin, E.; Werkman, C.; et al. Comprehensive stability analysis of 13 β-lactams and β-lactamase inhibitors in in vitro media, and novel supplement dosing strategy to mitigate thermal drug degradation. Antimicrob. Agents. Chemother. 2024, 68, e0139923. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhou, S.; Liang, Y.; Wei, Y.; Li, Y.; Long, T.; He, Q.; Li, M.; Zhou, Y.; Yu, Y.; et al. Natural flavonoids disrupt bacterial iron homeostasis to potentiate colistin efficacy. Sci. Adv. 2023, 9, eadg4205. [Google Scholar] [CrossRef]
- Gadar, K.; de Dios, R.; Kadeřábková, N.; Prescott, T.A.K.; Mavridou, D.A.I.; McCarthy, R.R. Disrupting iron homeostasis can potentiate colistin activity and overcome colistin resistance mechanisms in Gram-Negative Bacteria. Commun. Biol. 2023, 6, 937. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules. 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, X.; Zhang, M.; Zou, Z.; Yang, S.; Yi, T.; Wang, J.; Liu, D.; Shen, Y.; Dai, C.; et al. Dual effects of feed-additive-derived chelerythrine in combating mobile colistin resistance. Engineering. 2023, 32, 163–173. [Google Scholar] [CrossRef]
- Li, N.; Tan, S.; Cui, J.; Guo, N.; Wang, W.; Zu, Y.; Jin, S.; Xu, X.; Liu, Q.; Fu, Y. PA-1, a novel synthesized pyrrolizidine alkaloid, inhibits the growth of Escherichia coli and Staphylococcus aureus by damaging the cell membrane. J. Antibiot. 2014, 67, 689–696. [Google Scholar] [CrossRef]
- Jin, J.; Hua, G.; Meng, Z.; Gao, P. Antibacterial mechanisms of berberine and reasons for little resistance of bacteria. Chin. Herb. Med. 2011, 3, 27–35. [Google Scholar]
- Li, X.; Tang, Z.; Wen, L.; Jiang, C.; Feng, Q. Matrine: A review of its pharmacology, pharmacokinetics, toxicity, clinical application and preparation researches. J. Ethnopharmacol. 2021, 269, 113682. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, L.; Cai, Q.; Wang, M.; Wang, X.; Wang, S.; Ni, Z. Research progress on the pharmacological effects of matrine. Front. Neurosci. 2022, 16, 977374. [Google Scholar] [CrossRef]
- Pourahmad Jaktaji, R.; Koochaki, S. In vitro activity of honey, total alkaloids of Sophora alopecuroides and matrine alone and in combination with antibiotics against multidrug-resistant Pseudomonas aeruginosa isolates. Lett. Appl. Microbiol. 2022, 75, 70–80. [Google Scholar] [CrossRef]
- Wei, S.; Xiao, J.; Ju, F.; Liu, J.; Hu, Z. A review on the pharmacology, pharmacokinetics and toxicity of Sophocarpine. Front. Pharmacol. 2024, 15, 1353234. [Google Scholar] [CrossRef]
- Wang, R.; Deng, X.; Gao, Q.; Wu, X.; Han, L.; Gao, X.; Zhao, S.; Chen, W.; Zhou, R.; Li, Z.; et al. Sophora alopecuroides L.: An ethnopharmacological, phytochemical, and pharmacological review. J. Ethnopharmacol. 2020, 248, 112172. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.; Andersson, D.I. Evolutionary consequences of drug resistance: Shared principles across diverse targets and organisms. Nat. Rev. Genet. 2015, 16, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Dai, C.; Ciccotosto, G.D.; Cappai, R.; Hoyer, D.; Li, J. Polymyxins for CNS infections: Pharmacology and neurotoxicity. Pharmacol. Ther. 2018, 181, 85–90. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Fang, J.; Huang, L.; Wang, J.; Huang, X. Sophora flavescens Ait.: Traditional usage, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2015, 172, 10–29. [Google Scholar] [CrossRef]
- Guo, B.; Zhu, S.; Zhang, C.; Wang, H.; Bao, L. Study on combined antibacterial activities of matrine, oxymatrine and Cnidium Monnieri (L.) Cusson. Chin. Arch. Tradit. Chin. Med. 2011, 6, 1398–1400. [Google Scholar]
- Jia, F.; Zhou, Q.; Li, X.; Zhou, X. Total alkaloids of Sophora alopecuroides and matrine inhibit auto-inducer 2 in the biofilms of Staphylococcus epidermidis. Microb. Pathog. 2019, 136, 103698. [Google Scholar] [CrossRef]
- Cely-Veloza, W.; Kato, M.J.; Coy-Barrera, E. Quinolizidine-type alkaloids: Chemodiversity, occurrence, and bioactivity. ACS Omega. 2023, 8, 27862–27893. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Liu, T.; He, W.; Chen, Y.; Chen, X.-H.; Li, X.; Zhou, W.; Yi, J.; Ren, Z. Antitumor effect of matrine in human hepatoma G2 cells by inducing apoptosis and autophagy. World J. Gastroenterol. 2010, 16, 4281–4290. [Google Scholar] [CrossRef]
- Wang, X.; Wu, F.; Huang, Y.; Li, H.; Cao, X.-Y.; You, Y.; Meng, Z.; Sun, K.-Y.; Shen, X. Matrine suppresses NLRP3 inflammasome activation via regulating PTPN2/JNK/SREBP2 pathway in sepsis. Phytomedicine. 2023, 109, 154574. [Google Scholar] [CrossRef]
- Meng, J.; Wang, W.; Ding, J.; Gu, B.; Zhou, F.; Wu, D.; Fu, X.; Qiao, M.; Liu, J. The synergy effect of matrine and berberine hydrochloride on treating colibacillosis caused by an avian highly pathogenic multidrug-resistant Escherichia coli. Poult. Sci. 2024, 103, 104151. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, H.; Sun, P.; Khan, A.; Guo, J.; Zheng, X.; Sun, Y.; Fan, K.; Yin, W.; Li, H. Matrine exhibits antiviral activity in a PRRSV/PCV2 co-infected mouse model. Phytomedicine. 2020, 77, 153289. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhu, X.; Wang, P.; Zhu, K.; Liu, X.; Ma, D.; Zhao, Q.; Hao, Z. Combining with matrine restores ciprofloxacin efficacy against qnrS producing E. coli in vitro and in vivo. Microb. Pathog. 2025, 198, 107132. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, Z.; Yang, Y.; Wang, F.; Deng, L. Antimicrobial activity of Sophora alopecuroides alkaloids. J. Beijing Univ. Chem. Technol. 2011, 38, 84–88. [Google Scholar]
- Cai, W.; Fu, Y.; Zhang, W.; Chen, X.; Zhao, J.; Song, W.; Li, Y.; Huang, Y.; Wu, Z.; Sun, R.; et al. Synergistic effects of baicalein with cefotaxime against Klebsiella pneumoniae through inhibiting CTX-M-1 gene expression. BMC Microbiol. 2016, 16, 181. [Google Scholar] [CrossRef]
- Sabnis, A.; Hagart, K.L.; Klöckner, A.; Becce, M.; Evans, L.E.; Furniss, R.C.D.; Mavridou, D.A.; Murphy, R.; Stevens, M.M.; Davies, J.C.; et al. Colistin kills bacteria by targeting lipopolysaccharide in the cytoplasmic membrane. Elife. 2021, 10, e65836. [Google Scholar] [CrossRef]
- Smith, W.P.J.; Wucher, B.R.; Nadell, C.D.; Foster, K.R. Bacterial defences: Mechanisms, evolution and antimicrobial resistance. Nat. Rev. Microbiol. 2023, 21, 519–534. [Google Scholar] [CrossRef]
- Cai, J.; Shi, J.; Chen, C.; He, M.; Wang, Z.; Liu, Y. Structural-activity relationship-inspired the discovery of saturated fatty acids as novel colistin enhancers. Adv. Sci. 2023, 10, 2302182. [Google Scholar] [CrossRef]
- Rosas, N.C.; Lithgow, T. Targeting bacterial outer-membrane remodelling to impact antimicrobial drug resistance. Trends Microbiol. 2022, 30, 544–552. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, Y.; Yang, K.; Tong, Z.; Shi, J.; Li, R.; Xiao, X.; Ren, W.; Hardeland, R.; Reiter, R.J.; et al. Melatonin overcomes MCR-mediated colistin resistance in Gram-negative pathogens. Theranostics. 2020, 10, 10697–10711. [Google Scholar] [CrossRef]
- Stokes, J.M.; MacNair, C.R.; Ilyas, B.; French, S.; Côté, J.-P.; Bouwman, C.; Farha, M.A.; Sieron, A.O.; Whitfield, C.; Coombes, B.K.; et al. Pentamidine sensitizes Gram-negative pathogens to antibiotics and overcomes acquired colistin resistance. Nat. Microbiol. 2017, 2, 17028. [Google Scholar] [CrossRef]
- Xu, T.; Fang, D.; Li, F.; Wang, Z.; Liu, Y. Vitamin B6 resensitizes mcr-carrying Gram-negative bacteria to colistin. Commun. Biol. 2025, 8, 459. [Google Scholar] [CrossRef] [PubMed]
- Sundaramoorthy, N.S.; Suresh, P.; Selva Ganesan, S.; GaneshPrasad, A.; Nagarajan, S. Restoring colistin sensitivity in colistin-resistant E. coli: Combinatorial use of MarR inhibitor with efflux pump inhibitor. Sci. Rep. 2019, 9, 19845. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Tian, P.; Li, Q.; Meng, B.; Ding, Y.; Liu, Y.; Li, Y.; Yu, L.; Li, J. Gallium nitrate enhances antimicrobial activity of colistin against Klebsiella pneumoniae by inducing reactive oxygen species accumulation. Microbiol. Spectr. 2023, 11, e0033423. [Google Scholar] [CrossRef]
- Chen, X.; Song, M.; Tian, L.; Shan, X.; Mao, C.; Chen, M.; Zhao, J.; Sami, A.; Yin, H.; Ali, U.; et al. A plant peptide with dual activity against multidrug-resistant bacterial and fungal pathogens. Sci. Adv. 2025, 11, eadt8239. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, G.; Zhang, R.; Shen, Y.; Tyrrell, J.M.; Huang, X.; Zhou, H.; Lei, L.; Li, H.; Doi, Y.; et al. Prevalence, risk factors, outcomes, and molecular epidemiology of mcr-1-positive Enterobacteriaceae in patients and healthy adults from China: An epidemiological and clinical study. Lancet Infect. Dis. 2017, 17, 390–399. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, J.; Liu, S.; Du, J.; Jia, X.; Wang, Z.; Ge, L.; Cui, K.; Ga, Y.; Li, X.; et al. Inosine monophosphate overcomes the coexisting resistance of mcr-1 and blaNDM-1 in Escherichia coli. J. Adv. Res. 2025, S2090-1232(25)00203-6. [Google Scholar] [CrossRef]
- Ma, T.; Fu, J.; Xie, N.; Ma, S.; Lei, L.; Zhai, W.; Shen, Y.; Sun, C.; Wang, S.; Shen, Z.; et al. Fitness cost of blaNDM-5-carrying p3R-IncX3 plasmids in wild-type NDM-free Enterobacteriaceae. Microorganisms. 2020, 8, 377. [Google Scholar] [CrossRef]
- Wang, T.; Yi, X.; Le, T.H.; Sivachidambaram, V.; Zhou, Z. Selective pressure of various levels of erythromycin on the development of antibiotic resistance. Environ. Pollut. 2025, 368, 125757. [Google Scholar] [CrossRef]
- Emara, Y.; Jolliet, O.; Finkbeiner, M.; Heß, S.; Kosnik, M.; Siegert, M.-W.; Fantke, P. Comparative selective pressure potential of antibiotics in the environment. Environ. Pollut. 2023, 318, 120873. [Google Scholar] [CrossRef]
- Tantisuwanno, C.; Dang, F.; Bender, K.; Spencer, J.D.; Jennings, M.E.; Barton, H.A.; Joy, A. Synergism between rifampicin and cationic polyurethanes overcomes intrinsic resistance of Escherichia coli. Biomacromolecules 2021, 22, 2910–2920. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Origin (Isolation Location) | Resistance Profile | MIC (μg/mL) | Reference |
|---|---|---|---|---|
| ZJ807 | Zhejiang, clinical | mcr-1 | Colistin: 4 | [46] |
| BW25113 | Lab-constructed | mcr-1 | Colistin: 4 | [14] |
| 4F-2 | Unknown | mcr-1 | Colistin: 4 | this study |
| 2F-2 | Unknown | mcr-1 | Colistin: 4 | [47] |
| BW25113 | Lab-constructed | mcr-5 | Colistin: 4 | this study |
| 3R | Shandong, environmental | blaNDM-5 | Meropenem: 128 | [48] |
| Z107 | Zhejiang, clinical | blaOXA-232 | Meropenem: 32 | this study |
| Z18 | Zhejiang, clinical | blaNDM-5 | Meropenem: 32 | this study |
| Z456 | Zhejiang, clinical | blaKPC-2 | Meropenem: 32 | this study |
| Z270 | Zhejiang, clinical | blaNDM-4 | Meropenem: 32 | this study |
| Z690 | Zhejiang, clinical | blaNDM-5, blaKPC-2 | Meropenem: 64 | this study |
| E305-2 | Zhejiang, clinical | blaIMP-4 | Meropenem: 8 | this study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Li, X.; Zhao, L.; Liu, S.; Du, J.; Jia, X.; Ge, L.; Xu, J.; Cui, K.; Ga, Y.; et al. Matrine Restores Colistin Efficacy Against mcr-1-Carrying Escherichia coli. Molecules 2025, 30, 2122. https://doi.org/10.3390/molecules30102122
Wang Z, Li X, Zhao L, Liu S, Du J, Jia X, Ge L, Xu J, Cui K, Ga Y, et al. Matrine Restores Colistin Efficacy Against mcr-1-Carrying Escherichia coli. Molecules. 2025; 30(10):2122. https://doi.org/10.3390/molecules30102122
Chicago/Turabian StyleWang, Zhinan, Xiaowei Li, Liang Zhao, Saiwa Liu, Jingjing Du, Xi Jia, Lirui Ge, Jian Xu, Kexin Cui, Yu Ga, and et al. 2025. "Matrine Restores Colistin Efficacy Against mcr-1-Carrying Escherichia coli" Molecules 30, no. 10: 2122. https://doi.org/10.3390/molecules30102122
APA StyleWang, Z., Li, X., Zhao, L., Liu, S., Du, J., Jia, X., Ge, L., Xu, J., Cui, K., Ga, Y., Wang, J., & Xia, X. (2025). Matrine Restores Colistin Efficacy Against mcr-1-Carrying Escherichia coli. Molecules, 30(10), 2122. https://doi.org/10.3390/molecules30102122