

Protective Effects of Sophorae tonkinensis Gagnep. (Fabaceae) Radix et Rhizoma Water Extract on Carbon Tetrachloride-Induced Acute Liver Injury

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Experimental Animals
2.3. Preparation of the Test Drug
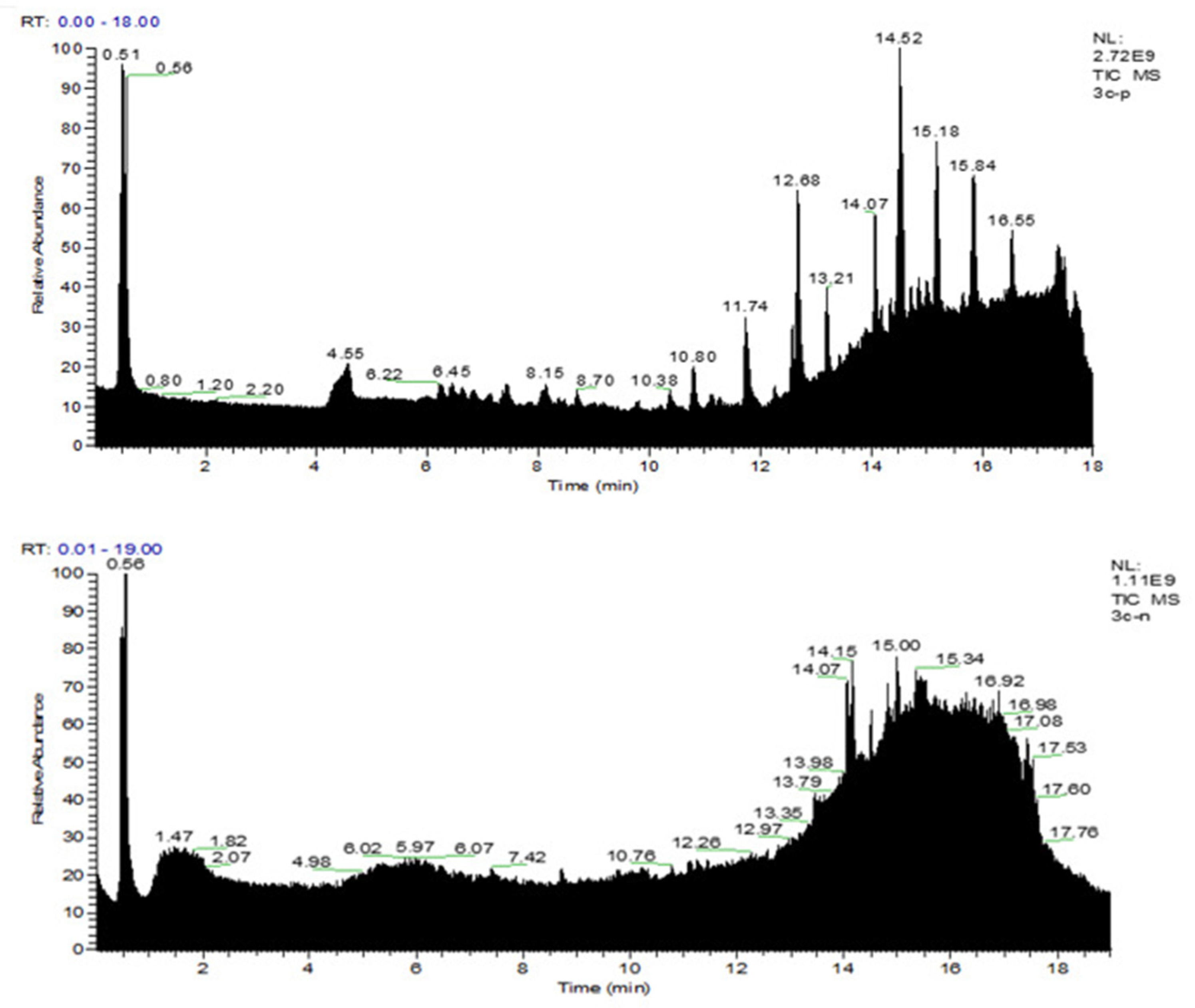
2.4. Ultra-High-Performance Liquid Chromatography-Tandem Mass Spectrometry (UPLC-MS/MS) Analysis of STR
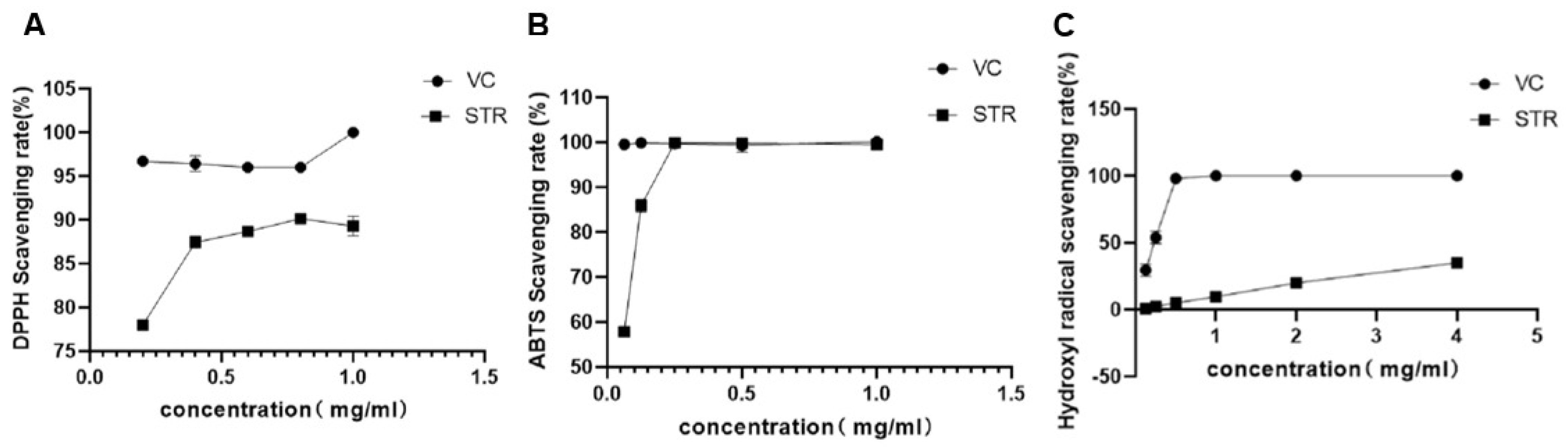
2.5. Analysis of STR Antioxidant Activity In Vitro
2.5.1. DPPH Radical Scavenging Test
2.5.2. ABTS Radical Scavenging Test
2.5.3. Hydroxyl Radical Scavenging Test
2.6. Animal Treatment
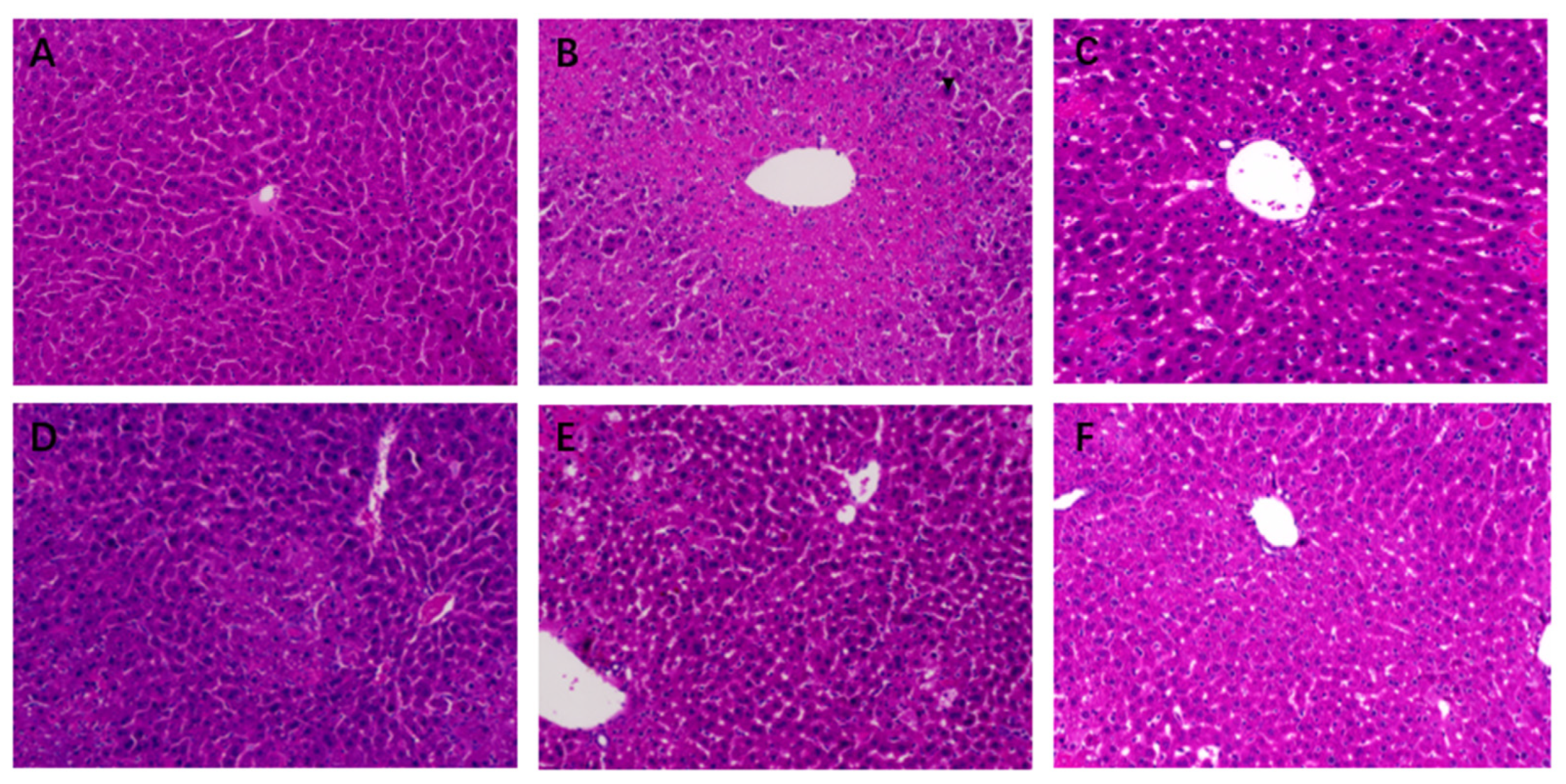
2.7. Histopathological Liver Examination
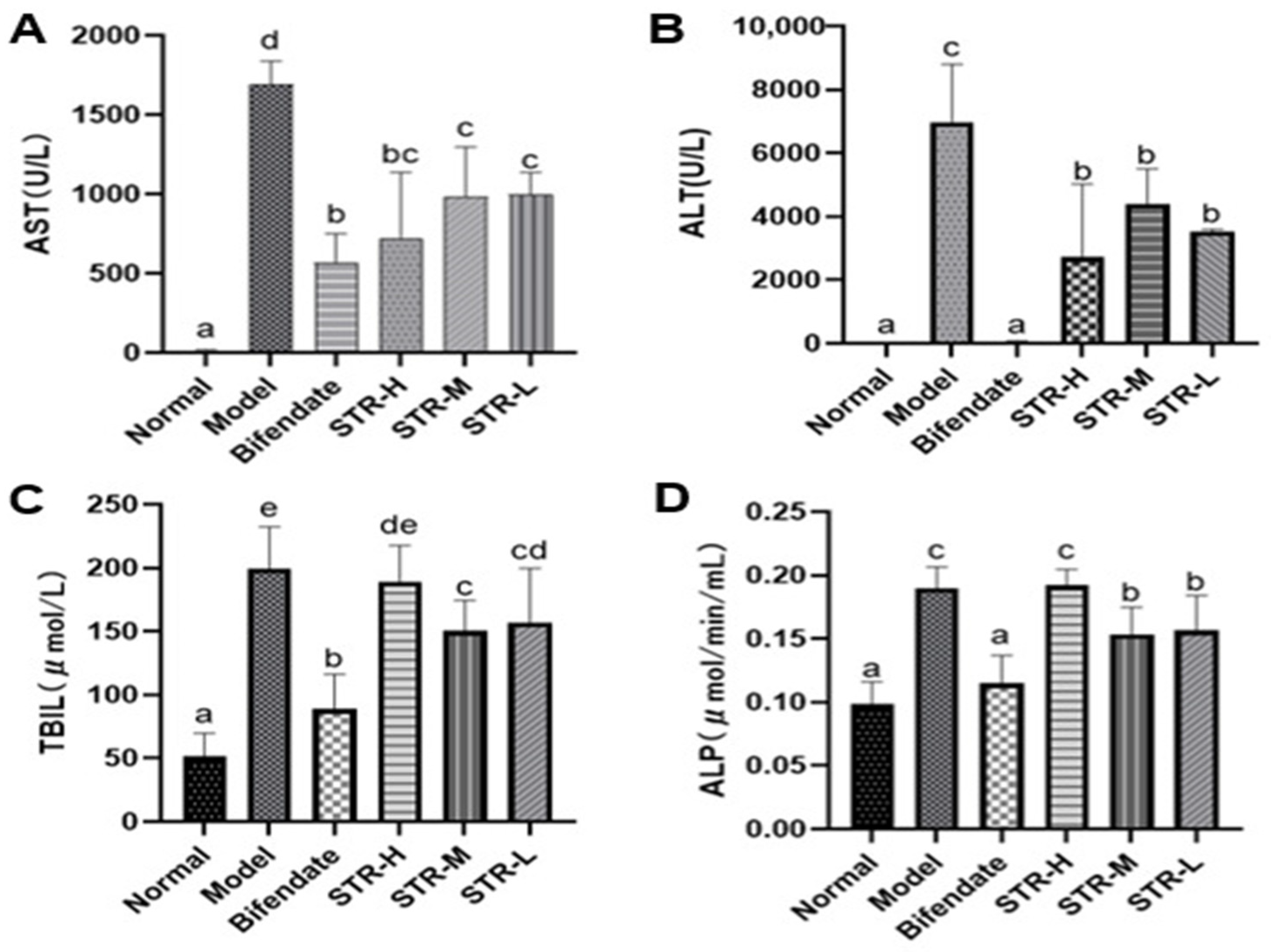
2.8. Serum Biochemistry
2.9. ELISA
2.10. Real-Time Polymerase Chain Reaction (qRT-PCR)
2.11. Hepatocyte Apoptosis
2.12. Statistical Analysis
3. Results
3.1. Quality Control Analysis of STR by UPLC-MS/MS
3.2. Antioxidant Assay of STR In Vitro
3.3. Effect of STR on Liver Histopathology in Mice
3.4. Effect of STR on Liver Function Indicators
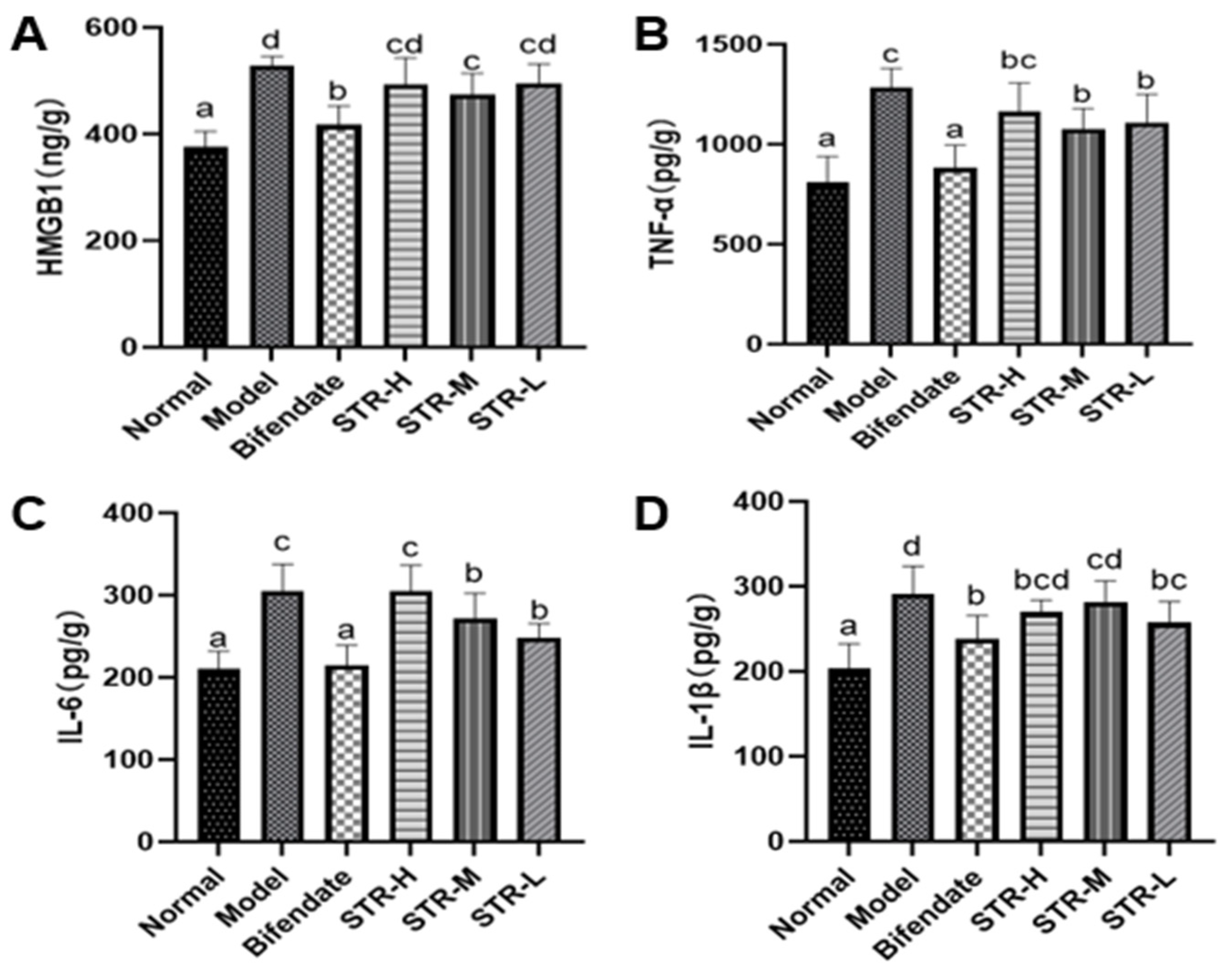
3.5. STR Treatment Reduces the CCL4-Induced Inflammatory Response
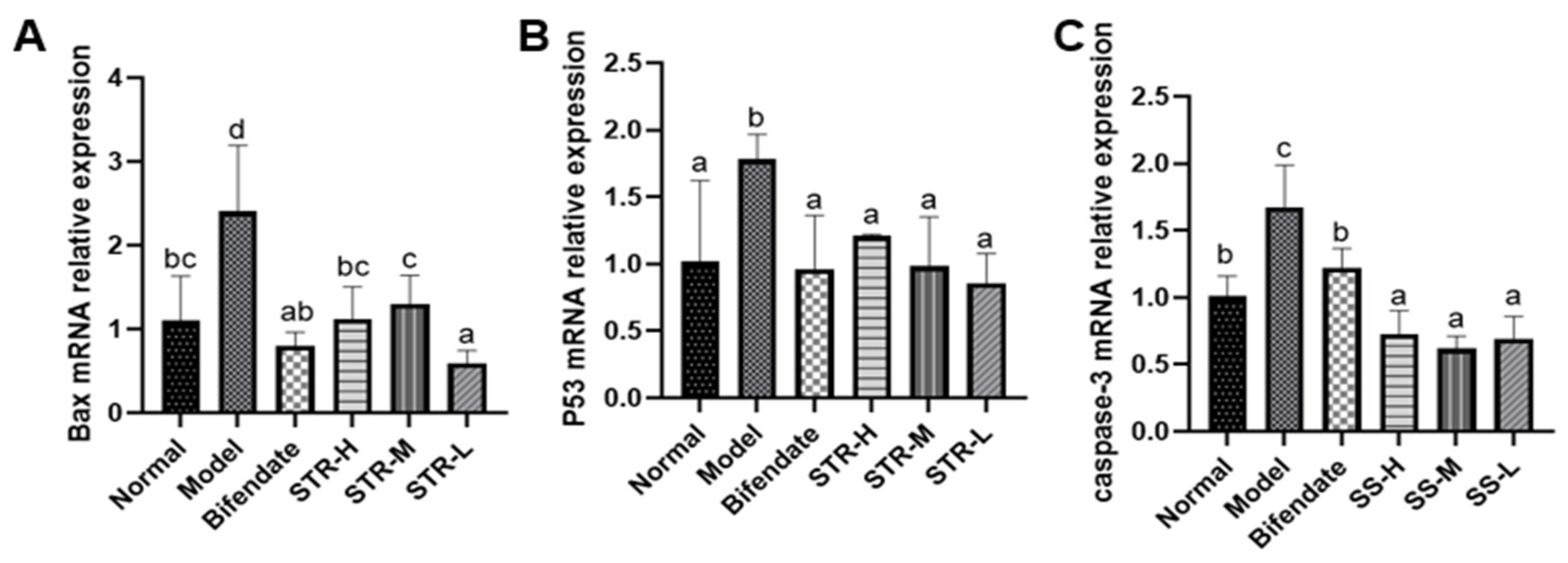
3.6. Effect of STR on CCL4-Induced Hepatocyte Apoptosis
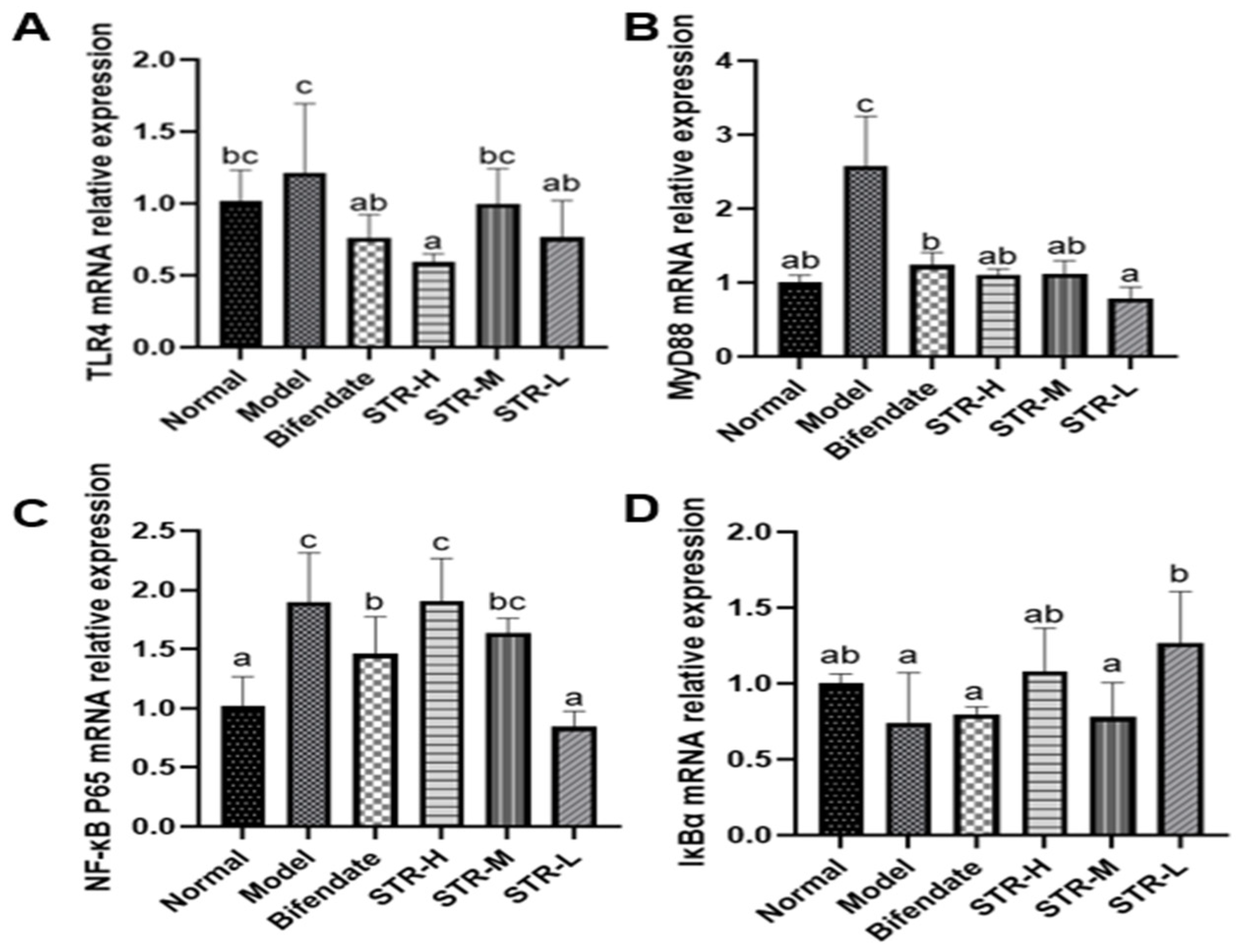
3.7. Effects of STR on the TLR4/MyD88/NF-κB Pathway in CCL4-Induced Liver Injury
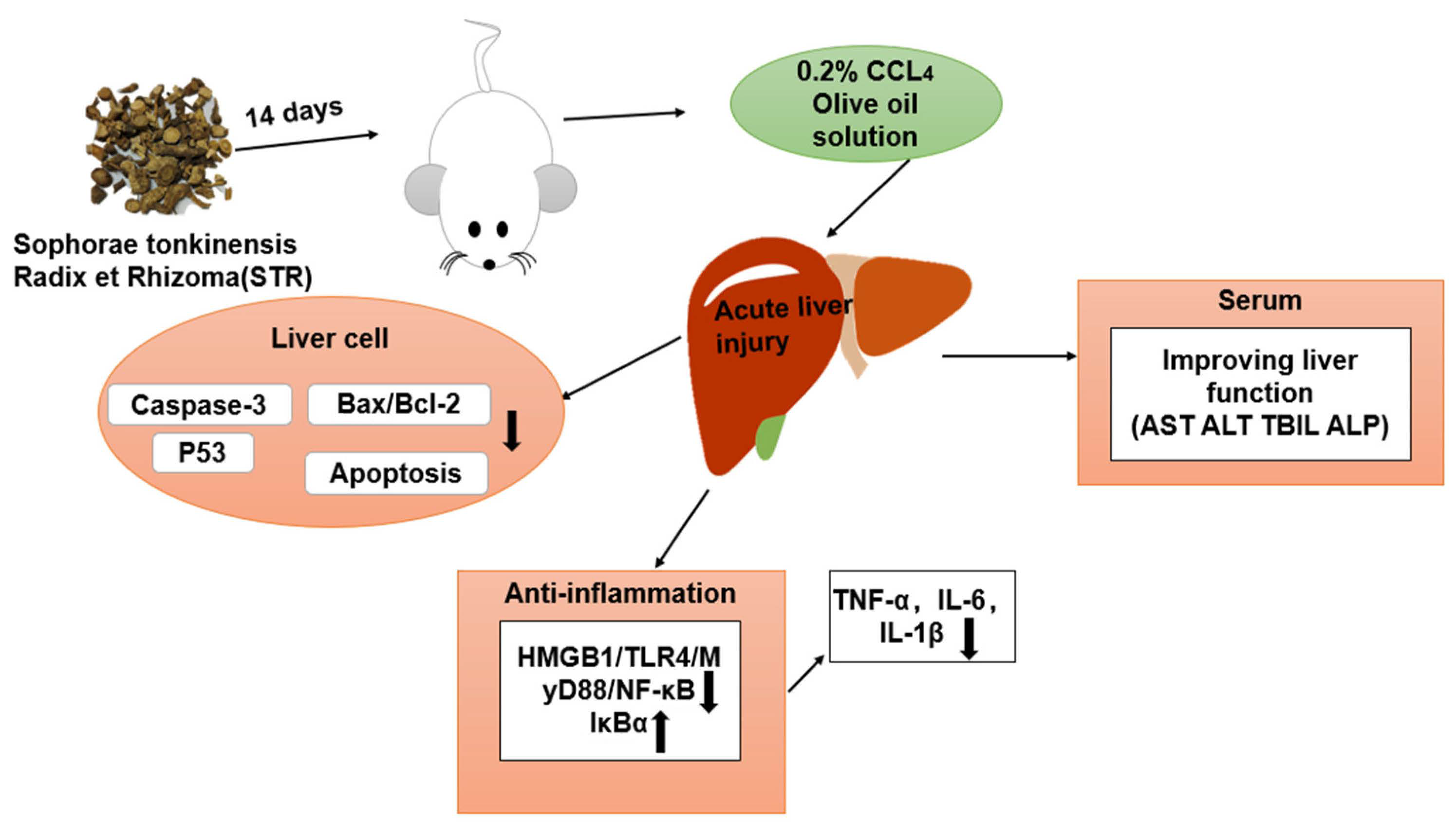
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell. Biol. 2004, 5, 836–847. [Google Scholar] [PubMed]
- Wu, Z.; Han, M.; Chen, T.; Yan, W.; Ning, Q. Acute liver failure: Mechanisms of immune-mediated liver injury. Liver Int. 2010, 30, 782–794. [Google Scholar] [CrossRef]
- Stocks, B.; Gonzalez-Franquesa, A.; Borg, M.L.; Björnholm, M.; Niu, L.; Zierath, J.R.; Deshmukh, A.S. Integrated Liver and Plasma Proteomics in Obese Mice Reveals Complex Metabolic Regulation. Mol. Cell. Proteomics 2022, 21, 100207. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Hu, Y.; Hu, F.; Li, H.; Chen, Y.; Zhang, J.; Yu, Y.; Jiang, S.; Weng, Q.; Tian, S.; et al. Hepatocyte glutathione S-transferase mu 2 prevents non-alcoholic steatohepatitis by suppressing ASK1 signaling. J. Hepatol. 2022, 76, 407–419. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Liu, M.; Zhou, X.; Wang, M.; Cao, K.; Jin, S.; Shan, A.; Feng, X. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem. Toxicol. 2022, 161, 112823. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jin, Z.; Lin, D.; Shen, T.; Zhang, J.; Li, D.; Wang, X.; Zhang, C.; Lin, Z.; Li, X.; et al. FGF21 alleviates acute liver injury by inducing the SIRT1-autophagy signalling pathway. J. Cell. Mol. Med. 2022, 26, 868–879. [Google Scholar] [CrossRef]
- Gao, Z.; Shen, Z.; Wu, J.; Song, Y.; Liu, N.; Deng, Q.; Xie, Y.; Liu, J. Interleukin-33 mediates both immune-related and non-immune-related inhibitory effects against hepatitis B virus. Antivir. Res. 2022, 206, 105404. [Google Scholar] [CrossRef]
- Xie, S.-Z.; Zhai, X.-Y.; Xi, S.-Y.; Qiu, Y.-K.; Zhang, Y.-M.; Kong, X.-J.; Li, Y.-H.; Zhu, L.; Wang, Z.; Zhang, S.-G.; et al. The Protective Effects of Zornia. diphylla (L.) Pers. Against Acute Liver Injury Induced by Carbon Tetrachloride in Mice. Front. Pharmacol. 2021, 12, 764282. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Zeh, H.J.; Lotze, M.T. High-mobility group box 1, oxidative stress, and disease. Antioxid. Redox. Signal. 2011, 14, 1315–1335. [Google Scholar] [CrossRef]
- Andersson, U.; Tracey, K.J. HMGB1 is a therapeutic target for sterile inflammation and infection. Annu. Rev. Immunol. 2011, 29, 139–162. [Google Scholar] [CrossRef]
- Jaeschke, H.; McGill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Lundbäck, P.; Lea, J.D.; Sowinska, A.; Ottosson, L.; Fürst, C.M.; Steen, J.; Aulin, C.; Clarke, J.I.; Kipar, A.; Klevenvall, L.; et al. A novel high mobility group box 1 neutralizing chimeric antibody attenuates drug-induced liver injury and postinjury inflammation in mice. Hepatology 2016, 64, 1699–1710. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jin, Q.; Yao, Q.; Xu, B.; Li, Z.; Tu, C. Quercetin attenuates the activation of hepatic stellate cells and liver fibrosis in mice through modulation of HMGB1-TLR2/4-NF-κB signaling pathways. Toxicol. Lett. 2016, 261, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Antoine, D.J.; Lu, Y.; Arriazu, E.; Leung, T.-M.; Klepper, A.L.; Branch, A.D.; Fiel, M.I.; Nieto, N. High mobility group box-1 (HMGB1) participates in the pathogenesis of alcoholic liver disease (ALD). J. Biol. Chem. 2014, 289, 22672–22691. [Google Scholar] [CrossRef] [PubMed]
- Pirnie, R.; Gillespie, K.P.; Weng, L.; Mesaros, C.; Blair, I.A. Characterization and Quantification of Oxidized High Mobility Group Box 1 Proteoforms Secreted from Hepatocytes by Toxic Levels of Acetaminophen. Chem. Res. Toxicol. 2022, 35, 1893–1902. [Google Scholar] [CrossRef]
- Ding, J.; Cui, X.; Liu, Q. Emerging role of HMGB1 in lung diseases: Friend or foe. J. Cell. Mol. Med. 2017, 21, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.-Z.; Xiang, D.; Xie, C.; Li, J.-J.; Hu, J.-J.; He, H.-L.; Yuan, Y.-S.; Gao, J.; Han, W.; Yu, Y. Protective effect of recombinant human IL-1Ra on CCl4-induced acute liver injury in mice. World J. Gastroenterol. 2010, 16, 2771–2779. [Google Scholar] [CrossRef]
- Shi, H.; Han, W.; Shi, H.; Ren, F.; Chen, D.; Chen, Y.; Duan, Z. Augmenter of liver regeneration protects against carbon tetrachloride-induced liver injury by promoting autophagy in mice. Oncotarget 2017, 8, 12637–12648. [Google Scholar] [CrossRef]
- Zhang, D.-G.; Zhang, C.; Wang, J.-X.; Wang, B.-W.; Wang, H.; Zhang, Z.-H.; Chen, Y.-H.; Lu, Y.; Tao, L.; Wang, J.-Q.; et al. Obeticholic acid protects against carbon tetrachloride-induced acute liver injury and inflammation. Toxicol. Appl. Pharmacol. 2017, 314, 39–47. [Google Scholar] [CrossRef]
- Chen, M.; Huang, W.; Wang, C.; Nie, H.; Li, G.; Sun, T.; Yang, F.; Zhang, Y.; Shu, K.; Wang, C.; et al. High-mobility group box 1 exacerbates CCl₄-induced acute liver injury in mice. Clin. Immunol. 2014, 153, 56–63. [Google Scholar] [CrossRef]
- Dai, C.; Xiao, X.; Li, D.; Tun, S.; Wang, Y.; Velkov, T.; Tang, S. Chloroquine ameliorates carbon tetrachloride-induced acute liver injury in mice via the concomitant inhibition of inflammation and induction of apoptosis. Cell Death Dis. 2018, 9, 1164. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Luo, D.; Lin, D.-C.; Wang, W.-Z.; Li, C.-J.; Zhuo, X.-F.; Wu, Z.-N.; Zhang, Y.-B.; Wang, G.-C.; Li, Y.-L. Five matrine-type alkaloids from Sophora tonkinensis. J. Nat. Med. 2021, 75, 682–687. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Yang, C.; Du, Y.; Wang, W.; Sun, M.; Liu, J.; Ma, B.; Pang, L.; Zeng, Y.; Zhang, Z.; et al. A Systematic Review of the Pharmacology, Toxicology and Pharmacokinetics of Matrine. Front. Pharmacol. 2020, 11, 01067. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.-M.; Li, Y.-H.; Hua, J.; Huang, F.-P.; Wang, H.-S.; Liang, D. Antiviral Matrine-Type Alkaloids from the Rhizomes of Sophora tonkinensis. J. Nat. Prod. 2015, 78, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.-L.; He, C.-M.; Cheng, Z.-H.; Chen, D.-F. Flavonoids rather than alkaloids as the diagnostic constituents to distinguish Sophorae. Flavescentis. Radix from Sophorae. Tonkinensis Radix et Rhizoma: An HPLC fingerprint study. Chin. J. Nat. Med. 2018, 16, 951–960. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, L.-M.; Tong, Y.; You, Y. [Pharmacological effect and toxicology of Sophorae. Tonkinensis Radix et Rhizoma]. Zhongguo Zhong Yao Za Zhi 2017, 42, 2439–2442. [Google Scholar] [CrossRef]
- Han, F.-M.; Wang, L.-X.; Chen, Y.; Chen, L.-M.; Feng, W.-H.; Wang, J.-Y.; Liu, D.-W.; You, Y.; Tong, Y. [Simultaneous determination of seven alkaloids and three flavonoids in Sophorae. Tonkinensis Radix et Rhizoma by HPLC]. Zhongguo Zhong Yao Za Zhi 2016, 41, 4628–4634. [Google Scholar] [CrossRef]
- Wang, L.; Lu, J.; Sun, W.; Gu, Y.; Zhang, C.; Jin, R.; Li, L.; Zhang, Z.; Tian, X. Hepatotoxicity induced by radix Sophorae tonkinensis in mice and increased serum cholinesterase as a potential supplemental biomarker for liver injury. Exp. Toxicol. Pathol. 2017, 69, 193–202. [Google Scholar] [CrossRef]
- Cai, L.; Zou, S.; Liang, D.; Luan, L. Structural characterization, antioxidant and hepatoprotective activities of polysaccharides from Sophorae tonkinensis Radix. Carbohydr. Polym. 2018, 184, 354–365. [Google Scholar] [CrossRef]
- Zhao, Q.; Wei, M.; Zhang, S.; Huang, Z.; Lu, B.; Ji, L. The water extract of Sophorae tonkinensis Radix et Rhizoma alleviates non-alcoholic fatty liver disease and its mechanism. Phytomedicine 2020, 77, 153270. [Google Scholar] [CrossRef]
- Gan, P.; Ding, L.; Hang, G.; Xia, Q.; Huang, Z.; Ye, X.; Qian, X. Oxymatrine Attenuates Dopaminergic Neuronal Damage and Microglia-Mediated Neuroinflammation Through Cathepsin D-Dependent HMGB1/TLR4/NF-κB Pathway in Parkinson’s Disease. Front. Pharmacol. 2020, 11, 776. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Chen, H.; Liu, H.; Yang, X.; Guo, P.; Cao, X.; Cai, Y.; Xu, H.; Yang, J. Structure characterization and antioxidant activity analysis of polysaccharides from Lanzhou Lily. Front. Nutr. 2022, 9, 976607. [Google Scholar] [CrossRef] [PubMed]
- Ti, Y.; Zhang, Y.; Ban, Y.; Wang, X.; Hou, Y.; Song, Z. Polysaccharide from Borani by subcritical water with different temperatures and investigation of its physicochemical properties and antioxidant activity. Front. Nutr. 2022, 9, 982695. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Kim, S.-W.; Oh, S.-A.; Seol, S.-I.; Davaanyam, D.; Lee, J.-K. Cytosolic HMGB1 Mediates LPS-Induced Autophagy in Microglia by Interacting with NOD2 and Suppresses Its Proinflammatory Function. Cells 2022, 11, 2410. [Google Scholar] [CrossRef]
- Liu, S.; Kang, W.; Mao, X.; Ge, L.; Du, H.; Li, J.; Hou, L.; Liu, D.; Yin, Y.; Liu, Y.; et al. Melatonin mitigates aflatoxin B1-induced liver injury via modulation of gut microbiota/intestinal FXR/liver TLR4 signaling axis in mice. J. Pineal Res. 2022, 73, e12812. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Wang, D.; Hu, Y.; Li, W.; Liu, F.; Zhu, X.; Li, X.; Zhang, H.; Bai, H.; Yang, Q.; et al. Serum amyloid A1 exacerbates hepatic steatosis via TLR4-mediated NF-κB signaling pathway. Mol. Metab. 2022, 59, 101462. [Google Scholar] [CrossRef]
- Mokdad, A.A.; Lopez, A.D.; Shahraz, S.; Lozano, R.; Mokdad, A.H.; Stanaway, J.; Murray, C.J.L.; Naghavi, M. Liver cirrhosis mortality in 187 countries between 1980 and 2010: A systematic analysis. BMC Med. 2014, 12, 145. [Google Scholar] [CrossRef]
- Hassan, A.; Fontana, R.J. The diagnosis and management of idiosyncratic drug-induced liver injury. Liver Int. 2019, 39, 31–41. [Google Scholar] [CrossRef]
- Meyerson, C.; Naini, B.V. Something old, something new: Liver injury associated with total parenteral nutrition therapy and immune checkpoint inhibitors. Hum. Pathol. 2020, 96, 39–47. [Google Scholar] [CrossRef]
- Berger, M.L.; Bhatt, H.; Combes, B.; Estabrook, R.W. CCl4-induced toxicity in isolated hepatocytes: The importance of direct solvent injury. Hepatology 1986, 6, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Nwidu, L.L.; Elmorsy, E.; Oboma, Y.I.; Carter, W.G. Hepatoprotective and antioxidant activities of leaf and stem extracts against carbon tetrachloride-induced hepatotoxicity. J. Taibah. Univ. Med. Sci. 2018, 13, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, Y.; Wen, S.; Li, Q.; Chen, R.; Lai, X.; Zhang, Z.; Zhou, Z.; Xie, Y.; Zheng, X.; et al. Extract of Jasminum. grandiflorum L. alleviates CCl-induced liver injury by decreasing inflammation, oxidative stress and hepatic CYP2E1 expression in mice. Biomed. Pharmacother. 2022, 152, 113255. [Google Scholar] [CrossRef]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 cytokine: An overview of the immune regulation, immune dysregulation, and therapeutic approach. Int. Immunopharmacol. 2022, 111, 109130. [Google Scholar] [CrossRef]
- Zhuang, Y.; Li, Y.; Li, X.; Xie, Q.; Wu, M. Atg7 Knockdown Augments Concanavalin A-Induced Acute Hepatitis through an ROS-Mediated p38/MAPK Pathway. PLoS ONE 2016, 11, e0149754. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liu, J.; Chen, T.-M.; Lan, Q.; Zhang, Q.-Y.; Liu, B.; Dai, D.; Zhang, W.-D.; Hu, L.-P.; Zhu, R.-Z. Dihydromyricetin alleviates carbon tetrachloride-induced acute liver injury via JNK-dependent mechanism in mice. World J. Gastroenterol. 2015, 21, 5473–5481. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Lin, B.; Pan, J.F.; Liong, E.C.; Xu, A.M.; Youdim, M.; Fung, M.L.; So, K.F.; Tipoe, G.L. Inhibition of caspase-9 aggravates acute liver injury through suppression of cytoprotective autophagy. Sci. Rep. 2016, 6, 32447. [Google Scholar] [CrossRef]
- Dai, C.; Li, J.; Tang, S.; Li, J.; Xiao, X. Colistin-induced nephrotoxicity in mice involves the mitochondrial, death receptor, and endoplasmic reticulum pathways. Antimicrob. Agents Chemother. 2014, 58, 4075–4085. [Google Scholar] [CrossRef]
- Wang, K. Autophagy and apoptosis in liver injury. Cell Cycle 2015, 14, 1631–1642. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Desouky, E.M.; Hozayen, W.G.; Bin-Jumah, M.; El-Nahass, E.-S.; Soliman, H.A.; Farghali, A.A. Mesoporous Silica Nanoparticles Trigger Liver and Kidney Injury and Fibrosis Via Altering TLR4/NF-κB, JAK2/STAT3 and Nrf2/HO-1 Signaling in Rats. Biomolecules 2019, 9, 528. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Y.; Pei, C.; Liang, J.; Ding, P.; Chen, S.; Hou, S.-Z. Monotropein alleviates secondary liver injury in chronic colitis by regulating TLR4/NF-κB signaling and NLRP3 inflammasome. Eur. J. Pharmacol. 2020, 883, 173358. [Google Scholar] [CrossRef]
- Qu, X.-Q.; Chen, Q.-F.; Shi, Q.-Q.; Luo, Q.-Q.; Zheng, S.-Y.; Li, Y.-H.; Bai, L.-Y.; Gan, S.; Zhou, X.-Y. Hepatocyte-Conditional Knockout of Phosphatidylethanolamine Binding Protein 4 Aggravated LPS/D-GalN-Induced Acute Liver Injury the TLR4/NF-κB Pathway. Front. Immunol. 2022, 13, 901566. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| GAPDH | CAA GAA GGT GGT GAA GCA GGC | CCA GGA AAT GAG CTT GAC AAA G |
| Bax | TGA GCG AGT GTC TCC GGC GAA T | GCA CTT TAG TGC ACA GGG CCT TG |
| P53 | CGC CGA CCT ATC CTT ACC ATC ATC | GGC AGT TCA GGG CAA AGG AC |
| Caspase3 | TGGGACTGATGAGGAGA | ACTGGATGAACCACGAC |
| MyD88 | TCATGTTCTCCATACCCTTGGT | AAACTGCGAGTGGGGTCAG |
| NF-κB P65 | GCCTCTGGCGAATGGCTTTA | TGCTTCGGCTGTTCGATGAT |
| IκBα | ATGCCAGAACGAGATAGTGAGC | AGGTGGCGCAGAAGTAGGT |
| TLR4 | TCTGGGGAGGCACATCTTCT | AGGTCCAAGTTGCCGTTTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Liu, A.; Liu, G.; Yang, C.; Zhou, Q.; Li, H.; Yang, H.; Yang, M.; Li, G.; Si, H.; et al. Protective Effects of Sophorae tonkinensis Gagnep. (Fabaceae) Radix et Rhizoma Water Extract on Carbon Tetrachloride-Induced Acute Liver Injury. Molecules 2022, 27, 8650. https://doi.org/10.3390/molecules27248650
Zhou C, Liu A, Liu G, Yang C, Zhou Q, Li H, Yang H, Yang M, Li G, Si H, et al. Protective Effects of Sophorae tonkinensis Gagnep. (Fabaceae) Radix et Rhizoma Water Extract on Carbon Tetrachloride-Induced Acute Liver Injury. Molecules. 2022; 27(24):8650. https://doi.org/10.3390/molecules27248650
Chicago/Turabian StyleZhou, Congcong, Aijing Liu, Gengsong Liu, Cheng Yang, Qiaoyan Zhou, Haizhu Li, Hongchun Yang, Mengmeng Yang, Gonghe Li, Hongbin Si, and et al. 2022. "Protective Effects of Sophorae tonkinensis Gagnep. (Fabaceae) Radix et Rhizoma Water Extract on Carbon Tetrachloride-Induced Acute Liver Injury" Molecules 27, no. 24: 8650. https://doi.org/10.3390/molecules27248650
APA StyleZhou, C., Liu, A., Liu, G., Yang, C., Zhou, Q., Li, H., Yang, H., Yang, M., Li, G., Si, H., & Ou, C. (2022). Protective Effects of Sophorae tonkinensis Gagnep. (Fabaceae) Radix et Rhizoma Water Extract on Carbon Tetrachloride-Induced Acute Liver Injury. Molecules, 27(24), 8650. https://doi.org/10.3390/molecules27248650