
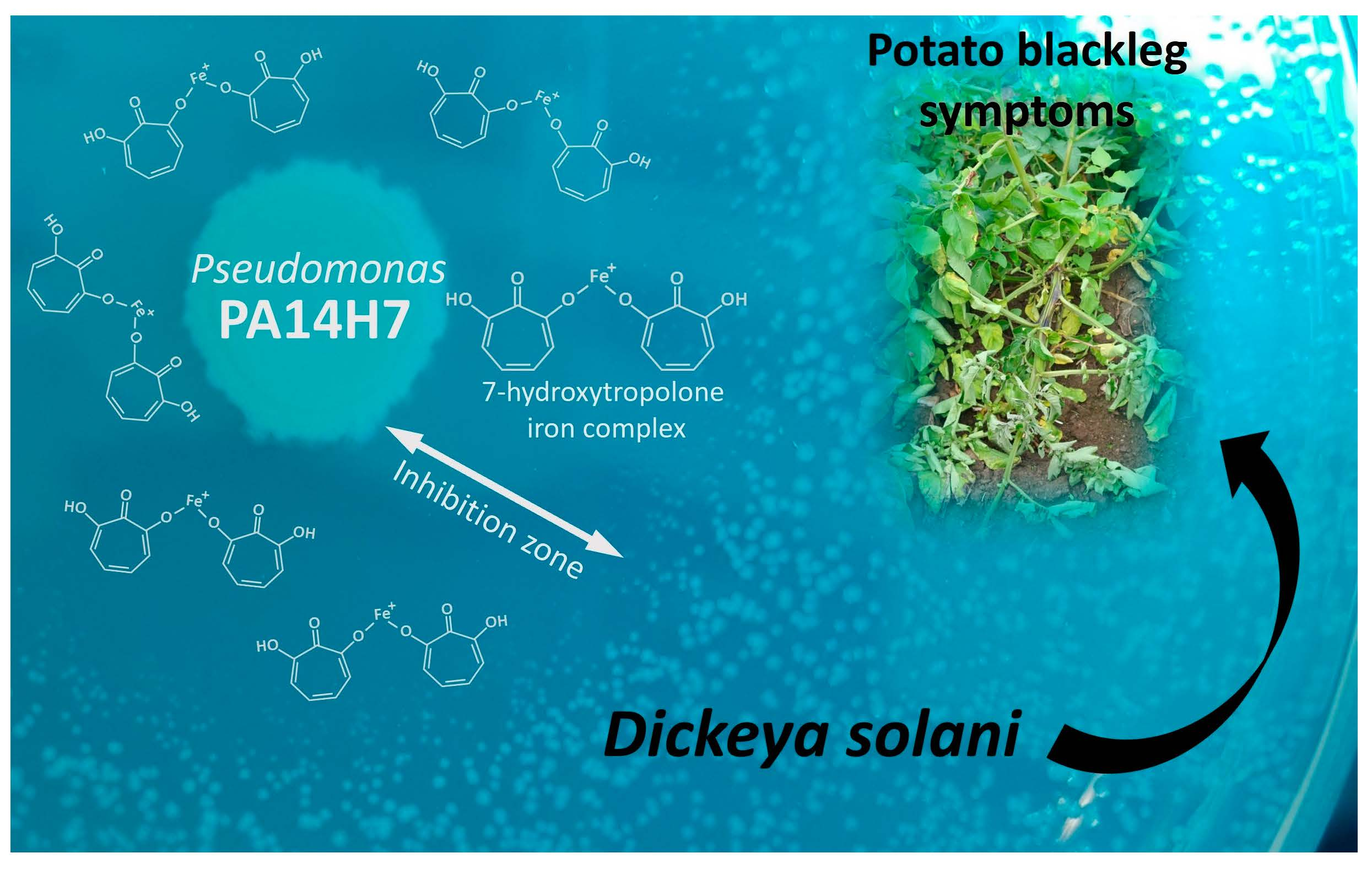
Pseudomonas PA14H7: Identification and Quantification of the 7-Hydroxytropolone Iron Complex as an Active Metabolite against Dickeya, the Causal Agent of Blackleg on the Potato Plant

 , ,
, ,  , , and
, , and
Abstract
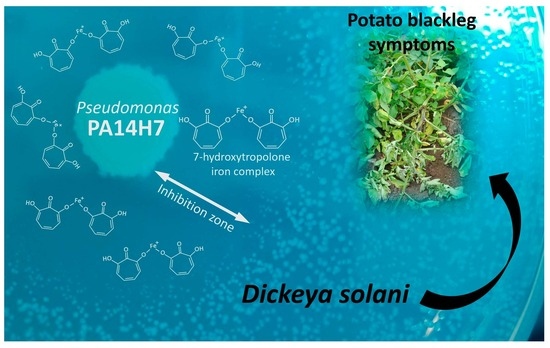
:
1. Introduction
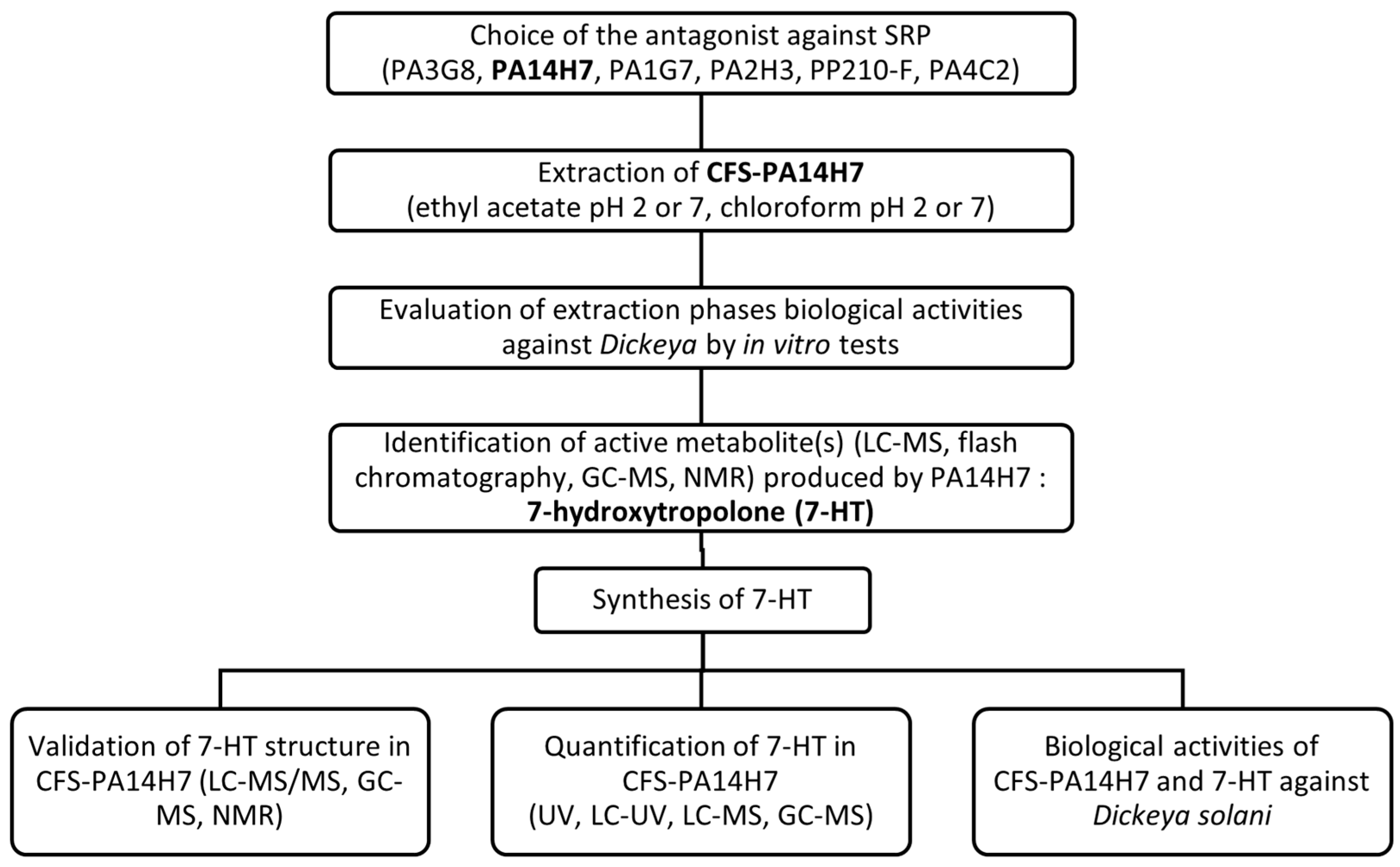
2. Results and Discussion
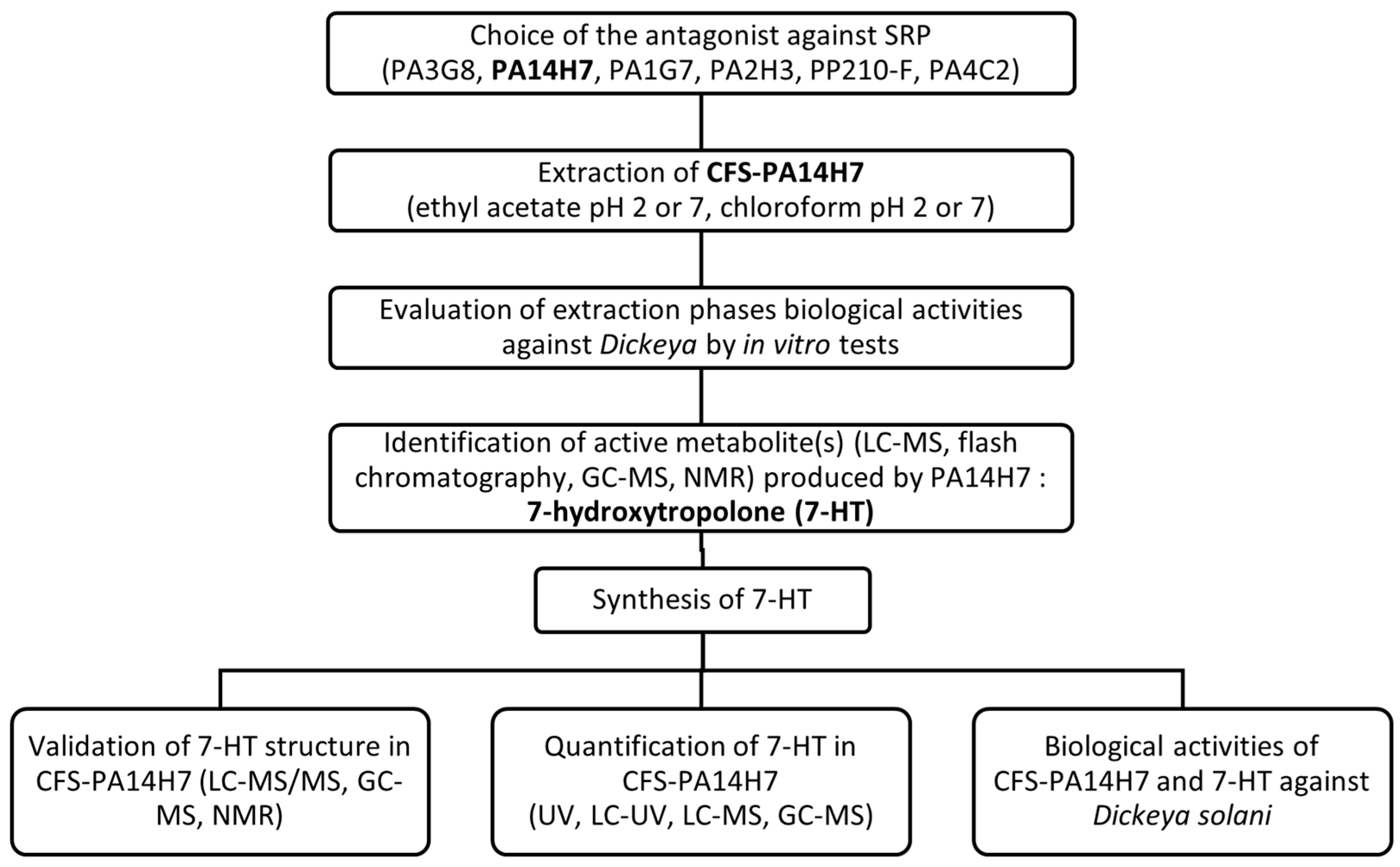
2.1. Choice of the Main Active Antagonist
2.2. Evaluation of CFS-PA14H7 Extraction Methods
2.3. Identification of the Active Metabolite(s)
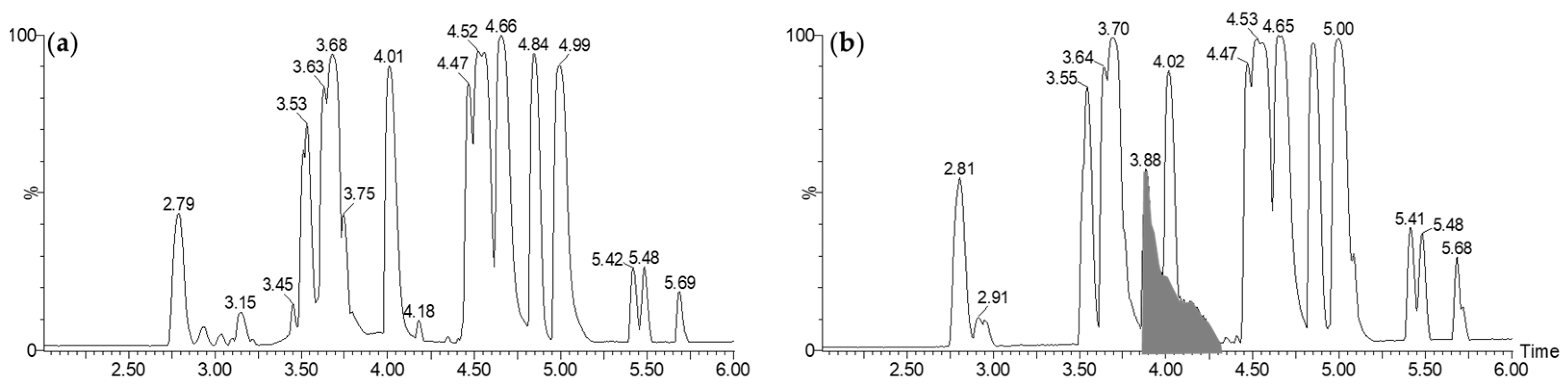
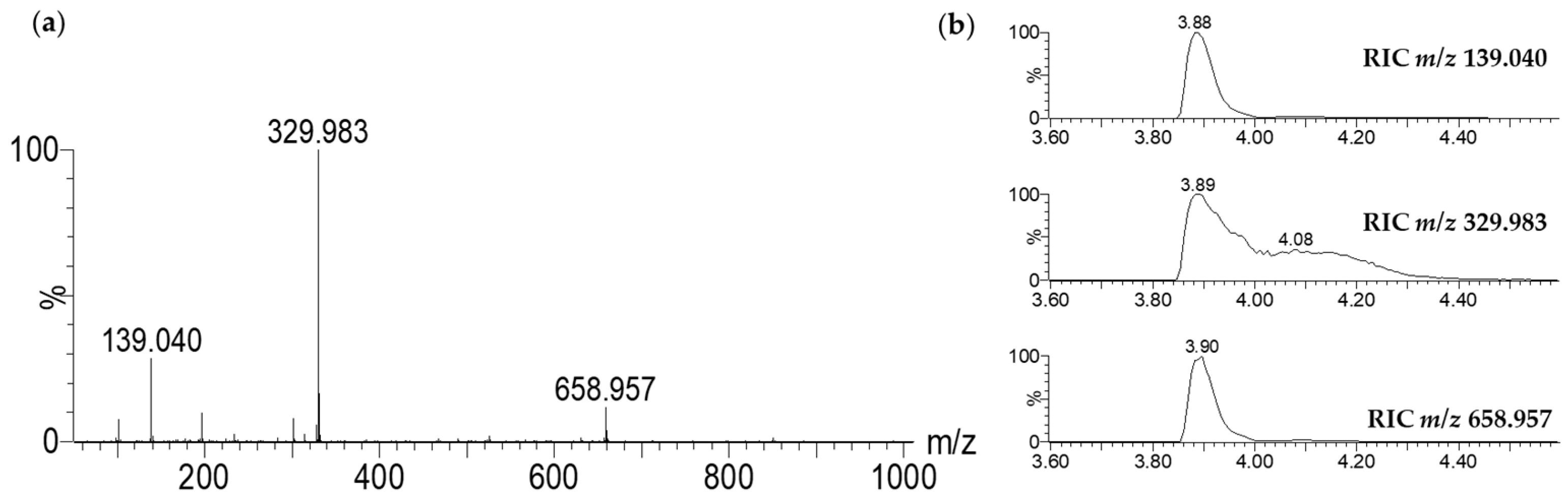
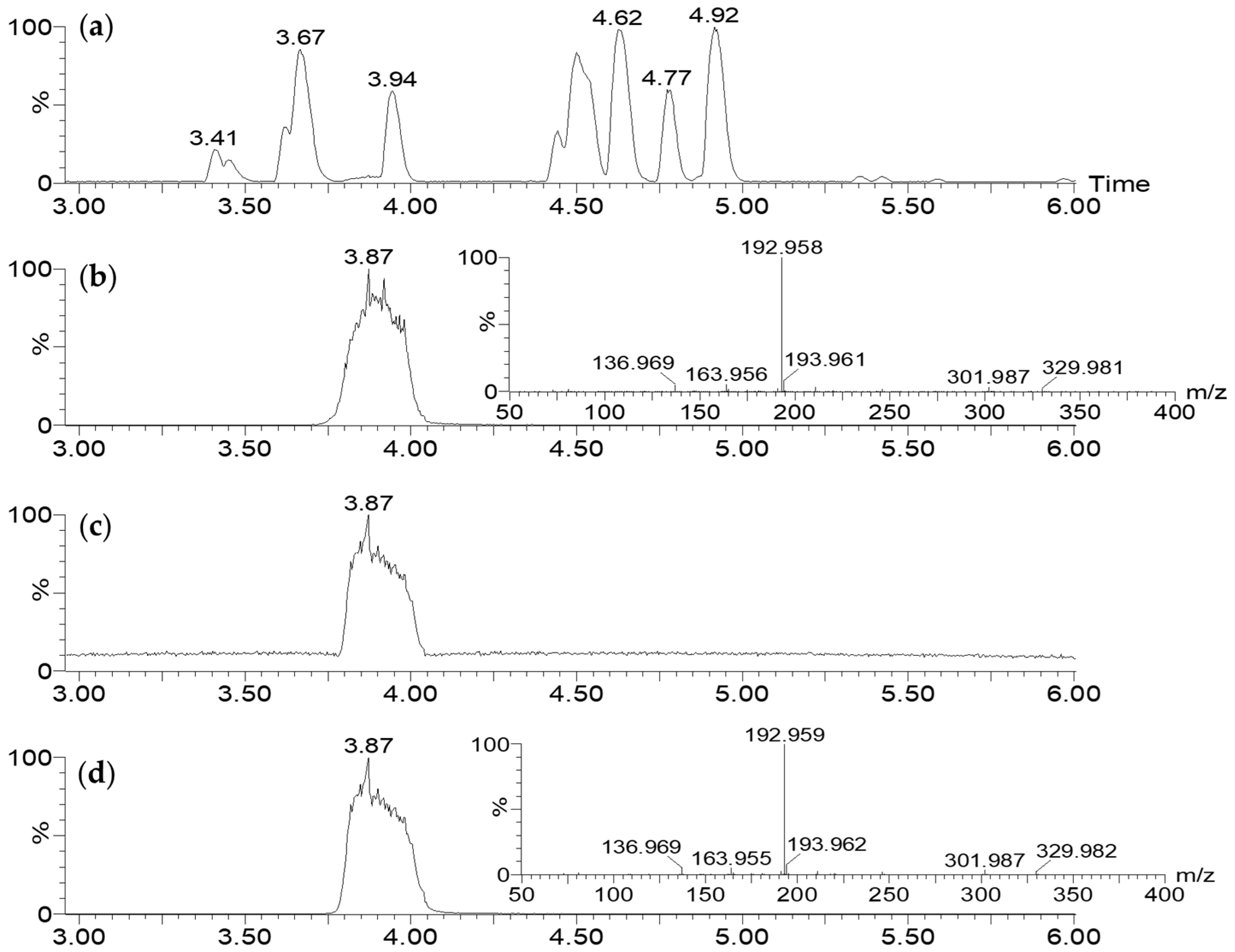
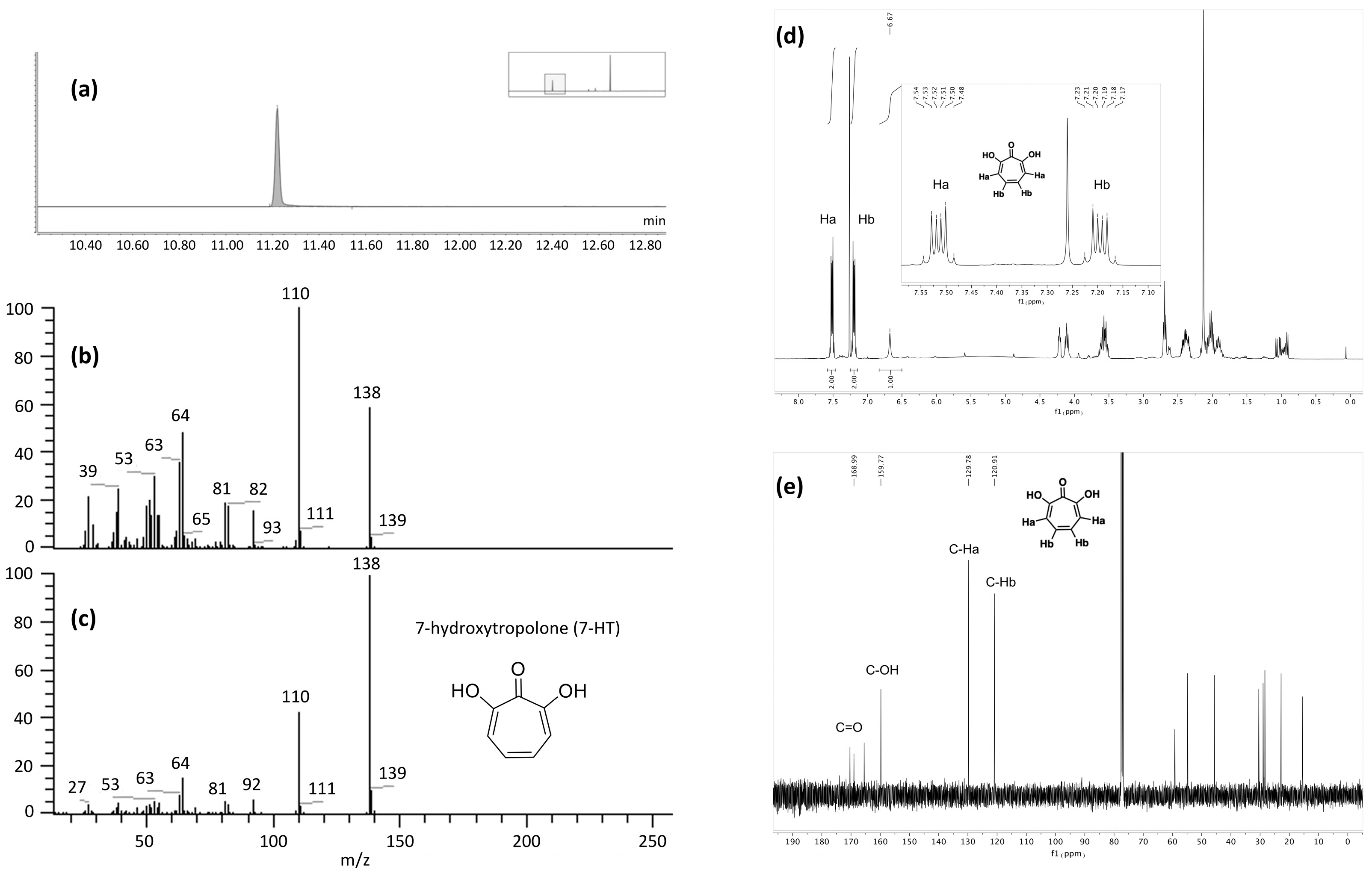
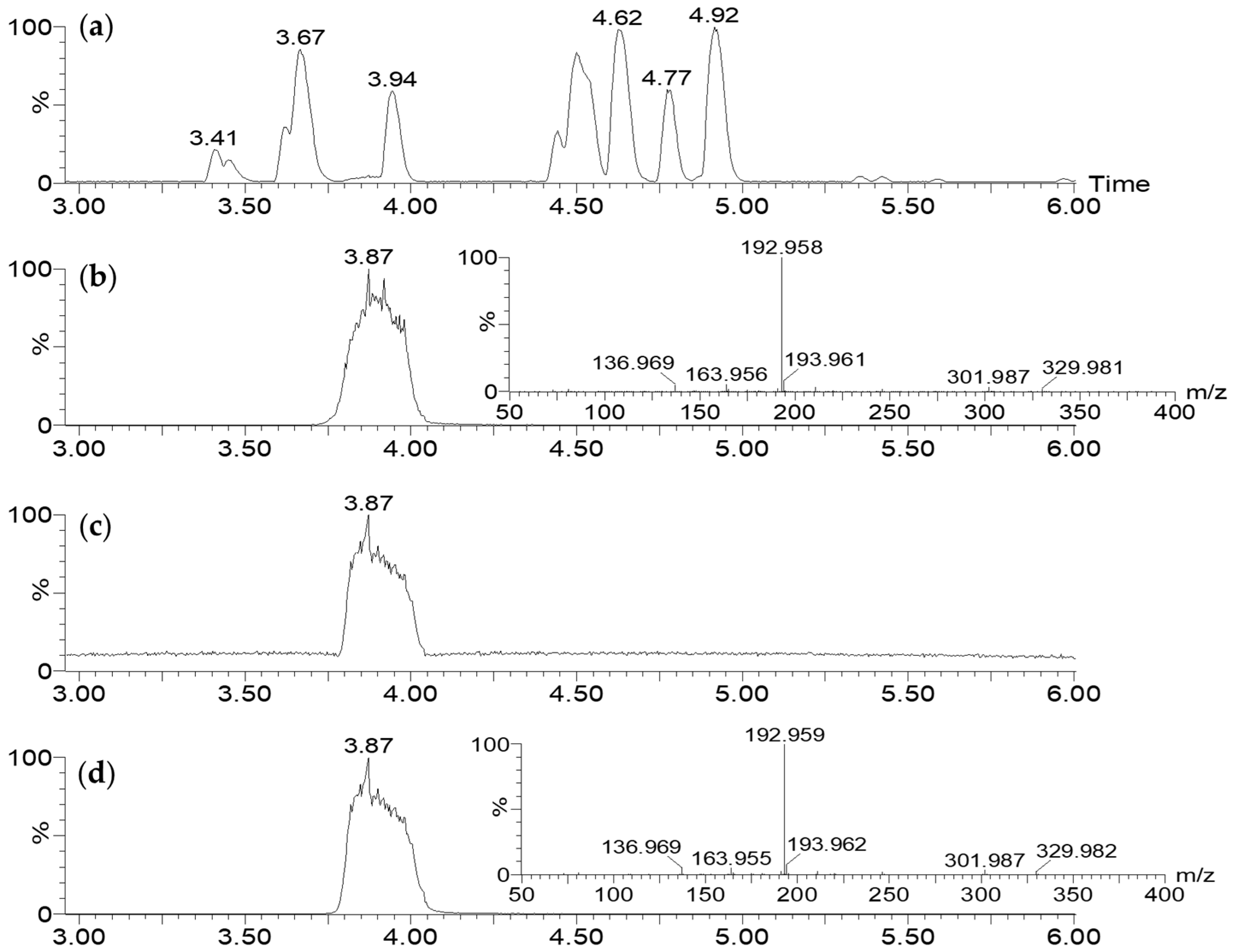
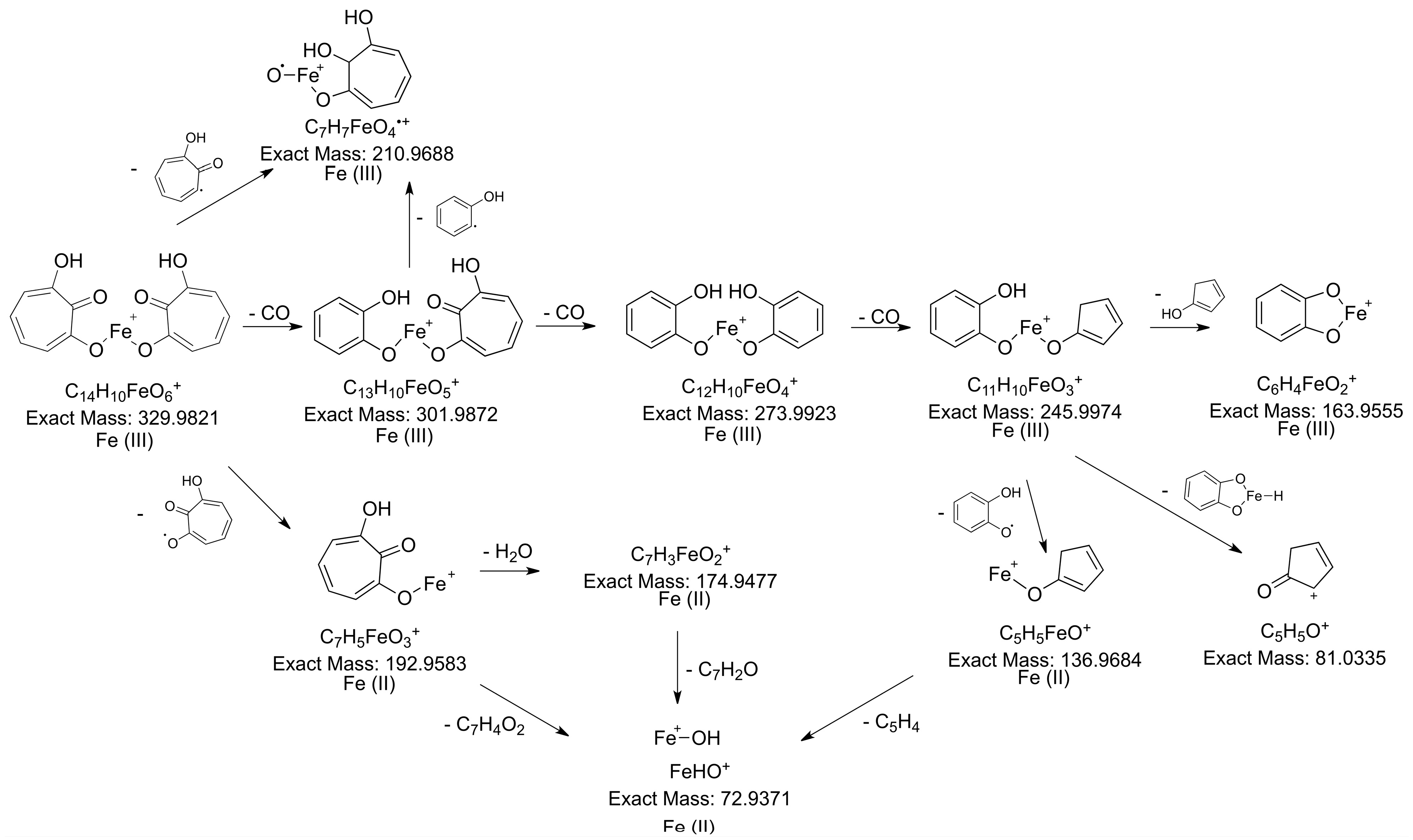
2.3.1. LC-MS Analysis
2.3.2. Biologically Driven Purification Using Flash Chromatography
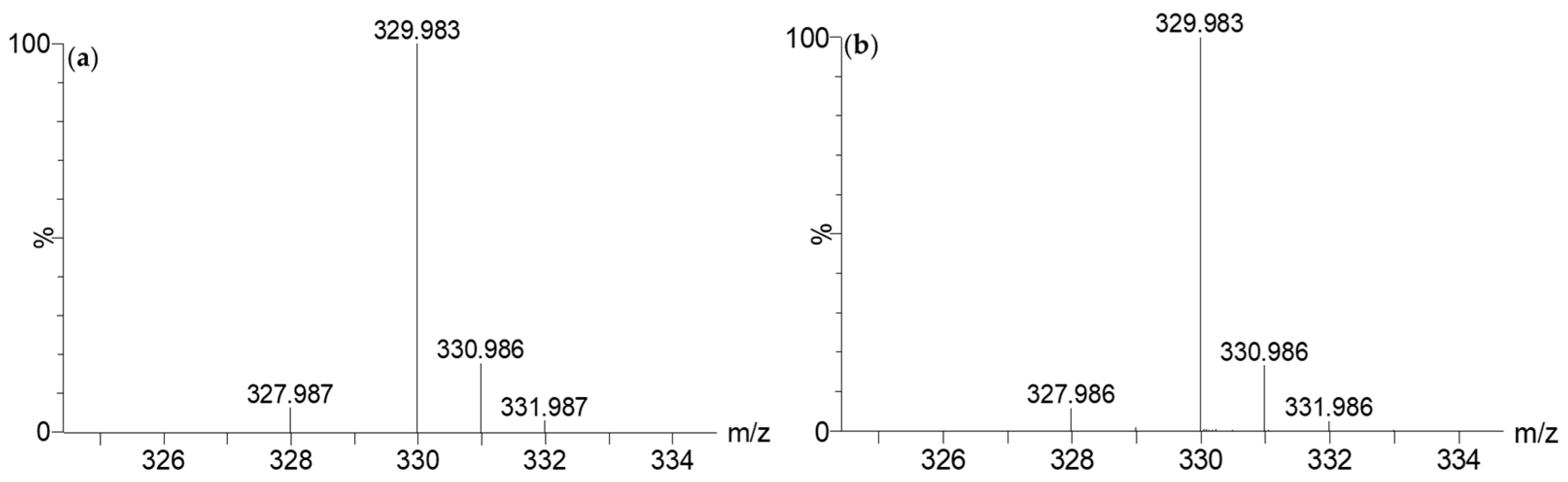
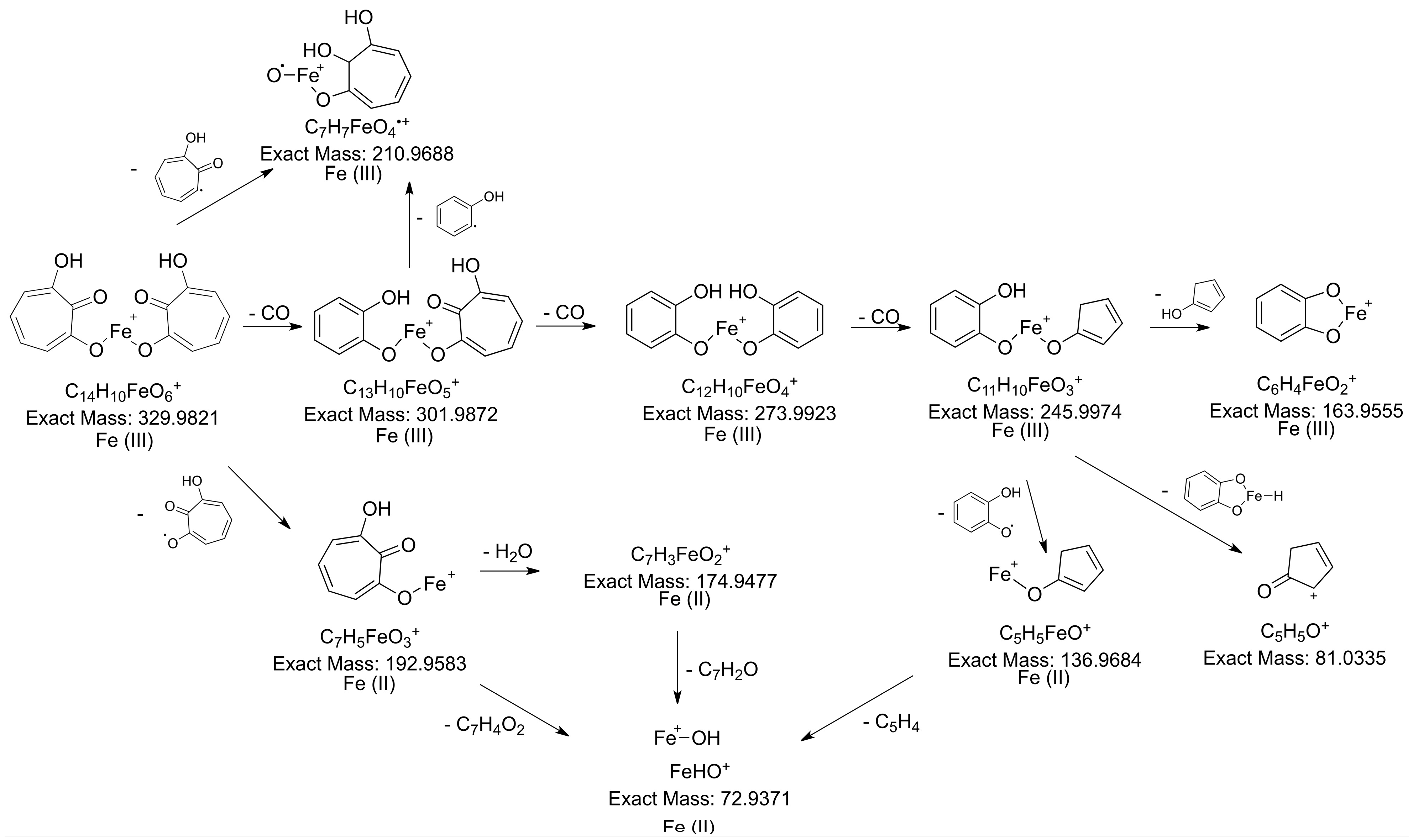
2.3.3. Determination of the Metabolite Structure
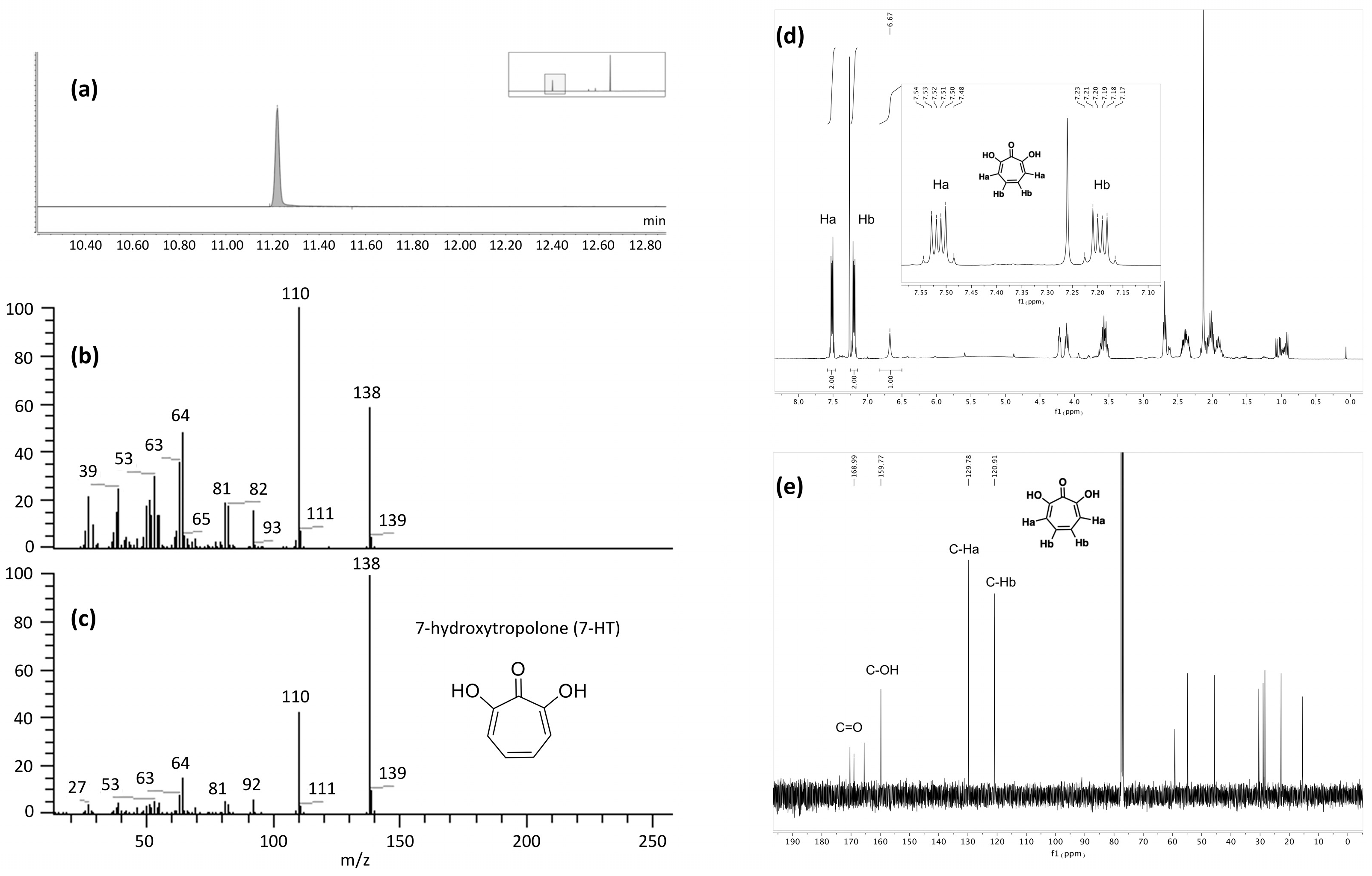
2.4. Validation of 7-HT Structure and Activity
2.4.1. Organic Synthesis of 7-HT
2.4.2. Quantification of 7-HT Produced by PA14H7
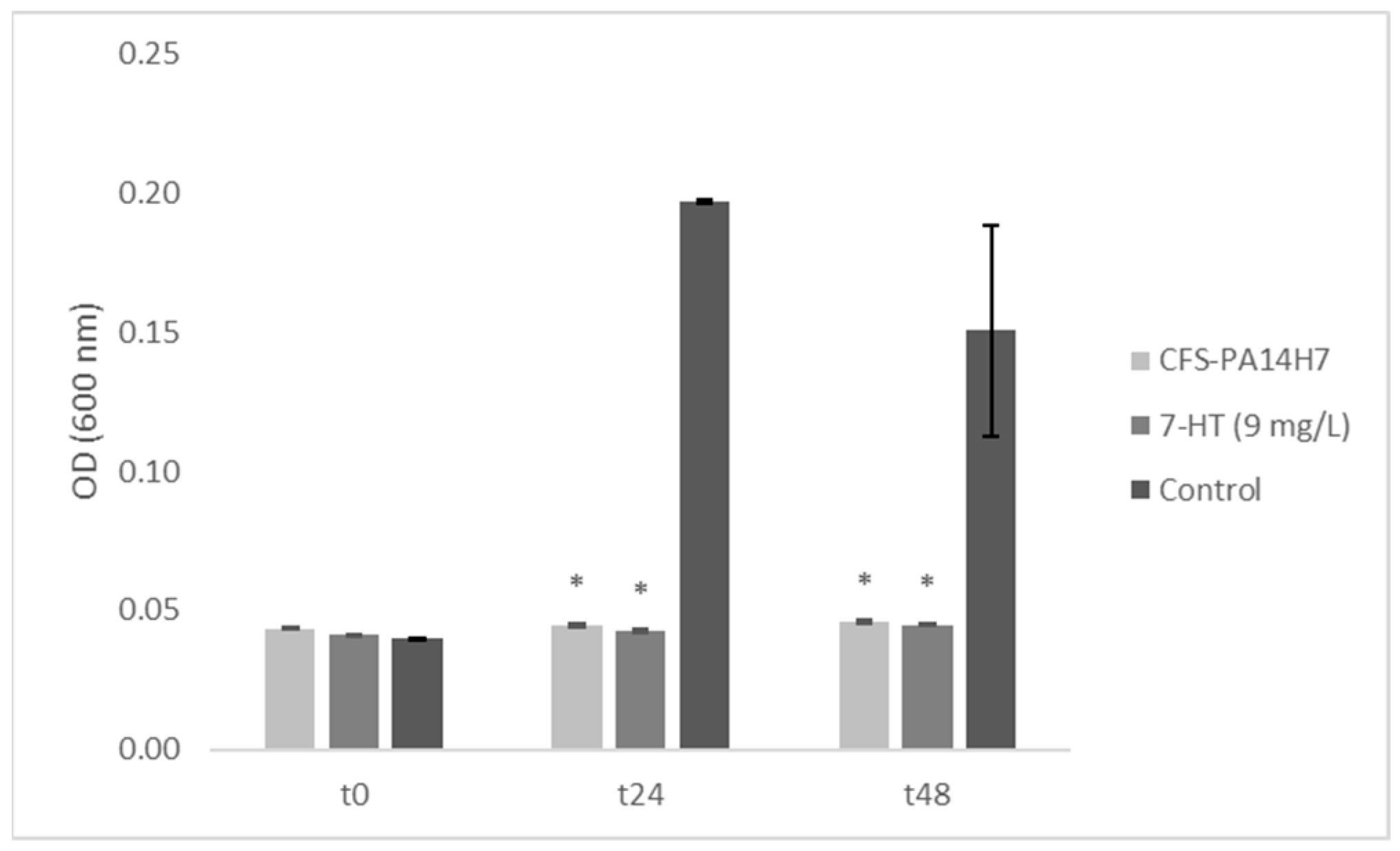
2.4.3. In Vitro Test of CFS-PA14H7 vs. Synthetic 7-HT
3. Materials and Methods
3.1. Materials
3.2. Biological Material
3.3. Biological Test
3.4. Kinetics Growth
3.5. Extraction Method of CFS-PA14H7
3.6. LC-UV-MS
3.7. Purification by Flash Chromatography
3.8. GC-MS
3.9. NMR
3.10. Quantification of 7-HT
3.11. Preliminary In Vivo Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Pérombelon, M.C.M. Potato Blackleg: Epidemiology, Host-Pathogen Interaction and Control. Neth. J. Plant Pathol. 1992, 98, 135–146. [Google Scholar] [CrossRef]
- Toth, I.K.; Barny, M.; Czajkowski, R.; Elphinstone, J.G.; Li, X.; Pédron, J.; Pirhonen, M.; Van Gijsegem, F. Pectobacterium and Dickeya: Taxonomy and Evolution. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 13–37. ISBN 978-3-030-61459-1. [Google Scholar]
- Van Der Wolf, J.M.; Acuña, I.; De Boer, S.H.; Brurberg, M.B.; Cahill, G.; Charkowski, A.O.; Coutinho, T.; Davey, T.; Dees, M.W.; Degefu, Y.; et al. Diseases Caused by Pectobacterium and Dickeya Species around the World. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., Van Der Wolf, J.M., Toth, I.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 215–261. ISBN 978-3-030-61458-4. [Google Scholar]
- Règlements Techniques de la Production, du Contrôle et de la Certification des Semences et Plants—SEMAE. Available online: https://www.semae.fr/service-officiel-controle-et-certification/reglements-techniques-production-controle-et-certification/ (accessed on 2 May 2023).
- UNECE. UNECE STANDARD D-1 Concerning the Marketing and Commercial Quality Control of Seed Potato; United Nations: New York, NY, USA; Geneva, Switzerland, 2018. [Google Scholar]
- Mukhametov, A.; Shamekova, M.; Dautkanova, D.; Kazhymurat, A.; Ilyassova, G. Seed Potato Production Regulatory Framework Established in Top Potato Producing Countries: Comparison of the GOST (Russia) and UNECE S-1 Certification Systems|Elsevier Enhanced Reader. J. Agric. Food Res. 2023, 11, 100520. [Google Scholar] [CrossRef]
- Zeng, Y.; Fulladolsa, A.C.; Houser, A.; Charkowski, A.O. Colorado Seed Potato Certification Data Analysis Shows Mosaic and Blackleg Are Major Diseases of Seed Potato and Identifies Tolerant Potato Varieties. Plant Dis. 2019, 103, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, B.; Nkuriyingoma, P.; Van Gijsegem, F. Economic Impact of Pectobacterium and Dickeya Species on Potato Crops: A Review and Case Study. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 263–282. ISBN 978-3-030-61459-1. [Google Scholar]
- Cirou, A. Développement de Stratégies d’anti-Virulence Ciblant La Régulation Quorum-Sensing Chez Pectobacterium atrosepticum, Bactérie Pathogène de La Pomme de Terre. Ph.D. Thesis, Université Paris-Saclay, Bures-sur-Yvette, France, 2010. [Google Scholar]
- Cirou, A.; Mondy, S.; An, S.; Charrier, A.; Sarrazin, A.; Thoison, O.; DuBow, M.; Faure, D. Efficient Biostimulation of Native and Introduced Quorum-Quenching Rhodococcus Erythropolis Populations Is Revealed by a Combination of Analytical Chemistry, Microbiology, and Pyrosequencing. Appl. Environ. Microbiol. 2012, 78, 481–492. [Google Scholar] [CrossRef]
- Blin, P.; Robic, K.; Khayi, S.; Cigna, J.; Munier, E.; Dewaegeneire, P.; Laurent, A.; Jaszczyszyn, Y.; Hong, K.; Chan, K.; et al. Pattern and Causes of the Establishment of the Invasive Bacterial Potato Pathogen Dickeya Solani and of the Maintenance of the Resident Pathogen D. dianthicola. Mol. Ecol. 2021, 30, 608–624. [Google Scholar] [CrossRef]
- Zomorodian, K.; Ghadiri, P.; Saharkhiz, M.J.; Moein, M.R.; Mehriar, P.; Bahrani, F.; Golzar, T.; Pakshir, K.; Fani, M.M. Antimicrobial Activity of Seven Essential Oils from Iranian Aromatic Plants against Common Causes of Oral Infections. Jundishapur J. Microbiol. 2015, 8, e17766. [Google Scholar] [CrossRef]
- Hajian-Maleki, H.; Baghaee-Ravari, S.; Moghaddam, M. Efficiency of Essential Oils against Pectobacterium carotovorum Subsp. Carotovorum Causing Potato Soft Rot and Their Possible Application as Coatings in Storage. Postharvest Biol. Technol. 2019, 156, 110928. [Google Scholar] [CrossRef]
- Azaiez, S.; Ben Slimene, I.; Karkouch, I.; Essid, R.; Jallouli, S.; Djebali, N.; Elkahoui, S.; Limam, F.; Tabbene, O. Biological Control of the Soft Rot Bacterium Pectobacterium carotovorum by Bacillus amyloliquefaciens Strain Ar10 Producing Glycolipid-like Compounds. Microbiol. Res. 2018, 217, 23–33. [Google Scholar] [CrossRef]
- Toth, I.K.; van der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror Lahkim, L.; Elphinstone, J.G. Dickeya Species: An Emerging Problem for Potato Production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- de Weert, S.; Bloemberg, G.V. Rhizosphere Competence and the Role of Root Colonization in Biocontrol. In Plant-Associated Bacteria; Gnanamanickam, S.S., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 317–333. ISBN 978-1-4020-4538-7. [Google Scholar]
- Raoul des Essarts, Y. Pathogénie de Dickeya dianthicola et Dickeya solani chez Solanum tuberosum, Développement et Evaluation de Stratégies de Lutte Biologique. Ph.D. Thesis, Université Paris Sud, Bures-sur-Yvette, France, 2015. [Google Scholar]
- Raoul des Essarts, Y.; Cigna, J.; Quêtu-Laurent, A.; Caron, A.; Munier, E.; Beury-Cirou, A.; Hélias, V.; Faure, D. Biocontrol of the Potato Blackleg and Soft Rot Diseases Caused by Dickeya dianthicola. Appl. Environ. Microbiol. 2016, 82, 268–278. [Google Scholar] [CrossRef]
- Cigna, J.; Robic, K.; Dewaegeneire, P.; Hélias, V.; Beury, A.; Faure, D. Efficacy of Soft-Rot Disease Biocontrol Agents in the Inhibition of Production Field Pathogen Isolates. Microorganisms 2023, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Syed Ab Rahman, S.F.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging Microbial Biocontrol Strategies for Plant Pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Arora, N.K. Secondary Metabolites of Fluorescent Pseudomonads in Biocontrol of Phytopathogens for Sustainable Agriculture. Appl. Soil Ecol. 2018, 125, 35–45. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-Associated Bacillus and Pseudomonas Antimicrobial Activities in Plant Disease Suppression via Biological Control Mechanisms—A Review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Dowling, D.; O’Gara, F. Metabolites of Pseudomonas Involved in the Biocontrol of Plant Disease. Trends Biotechnol. 1994, 12, 133–141. [Google Scholar] [CrossRef]
- Muzio, F.M.; Agaras, B.C.; Masi, M.; Tuzi, A.; Evidente, A.; Valverde, C. 7-hydroxytropolone Is the Main Metabolite Responsible for the Fungal Antagonism of Pseudomonas donghuensis Strain SVBP6. Environ. Microbiol. 2020, 22, 2550–2563. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pagnani, G.; Bernardi, M.; Mattedi, A.; Spera, D.M.; Gallo, M.D. Cell-Free Supernatants of Plant Growth-Promoting Bacteria: A Review of Their Use as Biostimulant and Microbial Biocontrol Agents in Sustainable Agriculture. Sustainability 2020, 12, 9917. [Google Scholar] [CrossRef]
- Jain, R.; Pandey, A. A Phenazine-1-Carboxylic Acid Producing Polyextremophilic Pseudomonas Chlororaphis (MCC2693) Strain, Isolated from Mountain Ecosystem, Possesses Biocontrol and Plant Growth Promotion Abilities. Microbiol. Res. 2016, 190, 63–71. [Google Scholar] [CrossRef]
- Winter, N.; Trauner, D. Thiocarbonyl Ylide Chemistry Enables a Concise Synthesis of (±)-Hippolachnin A. J. Am. Chem. Soc. 2017, 139, 11706–11709. [Google Scholar] [CrossRef]
- Takeshita, H.; Mori, A. An Improved Synthesis of 2,7-Dihydroxytropolone (3-Hydroxytropolone). Synthesis 1986, 578–579. [Google Scholar] [CrossRef]
- Hamor, T.A.; Watkin, D.J. The Crystal Structure of Tris(Tropolonato)Iron (III) (“Ferric Tropolone”). J. Chem. Soc. D 1969, 9, 440–441. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, M.; Yu, X.; Xie, Z. 7-Hydroxytropolone Produced and Utilized as an Iron-Scavenger by Pseudomonas Donghuensis. Biometals 2016, 29, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Roman, D.; Beemelmanns, C. Tropolone Natural Products. Nat. Prod. Rep. 2019, 36, 1137–1155. [Google Scholar] [CrossRef] [PubMed]
- Cigna, J.; Raoul des Essarts, Y.; Mondy, S.; Hélias, V.; Beury-Cirou, A.; Faure, D. Draft Genome Sequences of Pseudomonas Fluorescens Strains PA4C2 and PA3G8 and Pseudomonas Putida PA14H7, Three Biocontrol Bacteria against Dickeya Phytopathogens. Genome Announc. 2015, 3, e01503-14. [Google Scholar] [CrossRef]
- Pédron, J.; Mondy, S.; Raoul des Essarts, Y.; Van Gijsegem, F.; Faure, D. Genomic and Metabolic Comparison with Dickeya dadantii 3937 Reveals the Emerging Dickeya solani Potato Pathogen to Display Distinctive Metabolic Activities and T5SS/T6SS-Related Toxin Repertoire. BMC Genom. 2014, 15, 283. [Google Scholar] [CrossRef]
- Cox, R.J.; Al-Fahad, A. Chemical Mechanisms Involved during the Biosynthesis of Tropolones. Curr. Opin. Chem. Biol. 2013, 17, 532–536. [Google Scholar] [CrossRef]
- Haluk, J.-P.; Roussel-Bousta, C. Biosynthèse de tropolones dans les cals et les suspensions cellulaires à partir d’ébauches foliaires de plantules de Thuja plicata Donn. Ann. For. Sci. 2003, 60, 271–276. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.C.M.; van Veen, J.A.; van der Wolf, J.M. Control of Blackleg and Tuber Soft Rot of Potato Caused by Pectobacterium and Dickeya Species: A Review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CFS of Antagonistic Bacteria Strain | |||||||
|---|---|---|---|---|---|---|---|
| SRP Strain | Blank | PA3G8 | PA1G7 | PA4C2 | PA14H7 | PA2H3 | PP1-210F |
| P. atrosepticum P8-Me25a | − | − | ++ | − | ++ | − | ++ |
| P. parmentieri P13-CH22 | − | − | − | − | + | − | − |
| P. brasiliense P1-15C1 | − | − | − | − | + | − | − |
| P. polaris P1-10C1 | − | − | − | − | + | − | − |
| D. dianthicola P15-29 | − | − | − | − | + | − | − |
| D. solani P5-Sp1a | − | − | − | − | + | − | − |
| D. solani 3337 | − | − | − | − | + | − | − |
| Aqueous Phase (Dilution Factor) | Organic Phase (Dilution Factor) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Extraction Condition | 1 | 0.5 | 0.25 | 0.1 | 0.05 | 0.01 | 1 | 0.5 | 0.25 | 0.1 | 0.05 | 0.01 | |
| Ethyl acetate | pH 2 | − | − | − | − | − | − | ++ | ++ | ++ | ++ | ++ | − |
| pH 7 | − | − | − | − | − | − | ++ | ++ | ++ | ++ | + | − | |
| Chloroform | pH 2 | − | − | − | − | − | − | ++ | ++ | ++ | ++ | ++ | − |
| pH 7 | − | − | − | − | − | − | ++ | ++ | + | + | + | − | |
| Dilution Factor | ||||||||
|---|---|---|---|---|---|---|---|---|
| Fraction | 1 | 0.5 | 0.25 | 0.1 | 0.05 | 0.01 | 0.005 | 0.001 |
| F1 | − | − | − | − | − | − | − | − |
| F2 | ++ | ++ | ++ | ++ | ++ | + | + | − |
| F3 | ++ | ++ | + | − | − | − | − | − |
| Analytical Method | |||||
|---|---|---|---|---|---|
| Extraction Condition | LC-UV | LC-MS | GC-MS | ||
| Extraction of CFS PA14H7 1 | Ethyl acetate | pH 2 | 8.9 | 8.5 | 12.4 |
| pH 7 | 5.7 | 6.0 | 7.3 | ||
| Chloroform | pH 2 | 7.9 | 8.2 | 10.0 | |
| pH 7 | 10.3 | 8.5 | 8.9 | ||
| Synthetic 7-HT in water (9.6 mg/L) | Ethyl acetate | pH 2 | n.d. | 8.9 | 9.7 |
| Chloroform | pH 7 | n.d. | 11.3 | 9.2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munier-Lépinay, E.; Mathiron, D.; Quéro, A.; Khelifa, M.; Laclef, S.; Pilard, S. Pseudomonas PA14H7: Identification and Quantification of the 7-Hydroxytropolone Iron Complex as an Active Metabolite against Dickeya, the Causal Agent of Blackleg on the Potato Plant. Molecules 2023, 28, 6207. https://doi.org/10.3390/molecules28176207
Munier-Lépinay E, Mathiron D, Quéro A, Khelifa M, Laclef S, Pilard S. Pseudomonas PA14H7: Identification and Quantification of the 7-Hydroxytropolone Iron Complex as an Active Metabolite against Dickeya, the Causal Agent of Blackleg on the Potato Plant. Molecules. 2023; 28(17):6207. https://doi.org/10.3390/molecules28176207
Chicago/Turabian StyleMunier-Lépinay, Euphrasie, David Mathiron, Anthony Quéro, Mounia Khelifa, Sylvain Laclef, and Serge Pilard. 2023. "Pseudomonas PA14H7: Identification and Quantification of the 7-Hydroxytropolone Iron Complex as an Active Metabolite against Dickeya, the Causal Agent of Blackleg on the Potato Plant" Molecules 28, no. 17: 6207. https://doi.org/10.3390/molecules28176207
APA StyleMunier-Lépinay, E., Mathiron, D., Quéro, A., Khelifa, M., Laclef, S., & Pilard, S. (2023). Pseudomonas PA14H7: Identification and Quantification of the 7-Hydroxytropolone Iron Complex as an Active Metabolite against Dickeya, the Causal Agent of Blackleg on the Potato Plant. Molecules, 28(17), 6207. https://doi.org/10.3390/molecules28176207