
LC-MS-Based Metabolomics Reveals the Mechanism of Protection of Berberine against Indomethacin-Induced Gastric Injury in Rats
Abstract
1. Introduction
2. Results
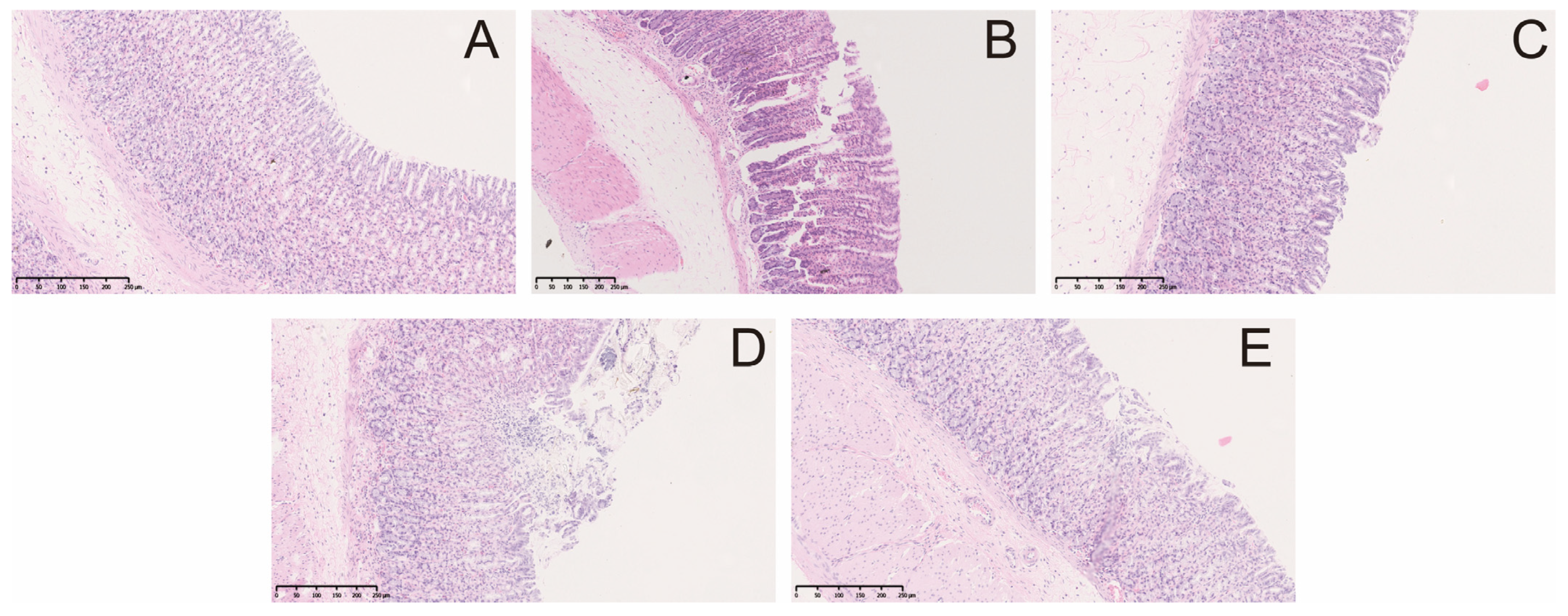
2.1. Effect of Berberine on Gastric Lesions Induced by Indomethacin
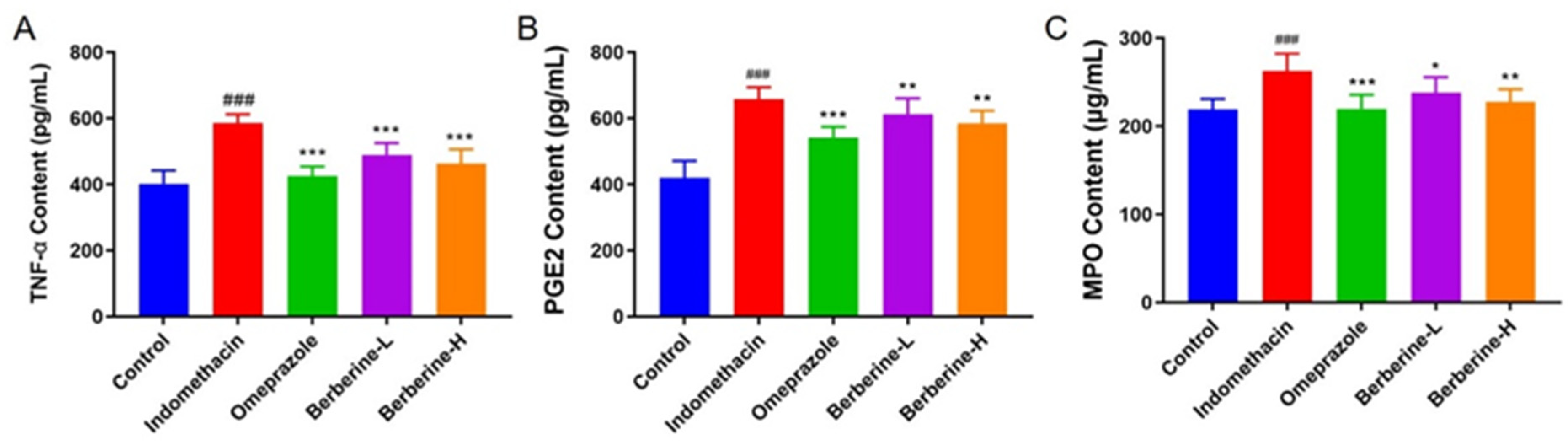
2.2. Effect of Berberine on Levels of Inflammatory Cytokine Markers in Serum
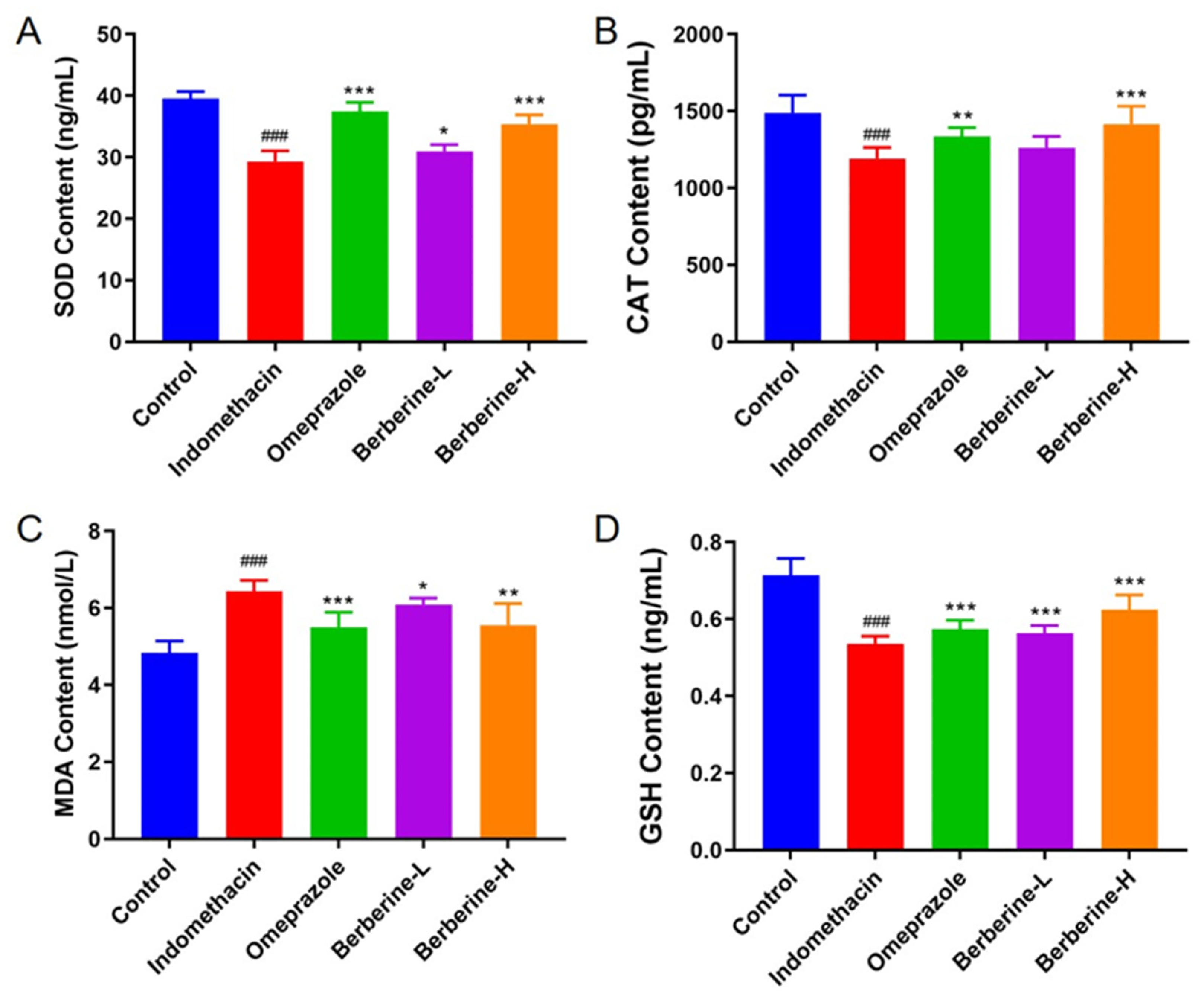
2.3. Effect of Berberine on Antioxidant Biomarkers in Serum
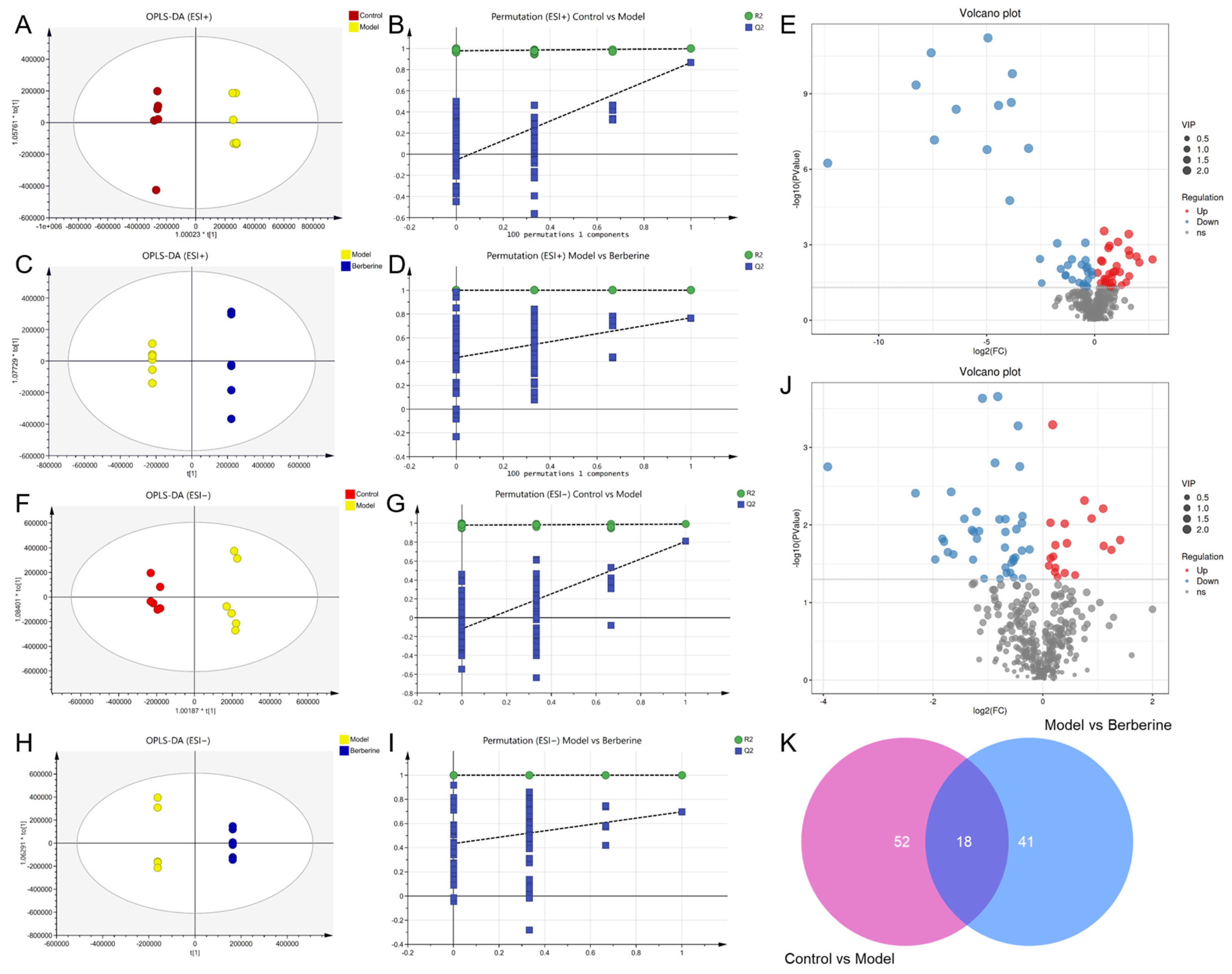
2.4. Multivariate Statistical Analysis and Potential Biomarkers Exploring
2.5. Identification and Analysis of Metabolic Biomarkers
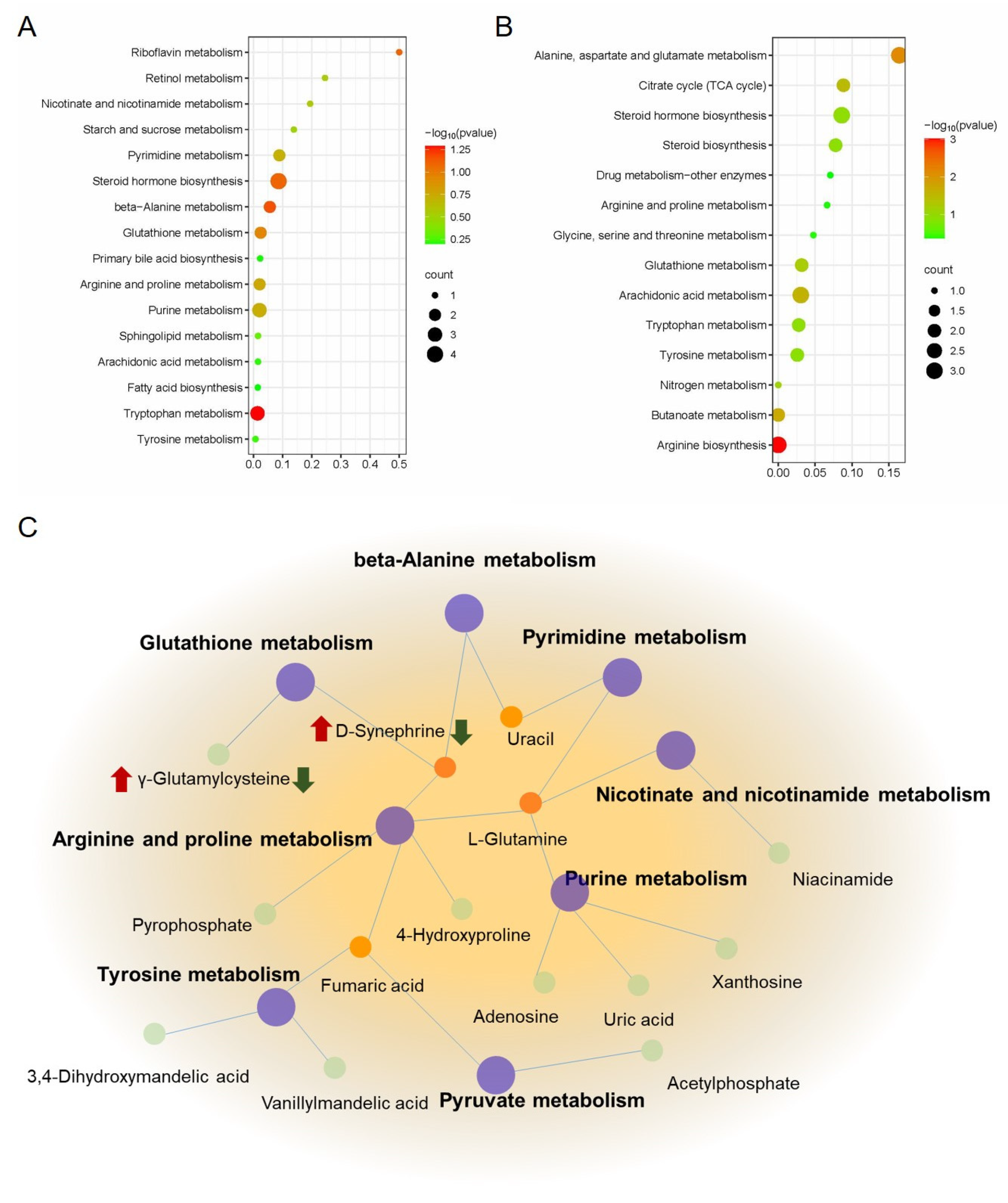
2.6. Metabolic Pathway Enrichment Analysis
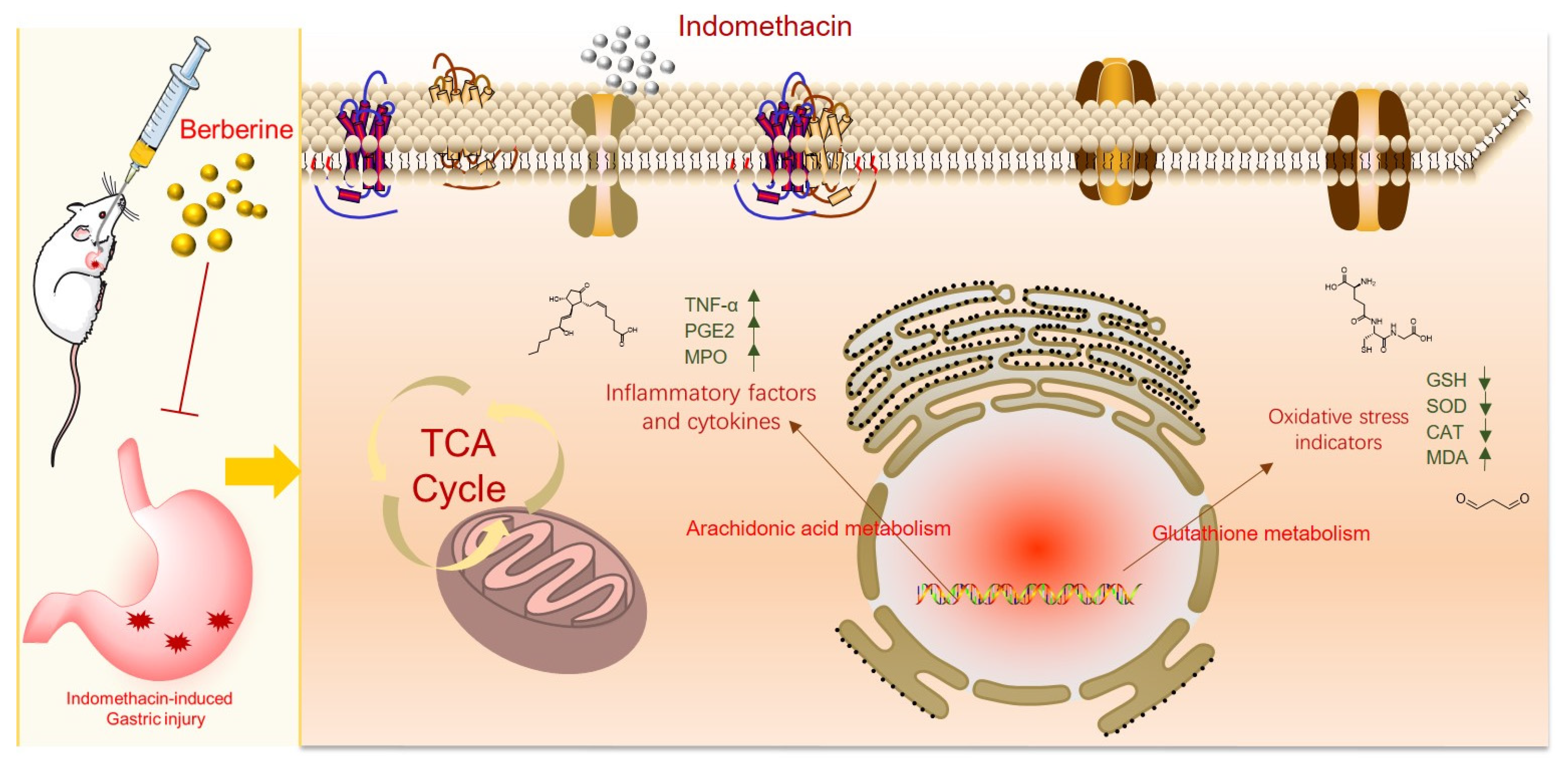
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Animals and Indomethacin-Induced GI Model Establishment
4.3. Histopathological Examination
4.4. Measurement of Specific Serum Biomarkers
4.5. Metabolomics Analysis
4.5.1. Serum Sample Handling
4.5.2. QC Samples
4.5.3. Chromatography and Mass Spectrometry
4.5.4. Data Processing and Multivariate Analysis
4.5.5. Potential Biomarker Identification and Pathway Enrichment Analysis
4.5.6. Pathway Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, Y.; Ren, S.; Wang, J.; Wang, Y.; Cui, Y.; Tian, M.; Wang, R.; Liu, H.; Zhao, Y. Dehydroevodiamine ameliorates indomethacin-induced gastric injury via inhibition of ERK and p38 signaling pathway. Phytomedicine 2021, 93, 153764. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wang, R.; Ren, S.; Liu, X.; Jing, M.; Li, R.; Tong, Y.; Wen, J.; Yang, T.; Wang, J.; et al. Zuojin Pill ameliorates inflammation in indomethacin-induced gastric injury via inhibition of MAPK pathway. J. Ethnopharmacol. 2021, 275, 114103. [Google Scholar] [CrossRef] [PubMed]
- Um, S.Y.; Park, J.H.; Chung, M.W.; Kim, K.B.; Kim, S.H.; Choi, K.H.; Lee, H.J. Nuclear magnetic resonance-based metabolomics for prediction of gastric damage induced by indomethacin in rats. Anal. Chim. Acta 2012, 722, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Allam, M.M.; El-Gohary, O.A. Gastroprotective effect of ghrelin against indomethacin-induced gastric injury in rats: Possible role of heme oxygenase-1 pathway. Gen. Physiol. Biophys. 2017, 36, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Alkushi, A.G.R.; Elsawy, N.A.M. Quercetin attenuates, indomethacin-induced acute gastric ulcer in rats. Folia Morphol. 2017, 76, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Nabil, M.; El Raey, M.A.; Abdo, W.; Abdelfattah, M.A.O.; El-Shazly, A.M.; Sobeh, M.; Mahmoud, M.F. Gastro-protective effects of Albizia anthelmintica leaf extract on indomethacin-induced gastric ulcer in Wistar rats: In silico and in vivo studies. Antioxidants 2021, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L. Prostaglandins, NSAIDs, and gastric mucosal protection: Why doesn’t the stomach digest itself? Physiol. Rev. 2008, 88, 1547–1565. [Google Scholar] [CrossRef] [PubMed]
- Mard, S.A.; Pipelzadeh, M.H.; Teimoori, A.; Neisi, N.; Mojahedin, S.; Khani, M.Z.; Ahmadi, I. Protective activity of crocin against indomethacin-induced gastric lesions in rats. J. Nat. Med. 2016, 70, 62–74. [Google Scholar] [CrossRef]
- Luo, C.; Chen, H.; Wang, Y.; Lin, G.; Li, C.; Tan, L.; Su, Z.; Lai, X.; Xie, J.; Zeng, H. Protective effect of coptisine free base on indomethacin-induced gastric ulcers in rats: Characterization of potential molecular mechanisms. Life Sci. 2018, 193, 47–56. [Google Scholar] [CrossRef]
- Brusselaers, N.; Wahlin, K.; Engstrand, L.; Lagergren, J. Maintenance therapy with proton pump inhibitors and risk of gastric cancer: A nationwide population-based cohort study in Sweden. BMJ Open 2017, 7, e017739. [Google Scholar] [CrossRef]
- Nadatani, Y.; Watanabe, T.; Suda, W.; Nakata, A.; Matsumoto, Y.; Kosaka, S.; Higashimori, A.; Otani, K.; Hosomi, S.; Tanaka, F.; et al. Gastric acid inhibitor aggravates indomethacin-induced small intestinal injury via reducing Lactobacillus johnsonii. Sci. Rep. 2019, 9, 17490. [Google Scholar] [CrossRef] [PubMed]
- Antonisamy, P.; Arasu, M.V.; Dhanasekaran, M.; Choi, K.C.; Aravinthan, A.; Kim, N.S.; Kang, C.W.; Kim, J. Protective effects of trigonelline against indomethacin-induced gastric ulcer in rats and potential underlying mechanisms. Food Funct. 2016, 7, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Warowicka, A.; Nawrot, R.; Gozdzicka-Jozefiak, A. Antiviral activity of berberine. Arch. Virol. 2020, 165, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tang, J.; Chen, S.; Hu, S.; Shen, C.; Xiang, J.; Chen, N.; Wang, J.; Ma, X.; Zhang, Y.; et al. Berberine for gastric cancer prevention and treatment: Multi-step actions on the Correa’s cascade underlie its therapeutic effects. Pharmacol. Res. 2022, 184, 106440. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Fukuda, Y.; Yamamoto, M.; Miura, N.; Fukutake, M.; Ishige, A.; Yamaguchi, R.; Nagasaki, M.; Saito, A.; Imoto, S.; Miyano, S.; et al. Orengedokuto and berberine improve indomethacin-induced small intestinal injury via adenosine. J. Gastroenterol. 2009, 44, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; Van Ommen, B.; Smilde, A.K. Large-scale human metabolomics studies: A strategy for data (pre-) processing and validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Zhang, Y.B.; Yang, X.W.; Xu, J.; Wang, Z.J.; Sun, Y.Z.; Xu, W.; Wang, Y.P. Application of UPLC-Triple TOF-MS/MS metabolomics strategy to reveal the dynamic changes of triterpenoid saponins during the decocting process of Asian ginseng and American ginseng. Food Chem. 2023, 424, 136425. [Google Scholar] [CrossRef]
- Corona, G.; Cannizzaro, R.; Miolo, G.; Caggiari, L.; De Zorzi, M.; Repetto, O.; Steffan, A.; De Re, V. Use of metabolomics as a complementary omic approach to implement risk criteria for first-degree relatives of gastric cancer patients. Int. J. Mol. Sci. 2018, 19, 750. [Google Scholar] [CrossRef]
- Lang, J.; Li, L.; Quan, Y.; Tan, R.; Zhao, J.; Li, M.; Zeng, J.; Chen, S.; Wang, T.; Li, Y.; et al. LC-MS-based metabolomics reveals the mechanism of anti-gouty arthritis effect of Wuwei Shexiang pill. Front. Pharmacol. 2023, 14, 1213602. [Google Scholar] [CrossRef]
- Du, Z.; Lu, Y.; Sun, J.; Chang, K.; Lu, M.; Fang, M.; Zeng, X.; Zhang, W.; Song, J.; Guo, X.; et al. Pharmacokinetics/pharmacometabolomics-pharmacodynamics reveals the synergistic mechanism of a multicomponent herbal formula, Baoyuan decoction against cardiac hypertrophy. Biomed. Pharmacother. 2021, 139, 111665. [Google Scholar] [CrossRef]
- Chan, A.W.; Gill, R.S.; Schiller, D.; Sawyer, M.B. Potential role of metabolomics in diagnosis and surveillance of gastric cancer. World J. Gastroenterol. 2014, 20, 12874–12882. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Yu, H.; Zhao, X.; Bao, Y.; Hong, C.S.; Zhang, P.; Tu, Y.; Yin, P.; Gao, P.; Wei, L.; et al. Metabolomics study of Roux-en-Y Gastric Bypass Surgery (RYGB) to treat Type 2 diabetes patients based on Ultraperformance Liquid Chromatography-Mass Spectrometry. J. Proteome Res. 2016, 15, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, H.; Yang, L.; Jiang, H.; Guo, S.; Li, Y.; Tao, S. Construction of a metabolomics profile of arsenic trioxide effect in gastric carcinoma cell line SGC7901. Acta Biochim. Biophys. Sin. 2016, 48, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Koushki, M.; Farrokhi Yekta, R.; Amiri-Dashatan, N.; Dadpay, M.; Goshadrou, F. Therapeutic effects of hydro-alcoholic extract of Achillea wilhelmsii C. Koch on indomethacin-induced gastric ulcer in rats: A proteomic and metabolomic approach. BMC Complement. Altern. Med. 2019, 19, 205. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, B.; Mokhtar, F.A.; El-Masry, T.A.; Elekhnawy, E.; Mostafa, S.A.; Abdelkader, D.H.; Elharty, M.E.; Saleh, A.; Negm, W.A. Antimicrobial activity of Brassica rapa L. flowers extract on gastrointestinal tract infections and antiulcer potential against indomethacin-induced gastric ulcer in rats supported by metabolomics profiling. J. Inflamm. Res. 2021, 14, 7411–7430. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Bao, T.; Bai, L.; Mu, X.; Ta, N.; Bao, M.; Li, Y.; Zhang, J.; Fu, M.; Chen, Y. Mongolian medicine formulae Ruda-6 alleviates indomethacin-induced gastric ulcer by regulating gut microbiome and serum metabolomics in rats. J. Ethnopharmacol. 2023, 314, 116545. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Z.; Liu, Y.H.; Liang, J.L.; Huang, Q.H.; Dou, Y.X.; Nie, J.; Zhuo, J.Y.; Wu, X.; Chen, J.N.; Su, Z.R.; et al. Protective role of beta-patchoulene from Pogostemon cablin against indomethacin-induced gastric ulcer in rats: Involvement of anti-inflammation and angiogenesis. Phytomedicine 2018, 39, 111–118. [Google Scholar] [CrossRef]
- Gomaa, A.M.S.; Abd El-Mottaleb, N.A.; Aamer, H.A. Antioxidant and anti-inflammatory activities of alpha lipoic acid protect against indomethacin-induced gastric ulcer in rats. Biomed. Pharmacother. 2018, 101, 188–194. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Li, X.; Dai, Y.; Wang, Q.; Wang, G.; Liu, D.; Gu, X.; Yu, D.; Ma, Y.; et al. Treatment mechanism of gardeniae fructus and its carbonized product against ethanol-induced gastric lesions in rats. Front. Pharmacol. 2019, 10, 750. [Google Scholar] [CrossRef]
- Ren, S.; Wei, Y.; Niu, M.; Li, R.; Wang, R.; Wei, S.; Wen, J.; Wang, D.; Yang, T.; Chen, X.; et al. Mechanism of rutaecarpine on ethanol-induced acute gastric ulcer using integrated metabolomics and network pharmacology. Biomed. Pharmacother. 2021, 138, 111490. [Google Scholar] [CrossRef]
- Pineda-Pena, E.A.; Martinez-Perez, Y.; Galicia-Moreno, M.; Navarrete, A.; Segovia, J.; Muriel, P.; Favari, L.; Castaneda-Hernandez, G.; Chavez-Pina, A.E. Participation of the anti-inflammatory and antioxidative activity of docosahexaenoic acid on indomethacin-induced gastric injury model. Eur. J. Pharmacol. 2018, 818, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Nolasco, C.; Barragán-Zarate, G.S.; Lagunez-Rivera, L.; Solano, R.; Favari, L.; Jiménez-Andrade, J.M.; Chávez-Piña, A.E. Role of LTB4 and nitric oxide in the gastroprotective effect of Prosthechea karwinskii leaves extract in the indomethacin-induced gastric injury in the rat. Nat. Prod. Res. 2022, 37, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Peña, E.A.; Capistran-Amezcua, D.; Reyes-Ramírez, A.; Xolalpa-Molina, S.; Chávez-Piña, A.E.; Figueroa, M.; Navarrete, A. Gastroprotective effect methanol extract of Caesalpinia coriaria pods against indomethacin- and ethanol-induced gastric lesions in Wistar rats. J. Ethnopharmacol. 2023, 305, 116057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Shi, Y.; Yang, C.; Lu, S.; Zhao, L.; Liu, X.; Zhou, D.; Luo, L.; Yin, Z. γ-glutamylcysteine alleviates insulin resistance and hepatic steatosis by regulating adenylate cyclase and IGF-1R/IRS1/PI3K/Akt signaling pathways. J. Nutr. Biochem. 2023, 119, 109404. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lupton, J.R.; Turner, N.D.; Fang, Y.-Z.; Yang, S. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, R.; Liu, H.; Wang, L.; Li, J.; Wu, S.; Chen, X.; Yang, X.; Zhao, Y. Berberine regulates macrophage polarization through IL-4-STAT6 signaling pathway in Helicobacter pylori-induced chronic atrophic gastritis. Life Sci. 2021, 266, 118903. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Hou, X.; Zeng, M.; Chen, X.; Chen, X.; Yang, T.; Xu, F.; Peng, J.; Peng, Q.; Cai, X.; et al. Traditional Chinese Medicine Li-Zhong-Tang accelerates the healing of indomethacin-induced gastric ulcers in rats by affecting TLR-2/MyD88 signaling pathway. J. Ethnopharmacol. 2020, 259, 112979. [Google Scholar] [CrossRef]
- Barboza, K.R.M.; Coco, L.Z.; Alves, G.M.; Peters, B.; Vasquez, E.C.; Pereira, T.M.C.; Meyrelles, S.S.; Campagnaro, B.P. Gastroprotective effect of oral kefir on indomethacin-induced acute gastric lesions in mice: Impact on oxidative stress. Life Sci. 2018, 209, 370–376. [Google Scholar] [CrossRef]
- Rasmussen, J.A.; Villumsen, K.R.; Ernst, M.; Hansen, M.; Forberg, T.; Gopalakrishnan, S.; Gilbert, M.T.P.; Bojesen, A.M.; Kristiansen, K.; Limborg, M.T. A multi-omics approach unravels metagenomic and metabolic alterations of a probiotic and synbiotic additive in rainbow trout (Oncorhynchus mykiss). Microbiome 2022, 10, 21. [Google Scholar] [CrossRef]
- Navarro-Reig, M.; Jaumot, J.; García-Reiriz, A.; Tauler, R. Evaluation of changes induced in rice metabolome by Cd and Cu exposure using LC-MS with XCMS and MCR-ALS data analysis strategies. Anal. Bioanal. Chem. 2015, 407, 8835–8847. [Google Scholar] [CrossRef]
- Ma, X.; Xie, S.; Wang, R.; Wang, Z.; Jing, M.; Li, H.; Wei, S.; Liu, H.; Li, J.; He, Q.; et al. Metabolomics profiles associated with the treatment of Zuojin Pill on patients with chronic nonatrophic gastritis. Front. Pharmacol. 2022, 13, 898680. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Luo, Z.; Li, D.; Qin, J.; Pan, Z.; Guo, B.; Deng, L.; Nong, Y.; Huang, Z.; He, Y.; et al. Investigation of the therapeutic effect of total alkaloids of Corydalis saxicola Bunting on CCl4-induced liver fibrosis in rats by LC/MS-Based metabolomics analysis and network pharmacology. Metabolites 2022, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, Y.; Jiang, H.; Yang, J.; Wang, Q.; Du, Y.; Xu, H. Integrated strategy for accurately screening biomarkers based on metabolomics coupled with network pharmacology. Talanta 2020, 211, 120710. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolism | m/z | Formula | Changed Trend | |
|---|---|---|---|---|---|
| Model/Control | Model/Berberine | ||||
| 1 | d-Synephrine | 168.0918 | C9H13NO2 | Up # | Down ** |
| 2 | l-2-Amino-6-oxoheptanedioate | 190.0863 | C7H11NO5 | Up ### | Down * |
| 3 | Butyryl-l-carnitine | 232.1549 | C11H21NO4 | Up ### | Down * |
| 4 | Estradiol | 273.2536 | C18H24O2 | Down # | Up * |
| 5 | Dehydroepiandrosterone | 288.2896 | C19H28O2 | Down # | Up * |
| 6 | Epipregnanolone | 319.2681 | C21H34O2 | Down # | Up * |
| 7 | 12-Keto-tetrahydro-leukotriene B4 | 336.3267 | C20H32O4 | Down ## | Up ** |
| 8 | (R)-3-Hydroxybutyric acid | 103.0397 | C4H8O3 | Down ## | Up * |
| 9 | 3-Methylthiopropionic acid | 118.9269 | C4H8O2S | Up # | Down * |
| 10 | Dodecanedioic acid | 229.1447 | C12H22O4 | Down ## | Up * |
| 11 | γ-Glutamylcysteine | 248.9601 | C8H14N2O5S | Up ### | Down ** |
| 12 | Stearic acid | 283.2605 | C18H36O2 | Down # | Up * |
| 13 | Corticosterone | 345.2060 | C21H30O4 | Down # | Up * |
| 14 | Indoleacetaldehyde | 160.0758 | C10H9NO | Down # | Up * |
| Time (min) | Positive Mode | Negative Mode | ||
|---|---|---|---|---|
| A (%) | B (%) | A (%) | B (%) | |
| 0–1 | 92 | 8 | 92 | 8 |
| 1–8 | 92–2 | 8–98 | 92–2 | 89–8 |
| 8–10 | 2 | 98 | 2 | 98 |
| 10–10.1 | 29–2 | 98–8 | 2–92 | 98–8 |
| 10.1–12 | 92 | 8 | 92 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Yang, X.-W. LC-MS-Based Metabolomics Reveals the Mechanism of Protection of Berberine against Indomethacin-Induced Gastric Injury in Rats. Molecules 2024, 29, 1055. https://doi.org/10.3390/molecules29051055
Xu J, Yang X-W. LC-MS-Based Metabolomics Reveals the Mechanism of Protection of Berberine against Indomethacin-Induced Gastric Injury in Rats. Molecules. 2024; 29(5):1055. https://doi.org/10.3390/molecules29051055
Chicago/Turabian StyleXu, Jing, and Xiu-Wei Yang. 2024. "LC-MS-Based Metabolomics Reveals the Mechanism of Protection of Berberine against Indomethacin-Induced Gastric Injury in Rats" Molecules 29, no. 5: 1055. https://doi.org/10.3390/molecules29051055
APA StyleXu, J., & Yang, X.-W. (2024). LC-MS-Based Metabolomics Reveals the Mechanism of Protection of Berberine against Indomethacin-Induced Gastric Injury in Rats. Molecules, 29(5), 1055. https://doi.org/10.3390/molecules29051055