
Different Types of Hypericum perforatum cvs. (Elixir, Helos, Topas) In Vitro Cultures: A Rich Source of Bioactive Metabolites and Biological Activities of Biomass Extracts

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results and Discussion
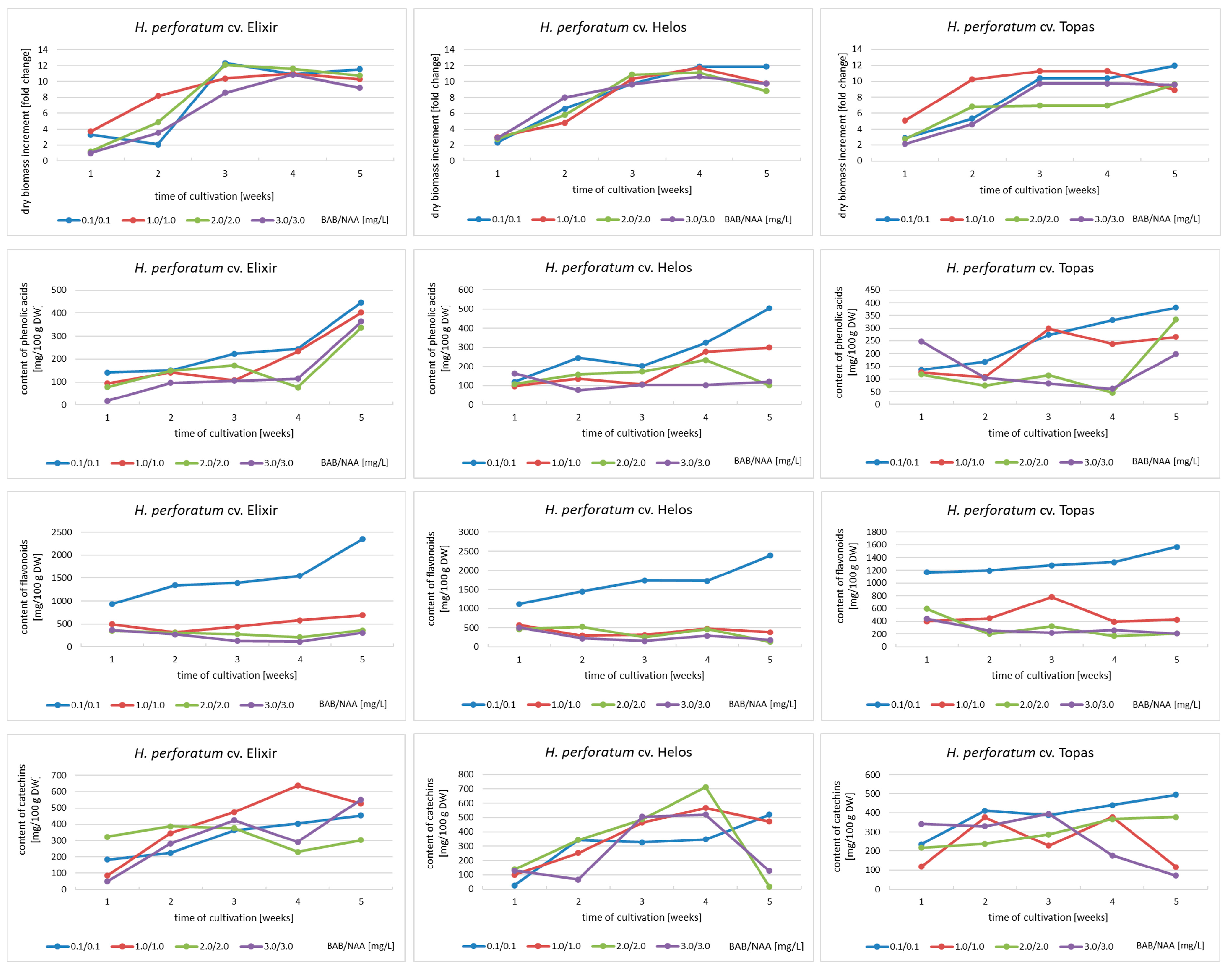
2.1. Dynamics of Metabolite Accumulation in Agitated Cultures
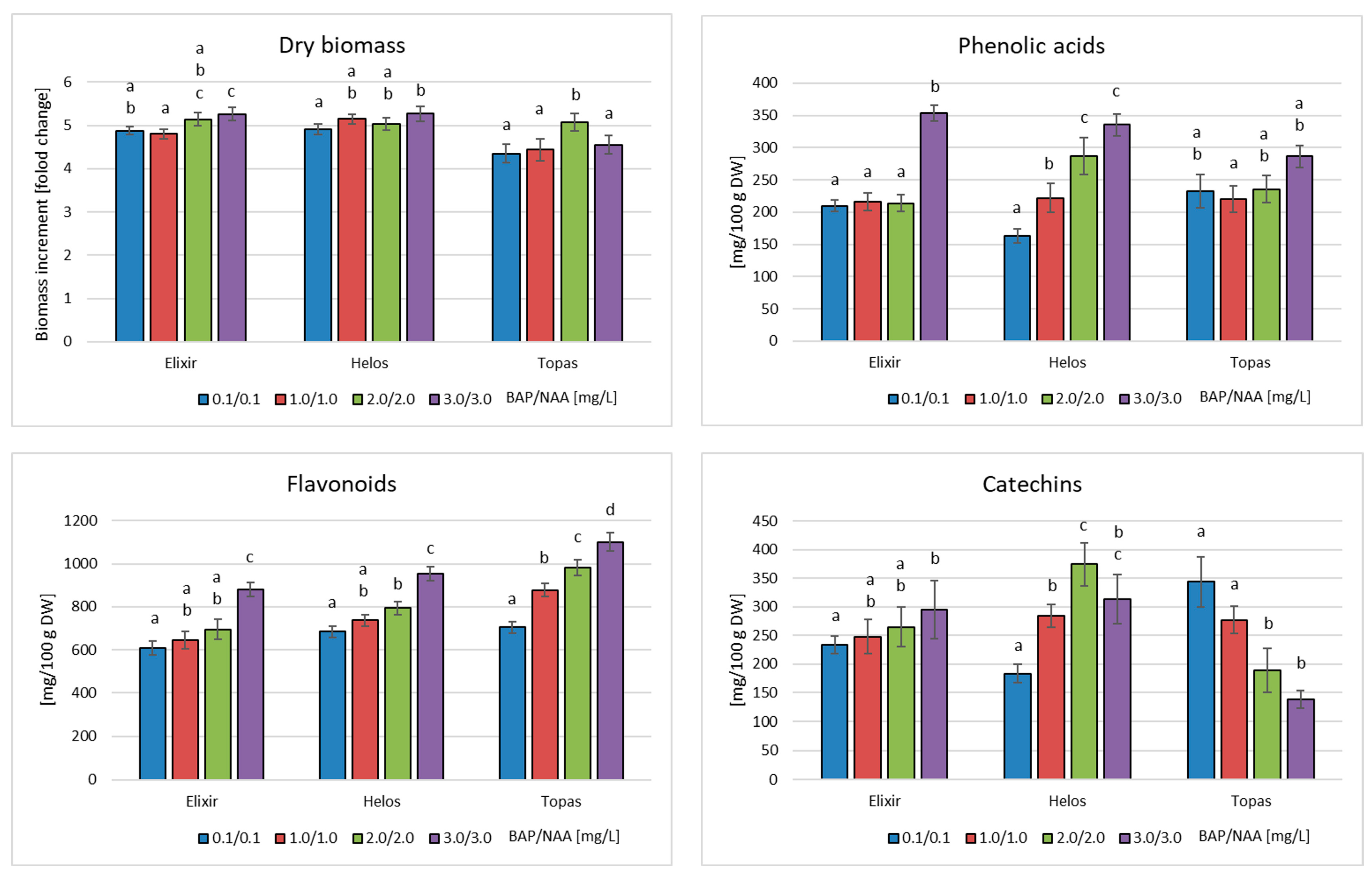
2.2. Cultures in Bioreactors
2.3. Total Phenolic, Flavonoid, and Condensed Tannin Contents of Selected Extracts
2.4. Activity of Selected Extracts
2.4.1. Antioxidant Activity
2.4.2. Artemia salina Lethality Bioassay
2.4.3. Antibacterial Activity
2.4.4. Antifungal Activity
2.5. Enhancing the Production of Secondary Metabolites by Phenylalanine Feeding
3. Materials and Methods
3.1. In Vitro Initial Cultures
3.2. Agitated Cultures
3.3. Cultures in Bioreactors
3.4. Feeding Culture Medium with Phenylalanine as the Biogenetic Precursor
3.5. Extracts Preparation
3.6. Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis
3.7. Total Phenolic, Flavonoid, and Condensed Tannin Contents
3.8. Antioxidant Activity
3.8.1. Free Radical Scavenging Activity
3.8.2. Reducing Power Assay
3.8.3. Ferrous Ions (Fe2+) Chelating Activity
3.9. Artemia Salina Lethality Bioassay
3.10. Antimicrobial Activity
3.10.1. Antibacterial Activity
3.10.2. Antifungal Activity
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| ASE | Ascorbic acid equivalent |
| BAP | 6-Benzylaminopurine |
| BHT | Butylated hydroxytoluene |
| DPPH | 1,1-Diphenyl-2-picrylhydrazyl |
| DW | Dry weight |
| GAE | Gallic acid equivalents |
| HAT | Hydrogen-atom transfer |
| MIC | Minimum Inhibitory Concentration |
| MS | Murashige and Skoog |
| NAA | 1-Naphthaleneacetic acid |
| PGR | Plant growth regulator |
| QE | Quercetin equivalents |
| SET | Single-electron transfer |
| TIS | Temporary immersion system |
References
- Kwiecień, I.; Nicosia, N.; Ekiert, H. Cultivation of Hypericum Perforatum (St. John’s Wort) and Biotechnological Approaches for Improvement of Plant Raw Material Quality. In Medicinal Plants. Sustainable Development and Biodiversity; Ekiert, H.M., Ramawat, K.G., Arora, J., Eds.; Springer: Cham, Switzerland, 2021; Volume 28, pp. 253–291. [Google Scholar]
- European Pharmacopoeia 10.0. European Directorate for the Quality of Medicines: Strasburg, France, 2020.
- Barnes, J.; Anderson, L.; St. Phillipson, J. John’s wort (Hypericum perforatum L.): A review of its chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2001, 53, 583–600. [Google Scholar] [CrossRef]
- ESCOP. Hyperici Herba—St. John’s Wort; European Scientific Cooperative on Phytotherapy Monographs: Exeter, UK, 2018. [Google Scholar]
- Apaydin, E.A.; Maher, A.R.; Shanman, R.; Booth, M.S.; Miles, J.N.V.; Sobero, M.E.; Hempel, S. A systematic review of St. John’s wort for major depressive disorder. Syst. Rev. 2016, 5, 148. [Google Scholar] [CrossRef]
- Khan, A.-U.; Gilani, A.-H.; Najeeb-ur-Rehman. Pharmacological studies on Hypericum perforatum fractions and constituents. Pharm. Biol. 2011, 49, 46–56. [Google Scholar] [CrossRef]
- Quave, C.L. Wound healing with botanicals: A review and future perspectives. Curr. Dermatol. Rep. 2018, 7, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Saddiqe, Z.; Naeem, I.; Maimoona, A. A review of the antibacterial activity of Hypericum perforatum L. J. Ethnopharmacol. 2010, 131, 511–521. [Google Scholar] [CrossRef]
- World Health Organization. Herba hyperici. In WHO Monographs on Selected Medicinal Plants; World Health Organization: Geneva, Switzerland, 2004; Volume 2, pp. 149–171. [Google Scholar]
- European Medicines Agency (EMA). Community Herbal Monograph on Hypericum Perforatum L. Herba (Traditional Use); Committee on Herbal Medicinal Products: London, UK, 2009. [Google Scholar]
- Agapouda, A.; Booker, A.; Kiss, T.; Hohmann, J.; Heinrich, M.; Csupor, D. Quality control of Hypericum perforatum L. analytical challenges and recent progress. J. Pharm. Pharmacol. 2019, 71, 15–37. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Murthy, H.N.; Lee, E.-J.; Paek, K.-Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Coste, A.; Pop, C.; Halmagyi, A.; Butiuc-Keul, A. Secondary Metabolites in Shoot Cultures of Hypericum. In Plant Cell and Tissue Differentiation and Secondary Metabolites; Reference Series in Phytochemistry; Ramawat, K., Ekiert, H., Goyal, S., Eds.; Springer Nature: Geneva, Switzerland, 2021; pp. 273–308. [Google Scholar]
- Kwiecień, I.; Szydłowska, A.; Kawka, B.; Beerhues, L.; Ekiert, H. Accumulation of biologically active phenolic acids in agitated shoot cultures of three Hypericum perforatum cultivars: ‘Elixir’, ‘Helos’ and ‘Topas’. Plant Cell Tissue Organ Cult. 2015, 123, 273–281. [Google Scholar] [CrossRef]
- Kwiecień, I.; Smolin, J.; Beerhues, L.; Ekiert, H. The impact of media composition on production of flavonoids in agitated shoot cultures of the three Hypericum perforatum L. cultivars ‘Elixir,’ ‘Helos,’ and ‘Topas’. In Vitro Cell. Dev. Biol. - Plant 2018, 54, 332–340. [Google Scholar] [CrossRef]
- Muszyńska, B.; Ekiert, H.; Kwiecień, I.; Maślanka, A.; Zodi, R.; Beerhues, L. Comparative analysis of therapeutically important indole compounds in in vitro cultures of Hypericum perforatum cultivars by HPLC and TLC analysis coupled with densitometric detection. Nat. Prod. Commun. 2014, 9, 1437–1440. [Google Scholar] [CrossRef]
- Jürgenliemk, C.; Nahrstedt, A. Phenolic compound from Hypericum perforatum. Planta Med. 2002, 68, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Bruni, R.; Sacchetti, G. Factors affecting polyphenol biosynthesis in wild and filed grown St John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682–725. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.; Dias, A.C.P.; Seabra, R.M.; Andrade, P.B.; Fernandes-Ferreira, M. The development and evaluation of an HPLC-DAD method for the analysis of the phenolic fractions from in vivo and in vitro biomass of Hypericum species. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 215–227. [Google Scholar] [CrossRef]
- Bertoli, A.; Giovannini, A.; Ruffoni, B.; Guardo, A.D.; Spinelli, G.; Mazzetti, M.; Pistelli, L. Bioactive constituent production in St. John’s Wort in vitro hairy roots. Regenerated plant lines. J. Agric. Food Chem. 2008, 56, 5078–5082. [Google Scholar] [CrossRef]
- Pasqua, G.; Avato, P.; Monacelli, B.; Santamari, A.R. Metabolites in cell suspension cultures, calli, and in vitro regenerated organs of Hypericum perforatum cv. Topas. Plant Sci. 2003, 165, 977–982. [Google Scholar] [CrossRef]
- Savio, L.E.B.; Astarita, L.V.; Santarém, E.R. Secondary metabolism in micropropagated Hypericum perforatum L. grown in nonaerated liquid medium. Plant Cell Tissue Organ Cult. 2012, 108, 465–472. [Google Scholar] [CrossRef]
- Čellárová, E. Effect of exogenous morphogenetic signals on differentiation in vitro and secondary metabolite formation in the genus Hypericum. Med. Aromat. Plant Sci. Biotechnol. 2011, 5, 62–69. [Google Scholar]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant growth regulator mediated consequences of secondary metabolites in medicinal plants. J. App. Res. Med. Arom. Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Shakya, P.; Marslin, G.; Siram, K.; Beerhues, L.; Franklin, G. Elicitation as a tool to improve the profiles of high-value secondary metabolites and pharmacological properties of Hypericum perforatum. J. Pharm. Pharmacol. 2019, 71, 70–82. [Google Scholar] [CrossRef]
- Gadzovska, S.; Maury, S.; Ounnar, S.; Righezza, M.; Kascakova, S.; Refregiers, M.; Spasenoski, M.; Joseph, C.; Hagège, D. Identification and quantification of hypericin and pseudohypericin in different Hypericum perforatum L. in vitro cultures. Plant Physiol. Biochem. 2005, 43, 591–601. [Google Scholar] [CrossRef]
- Treneva, G.; Markovska, Y.; Wolfram, E.; Danova, K. Effect of plant growth regulators on growth patterns and enzymatic antioxidant activities in Hypericum calycinum shoot cultures. Bulg. J. Agr. Sci. 2014, 20, 46–50. [Google Scholar] [CrossRef]
- Zobayed, S.; Murch, S.; Rupasinghe, H.; Saxena, P.K. Elevated carbon supply altered hypericin and hyperforin contents of St. John’s wort (Hypericum perforatum L. cv ‘New Stem’) grown in bioreactors. Plant Cell Tissue Organ Cult. 2003, 75, 143–149. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Murch, S.J.; Rupasinghe, H.P.V.; Saxena, P.K. In vitro production and chemical characterization of St. John’s wort (Hypericum perforatum L. cv ‘New Stem’), Plant Sci. 2004, 166, 333–340. [Google Scholar]
- Cui, X.H.; Murthy, H.N.; Jin, Y.X.; Yim, Y.H.; Kim, J.Y.; Paek, K.Y. Production of adventitious root biomass and secondary metabolites of Hypericum perforatum L. in a balloon type airlift reactor. Bioresour. Technol. 2011, 102, 10072–10079. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Q.; Yu, X.K.; Lian, M.L.; Park, S.Y.; Piao, X.C. Several factors affecting hypericin production of Hypericum perforatum during adventitious root culture in airlift bioreactors. Acta Physiol. Plant. 2014, 36, 975–981. [Google Scholar] [CrossRef]
- Cui, X.H.; Murthy, H.N.; Paek, K.Y. Pilot-Scale Culture of Hypericum Perforatum, L. Adventitious Roots in Airlift Bioreactors for the Production of Bioactive Compounds. Appl. Biochem. Biotechnol. 2014, 174, 784–792. [Google Scholar] [CrossRef]
- Mirzabe, A.H.; Hajiahmad, A.; Fadavi, A.; Rafiee, S. Temporary immersion systems (TISs): A comprehensive review. J. Biotechnol. 2022, 357, 56–83. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Bednarz, M.; Jafernik, K.; Luczkiewicz, M.; Ekiert, H. Bioreactor type affects the accumulation of phenolic acids and flavonoids in microshoot cultures of Schisandra chinensis (Turcz.) Baill. Plant Cell Tissue Organ Cult. 2019, 139, 199–206. [Google Scholar] [CrossRef]
- Kubica, P.; Szopa, A.; Kokotkiewicz, A.; Miceli, N.; Taviano, M.F.; Maugeri, A.; Cirmi, S.; Synowiec, A.; Gniewosz, M.; Elansary, H.O.; et al. Production of Verbascoside, Isoverbascoside and Phenolic Acids in Callus, Suspension, and Bioreactor Cultures of Verbena officinalis and Biological Properties of Biomass Extracts. Molecules 2020, 25, 5609. [Google Scholar] [CrossRef]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Production of specific flavonoids and verbascoside in shoot cultures of Scutellaria baicalensis. In Plant Cell and Tissue Differentiation and Secondary Metabolites; Reference Series in Phytochemistry; Ramawat, K., Ekiert, H., Goyal, S., Eds.; Springer Nature: Geneva, Switzerland, 2021; pp. 249–272. [Google Scholar]
- Kwiecień, I.; Miceli, N.; D’Arrigo, M.; Marino, A.; Ekiert, H. Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods. Molecules 2022, 27, 1140. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Klimek-Szczykutowicz, M.; Kokotkiewicz, A.; Dziurka, M.; Luczkiewicz, M.; Ekiert, H. Phenolic acid and flavonoid production in agar, agitated and bioreactor-grown microshoot cultures of Schisandra chinensis cv. Sadova No. 1—A valuable medicinal plant. J. Biotechnol. 2019, 305, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. 2005, 579, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, A.; Kwiecień, I.; Grabowski, M.; Rajek, K.; Cavò, E.; Taviano, M.F.; Miceli, N. Phenylalanine Increases the Production of Antioxidant Phenolic Acids in Ginkgo biloba Cell Cultures. Molecules 2021, 26, 4965. [Google Scholar] [CrossRef]
- Ekiert, H.; Kubica, P.; Kwiecień, I.; Jafernik, K.; Klimek-Szczykutowicz, M.; Szopa, A. Cultures of Medicinal Plants In Vitro as a Potential Rich Source of Antioxidants. In Plant Antioxidants and Health; Reference Series in Phytochemistry; Ekiert, H., Ramawat, K., Arora, J., Eds.; Springer Nature: Geneva, Switzerland, 2022; pp. 267–309. [Google Scholar]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Zhang, S.; Gai, Z.; Gui, T.; Chen, J.; Chen, Q.; Li, Y. Antioxidant Effects of Protocatechuic Acid and Protocatechuic Aldehyde: Old Wine in a New Bottle. Evid. Based Complement. Alternat. Med. 2021, 2021, 6139308. [Google Scholar] [CrossRef]
- Wang, L.; Pan, X.; Jiang, L.; Chu, Y.; Gao, S.; Jiang, X.; Zhang, Y.; Chen, Y.; Luo, S.; Peng, C. The Biological Activity Mechanism of Chlorogenic Acid and Its Applications in Food Industry: A Review. Front. Nutr. 2022, 9, 943911. [Google Scholar] [CrossRef]
- Silva, B.A.; Malva, J.O.; St. Dias, A. John’s Wort (Hypericum perforatum) extracts and isolated phenolic compounds are effective antioxidants in several in vitro models of oxidative stress. Food Chem. 2008, 110, 611–619. [Google Scholar] [CrossRef]
- Orčić, D.Z.; Mimica-Dukić, N.M.; Francišković, M.M.; Petrović, S.S.; Jovin, E.D. Antioxidant activity relationship of phenolic compounds in Hypericum perforatum L. Chem. Cent. J. 2011, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Siddhuraju, P.; Manian, S. The antioxidant activity and free radical-scavenging capacity of dietary phenolic extracts from horse gram (Macrotyloma uniflorum (Lam.) Verdc.) seeds. Food Chem. 2007, 105, 950–958. [Google Scholar] [CrossRef]
- Borg, D.C. Oxygen free radicals and tissue injury. In Oxygen Free Radicals in Tissue Damage, 1st ed.; Tarr, M., Samson, F., Eds.; Birkhauser: Boston, MA, USA, 1993; pp. 12–53. [Google Scholar]
- Lopes, G.K.B.; Schulman, H.M.; Hermes-Lima, M. Polyphenol tannic acid inhibits hydroxyl radical formation from Fenton reaction by complexing ferrous ions. Biochim. Biophys. Acta. 1999, 1472, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.G., Jr.; Dalvi, L.T.; Silva, J.M.C.; Lopes, G.K.B.; Alonso, A.; Hermes-Lima, M. The antioxidant effect of tannic acid on the in vitro copper-mediated formation of free radicals. Arch. Biochem. Biophys. 2005, 437, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovič, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Rajabi, S.; Ramazani, A.; Hamidi, M.; Naji, T. Artemia salina as a model organism in toxicity assessment of nanoparticles. DARU J. Pharm. Sci. 2015, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Lagarto Parra, A.; Silva Yhebra, R.; Guerra Sardiñas, I.; Iglesias Buela, L. Comparative study of the assay of Artemia salina L. and the estimate of the medium lethal dose (LD50 value) in mice, to determine oral acute toxicity of plant extracts. Phytomedicine 2001, 8, 395–400. [Google Scholar] [CrossRef]
- Mirmalek, S.A.; Azizi, M.A.; Jangholi, E.; Yadollah-Damavandi, S.; Javidi, M.A.; Parsa, Y.; Parsa, T.; Salimi-Tabatabaee, S.A.; Ghasemzadeh Kolagar, H.; Alizadeh-Navaei, R. Cytotoxic and apoptogenic effect of hypericin, the bioactive component of Hypericum perforatum on the MCF-7 human breast cancer cell line. Cancer Cell Int. 2016, 16, 3. [Google Scholar] [CrossRef]
- Peron, A.P.; Mariucci, R.G.; de Almeida, I.V.; Düsman, E.; Mantovani, M.S.; Vicentini, V.E.P. Evaluation of the cytotoxicity, mutagenicity and antimutagenicity of a natural antidepressant, Hypericum perforatum L. (St. John’s wort), on vegetal and animal test systems. BMC Complement. Altern. Med. 2013, 13, 97. [Google Scholar] [CrossRef] [PubMed]
- Rancic, A.; Sokovic, M.; Vukojevic, J.; Simic, A.; Marin, P.; Duletic-Lausevic, S.; Djokovic, D. Chemical composition and antimicrobial activities of essential oils of Myrrhis odorata (L.) Scop., Hypericum perforatum (L.) and Helichrysum arenarium (L.) Moench. J. Essent. Oil Res. 2005, 17, 341–345. [Google Scholar] [CrossRef]
- Radulovic, N.; Stankov-Jovanovic, V.; Stojanovic, G.; Smelcerovic, A.; Spiteller, M.; Asakawa, Y. Screening of in vitro antimicrobial and antioxidant activity of nine Hypericum species from the Balkans. Food Chem. 2007, 103, 15–21. [Google Scholar] [CrossRef]
- Saroglou, V.; Marin, P.D.; Rancic, A.; Veljic, M.; Skaltsa, H. Composition and antimicrobial activity of the essential oil of six Hypericum species from Serbia. Biochem. Syst. Ecol. 2007, 35, 146–152. [Google Scholar] [CrossRef]
- Brondz, I.; Greibrokk, T.; Growth, P.A.; Aasen, A.J. The relative stereochemistry of hyperforin-an antibiotic from Hypericum perforatum L. Tetrahedron Lett. 1982, 23, 1299–1300. [Google Scholar] [CrossRef]
- Mullaicharam, A.R.; Halligudi, N. St John’s wort (Hypericum perforatum L.): A Review of its Chemistry, Pharmacology and Clinical properties. Int. J. Res. Phy. Pharm. Sci. 2018, 1, 5–11. [Google Scholar] [CrossRef]
- Reichling, J.; Weseler, A.; Saller, R. A current review of the antimicrobial activity of Hypericum perforatum L. Pharmacopsychiatry 2001, 34 (Suppl. S1), 116–118. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Statti, G.; Conforti, F. Hypericum Spp.: An Update on The Biological Activities And Metabolic Profiles. Mini-Rev. Med. Chem. 2020, 20, 66–87. [Google Scholar] [CrossRef]
- Schempp, C.M.; Pelz, K.; Wittmer, A.; Schöpf, E.; Simon, J.C. Antibacterial activity of hyperforin from St John’s wort, against multiresistant Staphylococcus aureus and gram-positive bacteria. Lancet 1999, 353, 2129. [Google Scholar] [CrossRef] [PubMed]
- Mazandarani, M.; Yassaghi, S.; Rezaei, M.B.; Mansourian, A.R.; Ghaemi, E.O. Ethnobotany and antibacterial activities of two endemic species of Hypericum in North-East of Iran. Asian J. Plant Sci. 2007, 6, 354–358. [Google Scholar] [CrossRef]
- Avato, P.; Raffo, F.; Guglielmi, G.; Vitali, C.; Rosato, A. Extracts from St John’s Wort and their antimicrobial Activity. Phytother. Res. 2004, 18, 230–232. [Google Scholar] [CrossRef]
- Barbagallo, C.; Chisari, G. Antimicrobial activity of three Hypericum species. Fitoterapia 1987, 58, 175–177. [Google Scholar]
- Sytar, O.; Švedienė, J.; Ložienė, K.; Paškevičius, A.; Kosyan, A.; Taran, N. Antifungal properties of hypericin, hypericin tetrasulphonic acid and fagopyrin on pathogenic fungi and spoilage yeasts. Pharm. Biol. 2016, 54, 3121–3125. [Google Scholar] [CrossRef]
- Milosevic, T.; Solujic, S.; Sukdolak, S. In Vitro Study of Ethanolic Extract of Hypericum perforatum L. on Growth and Sporulation of Some Bacteria and Fungi. Turk. J. Biol. 2007, 31, 237–241. [Google Scholar]
- Maskovic, P.; Solujic, S. A preliminary study on the antifungal activity of the ethanol extract of Hyperiucm perforatum L. The book of abstracts. In Proceedings of the Eleventh Annual Conference “YUCOMAT 2009”, Herceg-Novi, Serbia, 31 August–4 September 2009; p. 1922009. [Google Scholar]
- Liu, X.N.; Zhang, X.Q.; Zhang, S.X.; Sun, J.S. Regulation of metabolite production by precursors and elicitors in liquid cultures of Hypericum perforatum. Plant Cell Tissue Organ Cult. 2007, 91, 1–7. [Google Scholar] [CrossRef]
- Karppinen, K.; Hokkanen, J.; Tolonen, A.; Mattila, S.; Hohtola, A. Biosynthesis of hyperforin and adhyperforin from amino acid precursors in shoot cultures of Hypericum perforatum. Phytochemistry 2007, 68, 1038–1045. [Google Scholar] [CrossRef]
- Skrzypczak-Pietraszek, E.; Słota, J.; Pietraszek, J. The influence of L-phenylalanine, methyl jasmonate and sucrose concentration on the accumulation of phenolic acids in Exacum affine Balf. f. ex Regel shoot culture. Acta Biochim. Pol. 2014, 61, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Kubica, P.; Komsta, Ł.; Walkowicz-Bożek, A.; Ekiert, H. The effect of feeding culture media with biogenetic precursors on high production of depsides in agitated shoot cultures of black and red aronias. Plant Cell Tissue Organ Cult. 2020, 142, 379–399. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blažević, I.; Đulović, A.; Miazga-Karska, M.; Klimek, K.; Ekiert, H.; Szopa, A. Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts. Molecules 2021, 26, 4660. [Google Scholar] [CrossRef]
- Sajjalaguddam, R.R.; Paladugu, A. Phenylalanine enhances Quercetin content in In vitro cultures of Abutilon indicum L. J. Appl. Pharm. Sci. 2015, 5, 80–84. [Google Scholar] [CrossRef]
- Biswas, T.; Mathur, A.; Gupta, V.; Luqman, S.; Mathur, A.K. Elicitation and phenylalanine precursor feeding based modulation of in vitro anthocyanin production, enzyme activity and gene expression in an Indian ginseng congener-Panax sikkimensis Ban. Ind. Crops Prod. 2020, 145, 111986. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ellnain-Wojtaszek, M.; Zgórka, G. High-performance liquid chromatography and thin-layer chromatography of phenolic acids from Ginkgo biloba L. leaves collected within vegetative period. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 1457–1471. [Google Scholar] [CrossRef]
- Gao, X.; Ohlander, M.; Jeppsson, N.; Björk, L.; Trajkovski, V. Changes in antioxidant effects and their relationship to phytonutrients in fruits of sea buckthorn (Hippophae rhamnoides L.) during maturation. J. Agric. Food Chem. 2000, 48, 1485–1490. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug. Anal. 2002, 10, 178–182. [Google Scholar]
- Julkunen-Titto, R. Phenolic constituents in the leaves of northern willows: Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Ohnishi, M.; Morishita, H.; Iwahashi, H.; Shitzuo, T.; Yoshiaki, S.; Kimura, M.; Kido, R. Inhibitory effects of chlorogenic acid on linoleic acid, peroxidation and haemolysis. Phytochemistry 1994, 36, 579–583. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Taviano, M.F.; Rashed, K.; Filocamo, A.; Cacciola, F.; Dugo, P.; Mondello, L.; Bisignano, C.; Acquaviva, R.; D’Arrigo, M.; Miceli, N. Phenolic profile and biological properties of the leaves of Ficus vasta Forssk. (Moraceae) growing in Egypt. BMC Complement. Altern. Med. 2018, 18, 161–171. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. perforatum Extracts | Polyphenol Content mg GAE/g Extract (DW) | Flavonoid Content mg QE/g Extract (DW) | Condensed Tannins mg CE/g Extract (DW) |
|---|---|---|---|
| cv. Helos | 75.99 ± 0.19 a | 23.21 ± 0.29 a | 19.85 ± 0.36 a |
| cv. Elixir | 69.06 ± 1.17 b | 26.86 ± 0.22 a | 6.21 ± 0.23 b |
| cv. Topas | 75.49 ± 0.34 a | 48.33 ± 0.27 b | 16.31 ± 0.96 a |
| H. perforatum Extracts | DPPH Test IC50 (mg/mL) | Reducing Power Assay ASE/mL | Fe2+ Chelating Activity IC50 (mg/mL) |
|---|---|---|---|
| cv. Helos | 0.586 ± 0.062 a | 16.429 ± 0.229 a | 0.492 ± 0.012 a |
| cv. Elixir | 1.625 ± 0.048 b | 12.942 ± 0.839 b | 0.530 ± 0.100 b |
| cv. Topas | 0.512 ± 0.036 c | 20.765 ± 0.001 c | 0.505 ± 0.015 b |
| Standard | BHT 0.0656 ± 0.008 d | BHT 1.131 ± 0.037 d | EDTA 0.0067 ± 0.0003 c |
| Species of Microorganisms | Antibacterial Activity (MIC, mg/mL) | ||
|---|---|---|---|
| H. perforatum cv. Elixir | H. perforatum cv. Helos | H. perforatum cv. Topas | |
| Staphylococcus aureus | 15.0 | 7.5 | 10.0 |
| Staphylococcus epidermidis | 15.0 | 2.5 | 5.0 |
| Enterococcus faecalis | 15.0 | 10.0 | 10.0 |
| Enterococcus faecium | 15.0 | 10.0 | 10.0 |
| Bacillus subtilis | 10.0 | 2.5 | 2.5 |
| Escherichia coli | 40.0 | 40.0 | 40.0 |
| Enterobacter aerogenes | 40.0 | 40.0 | 40.0 |
| Enterobacter cloacae | 30.0 | 10.0 | 20.0 |
| Klebsiella pneumoniae | 40.0 | 30.0 | 40.0 |
| Pseudomonas aeruginosa | 30.0 | 20.0 | 30.0 |
| Species of Microorganisms | Antifungal Activity (MIC, mg/mL) | ||
|---|---|---|---|
| H. perforatum cv. Elixir | H. perforatum cv. Helos | H. perforatum cv. Topas | |
| Candida albicans | 7.5 | 15.0 | 10.0 |
| Candida krusei | 7.5 | 30.0 | 20.0 |
| Candida quilliermondii | 30.0 | 30.0 | 30.0 |
| Aspergillus flavus | 30.0 | 40.0 | 40.0 |
| Penicillium chrysogenum | 15.0 | 30.0 | 30.0 |
| Trichophyton tonsurans | 7.5 | 2.5 | 2.5 |
| Metabolites [mg/100 g DW] 1 | Hypericum perforatum | ||||||
|---|---|---|---|---|---|---|---|
| cv. Elixir | cv. Helos | cv. Topas | |||||
| Day | Control | Phe | Control | Phe | Control | Phe | |
| Protocatechuic acid | 2 | 32.52 a | 26.55 a | 43.93 abcd | 38.79 abc | 41.51 abcd | 39.10 abc |
| 4 | 61.60 cde | 58.65 bcde | 65.03 de | 64.94 de | 49.12 abcde | 31.63 a | |
| 7 | 68.97 e | 71.41 e | 72.14 e | 64.22 de | 57.26 bcde | 35.82 ab | |
| Neochlorogenic acid | 2 | 118.75 a | 118.19 a | 137.25 abc | 115.53 abc | 138.67 a | 123.06 a |
| 4 | 130.16 abc | 125.81 ab | 159.96 bc | 119.56 a | 122.30 a | 122.78 a | |
| 7 | 108.04 a | 120.11 a | 162.35 c | 131.43 abc | 124.18 a | 119.51 ab | |
| 3,4-Dihydroxyphenylacetic acid | 2 | 21.94 a | 35.76 a | 29.54 a | 25.93 a | 29.57 a | 48.34 ab |
| 4 | 177.65 de | 126.02 c | 127.06 c | 128.05 c | 82.90 b | 158.94 cd | |
| 7 | 192.39 def | 264.95 g | 222.31 f | 170.06 de | 129.14 c | 201.26 ef | |
| Chlorogenic acid | 2 | 2.55 a | 6.99 defg | 2.97 abc | 5.56 cdef | 3.16 abc | 2.83 a |
| 4 | 4.08 abc | 4.89 abcd | 2.92 ab | 7.78 fg | 5.10 abcde | 4.58 abcd | |
| 7 | 5.50 bcdef | 15.56 h | 4.64 abcd | 7.68 efg | 3.88 abc | 8.33 g | |
| Cryptochlorogenic acid | 2 | 47.37 cdef | 20.10 ab | 45.18 cde | 33.37 bcd | 61.32 efgh | 53.16 defg |
| 4 | 69.90 ghij | 67.64 fghi | 78.75 hij | 56.69 efg | 72.29 ghij | 29.99 abc | |
| 7 | 71.50 ghij | 90.34 j | 83.02 ij | 44.75 cde | 65.95 efghi | 10.50 a | |
| p-Hydroxybenzoic acid | 2 | 0.00 a | 120.46 bc | 0.00 a | 106.15 b | 0.00 a | 131.15 bc |
| 4 | 0.00 a | 129.16 bc | 0.00 a | 141.78 cd | 0.00 a | 123.47 bc | |
| 7 | 0.00 a | 139.10 cd | 0.00 a | 164.98 d | 0.00 a | 162.49 d | |
| Vanillic acid | 2 | 0.01 a | 0.01 a | 0.19 bcd | 0.24 cde | 0.01 a | 0.34 ef |
| 4 | 0.01 a | 0.07 ab | 0.34 ef | 0.31 de | 0.29 de | 0.31 de | |
| 7 | 0.01 a | 0.12 abc | 0.82 gh | 0.44 f | 0.71 g | 0.85 h | |
| p-Coumaric acid | 2 | 0.00 a | 59.47 bcd | 0.00 a | 45.48 b | 0.00 a | 56.24 bc |
| 4 | 0.00 a | 67.12 bcd | 0.00 a | 71.29 cd | 0.00 a | 64.22 bcd | |
| 7 | 0.00 a | 69.50 cd | 0.00 a | 78.15 d | 0.00 a | 74.36 cd | |
| Total phenolic acids | 2 | 223.14 a | 387.53 bc | 259.06 ab | 371.05 bc | 274.24 ab | 454.22 cde |
| 4 | 443.39 cd | 579.35 def | 434.04 cd | 590.40 ef | 332.00 abc | 535.91 def | |
| 7 | 446.40 cde | 771.09 g | 545.27 def | 661.69 fg | 381.12 bc | 613.12 f | |
| Hyperoside | 2 | 3.84 a | 145.29 cd | 9.79 a | 110.52 b | 8.85 a | 94.84 b |
| 4 | 6.56 a | 117.42 b | 6.37 a | 119.94 bc | 6.82 a | 113.56 b | |
| 7 | 11.61 a | 220.39 e | 9.71 a | 158.03 d | 7.89 a | 155.96 d | |
| Rutoside | 2 | 41.80 efg | 53.63 fgh | 45.01 efgh | 18.45 abc | 54.99 fgh | 13.09 ab |
| 4 | 65.17 hi | 39.61 def | 59.18 fgh | 31.66 cde | 53.14 fgh | 62.29 ghi | |
| 7 | 97.56 j | 91.48 j | 6.60 a | 131.31 k | 56.22 fgh | 83.25 ij | |
| Apigetrin | 2 | 0.00 a | 59.76 bc | 0.00 a | 45.67 b | 0.00 a | 42.54 b |
| 4 | 0.00 a | 75.67 cd | 0.00 a | 83.46 de | 0.00 a | 75.36 cd | |
| 7 | 0.00 a | 97.34 ef | 0.00 a | 115.64 fg | 0.00 a | 120.47 g | |
| Quercitrin | 2 | 62.57 c | 56.74 bc | 66.51 c | 28.32 ab | 57.02 bc | 17.30 a |
| 4 | 69.91 c | 166.08 fg | 54.71 bc | 151.34 ef | 55.07 bc | 135.10 e | |
| 7 | 102.60 d | 196.36 h | 9.27 a | 184.63 gh | 60.18 c | 126.36 de | |
| Quercetin | 2 | 1396.34 abcd | 1237.51 ab | 1606.27 cde | 1582.11 cde | 1175.05 a | 1776.04 e |
| 4 | 1729.13 e | 1700.44 de | 1541.45 bcde | 1522.16 bcde | 1184.85 a | 1726.70 e | |
| 7 | 2233.06 f | 2431.22 f | 2335.43 f | 2258.23 f | 1335.28 abc | 2796.61 g | |
| Luteolin | 2 | 9.89 ab | 69.33 g | 4.47 a | 29.45 bcde | 12.63 abc | 58.70 g |
| 4 | 15.14 abcd | 37.54 ef | 10.58 ab | 33.35 cdef | 17.86 abcde | 33.79 def | |
| 7 | 13.33 abcd | 27.72 bcde | 15.03 abcd | 33.21 cdef | 17.53 abcde | 51.35 fg | |
| Kaempferol | 2 | 7.30 a | 14.09 abcd | 8.16 a | 13.88 abcd | 9.10 a | 10.60 ab |
| 4 | 10.24 a | 19.98 cd | 9.60 ab | 19.26 bcd | 9.09 a | 11.47 abc | |
| 7 | 9.68 a | 21.54 d | 9.63 a | 5.57 a | 22.66 d | 10.72 abc | |
| Total flavonoids | 2 | 1521.74 ab | 1636.35 abc | 1740.21 abcd | 1828.41 bcd | 1317.65 a | 2013.11 cde |
| 4 | 1896.15 bcd | 2156.73 def | 1681.89 abc | 1961.16 cde | 1326.83 a | 2158.28 def | |
| 7 | 2467.83 fg | 3086.05 hi | 2385.67 ef | 2886.63 gh | 1499.76 ab | 3344.73 i | |
| Epigallocatechin | 2 | 0.00 a | 20.54 bc | 0.00 a | 29.54 d | 0.00 a | 15.15 b |
| 4 | 0.00 a | 29.15 cd | 0.00 a | 31.54 d | 0.00 a | 16.45 b | |
| 7 | 0.00 a | 36.05 d | 0.00 a | 33.48 d | 0.00 a | 20.47 b | |
| Catechin | 2 | 220.80 e | 112.03 bc | 176.16 d | 138.98 c | 224.56 ef | 86.71 b |
| 4 | 227.90 ef | 175.12 d | 258.36 f | 139.39 c | 232.45 ef | 124.30 c | |
| 7 | 214.26 e | 120.54 bc | 311.68 g | 21.10 a | 236.07 ef | 15.59 a | |
| Epicatechin | 2 | 182.97 cd | 361.71 g | 149.76 bc | 324.94 g | 218.56 def | 141.44 ab |
| 4 | 223.19 def | 460.78 i | 217.64 def | 428.10 hi | 209.20 de | 253.56 f | |
| 7 | 253.97 f | 407.47 h | 195.33 d | 450.95 i | 244.60 ef | 101.25 a | |
| Epicatechin gallate | 2 | 0.00 a | 45.04 de | 0.00 a | 58.14 fg | 0.00 a | 23.16 b |
| 4 | 0.00 a | 51.54 ef | 0.00 a | 60.50 fg | 0.00 a | 29.46 bc | |
| 7 | 0.00 a | 58.25 fg | 0.00 a | 67.24 g | 0.00 a | 35.41 cd | |
| Total catechins | 2 | 403.76 cd | 539.32 fghi | 325.92 bc | 551.60 ghi | 443.12 de | 266.46 b |
| 4 | 451.09 de | 716.58 k | 475.99 defg | 659.53 jk | 441.65 de | 423.77 d | |
| 7 | 468.23 def | 622.31 ij | 507.01 efgh | 572.77 hi | 480.67 defg | 172.72 a | |
| Groups of Microorganisms | Name and Number of Strains |
|---|---|
| Gram-positive bacteria | 1. Staphylococcus aureus ATCC 6538 P |
| 2. Staphylococcus epidermidis S3 | |
| 3. Enterococcus faecalis ATCC 8040/1 | |
| 4. Enterococcus faecium 34B/8 | |
| 5. Bacillus subtilis ATCC 6633 | |
| Gram-negative bacteria | 6. Escherichia coli ATCC 8739 |
| 7. Enterobacter aerogenes 35B | |
| 8. Enterobacter cloacae 382/2 | |
| 9. Klebsiella pneumoniae ATCC 16903 | |
| 10. Pseudomonas aeruginosa ATCC 27853 | |
| Yeast-like fungi | 11. Candida albicans PCM 1409 PZH |
| 12. Candida krusei S220 | |
| 13. Candida quilliermondii 11 | |
| Molds | 14. Aspergillus flavus 35/1 |
| 15. Penicillium chrysogenum ATCC 10106 | |
| Dermatophytes | 16. Trichophyton tonsurans 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecień, I.; Miceli, N.; Kędzia, E.; Cavò, E.; Taviano, M.F.; Beerhues, L.; Ekiert, H. Different Types of Hypericum perforatum cvs. (Elixir, Helos, Topas) In Vitro Cultures: A Rich Source of Bioactive Metabolites and Biological Activities of Biomass Extracts. Molecules 2023, 28, 2376. https://doi.org/10.3390/molecules28052376
Kwiecień I, Miceli N, Kędzia E, Cavò E, Taviano MF, Beerhues L, Ekiert H. Different Types of Hypericum perforatum cvs. (Elixir, Helos, Topas) In Vitro Cultures: A Rich Source of Bioactive Metabolites and Biological Activities of Biomass Extracts. Molecules. 2023; 28(5):2376. https://doi.org/10.3390/molecules28052376
Chicago/Turabian StyleKwiecień, Inga, Natalizia Miceli, Elżbieta Kędzia, Emilia Cavò, Maria Fernanda Taviano, Ludger Beerhues, and Halina Ekiert. 2023. "Different Types of Hypericum perforatum cvs. (Elixir, Helos, Topas) In Vitro Cultures: A Rich Source of Bioactive Metabolites and Biological Activities of Biomass Extracts" Molecules 28, no. 5: 2376. https://doi.org/10.3390/molecules28052376
APA StyleKwiecień, I., Miceli, N., Kędzia, E., Cavò, E., Taviano, M. F., Beerhues, L., & Ekiert, H. (2023). Different Types of Hypericum perforatum cvs. (Elixir, Helos, Topas) In Vitro Cultures: A Rich Source of Bioactive Metabolites and Biological Activities of Biomass Extracts. Molecules, 28(5), 2376. https://doi.org/10.3390/molecules28052376