

Four Chemotherapeutic Compounds That Limit Blood-Brain-Barrier Invasion by Toxoplasma gondii

 ,
, 

Abstract
:1. Introduction
2. Results
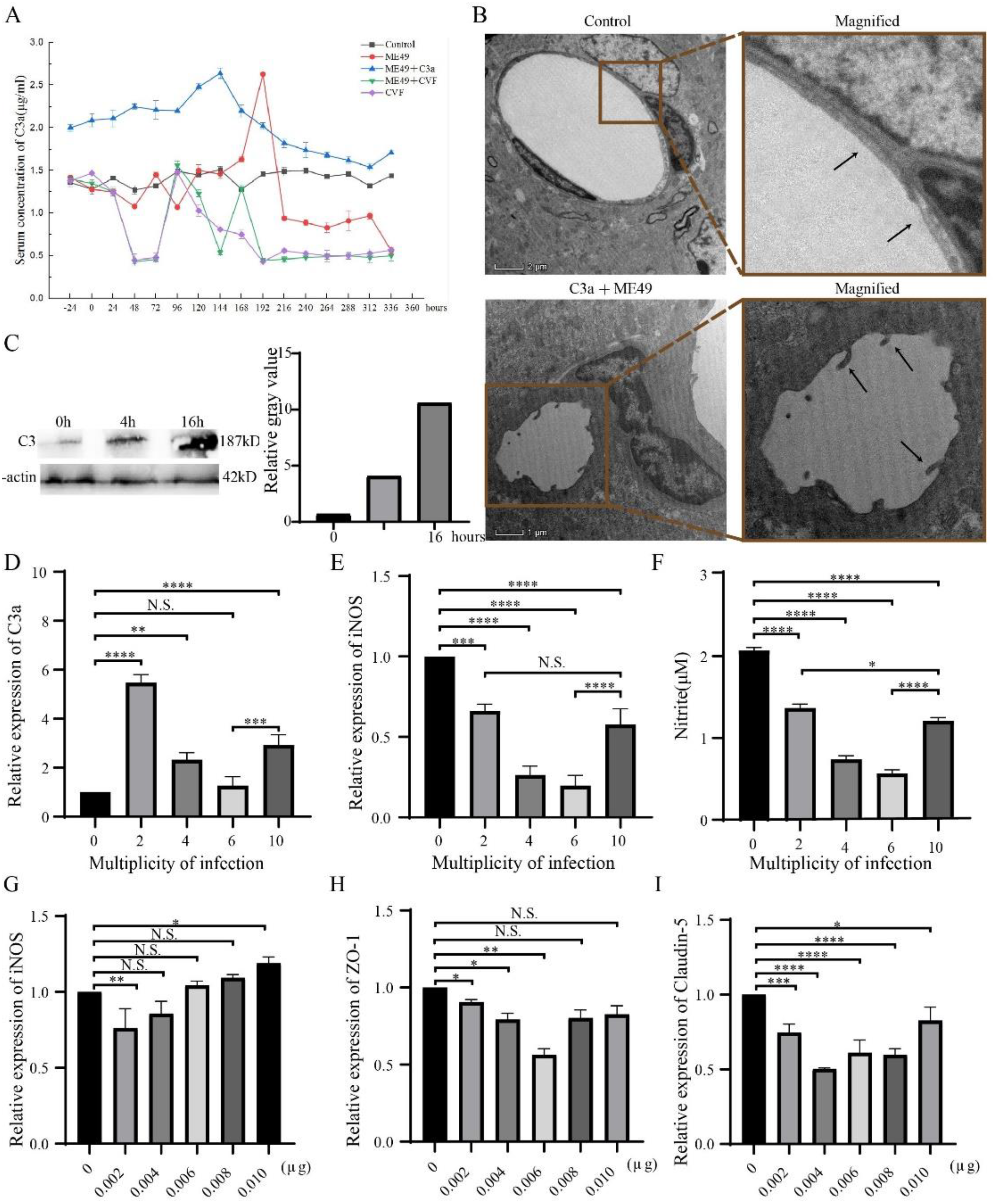
2.1. T. gondii Increased C3 Expression and BBB Disruption
2.2. SB290157 Combined with C3a at Low Concentration Inhibited the Expression of C3a, iNOS, and Vimentin
2.3. NSC23766 Promoted TJ Proteins Express
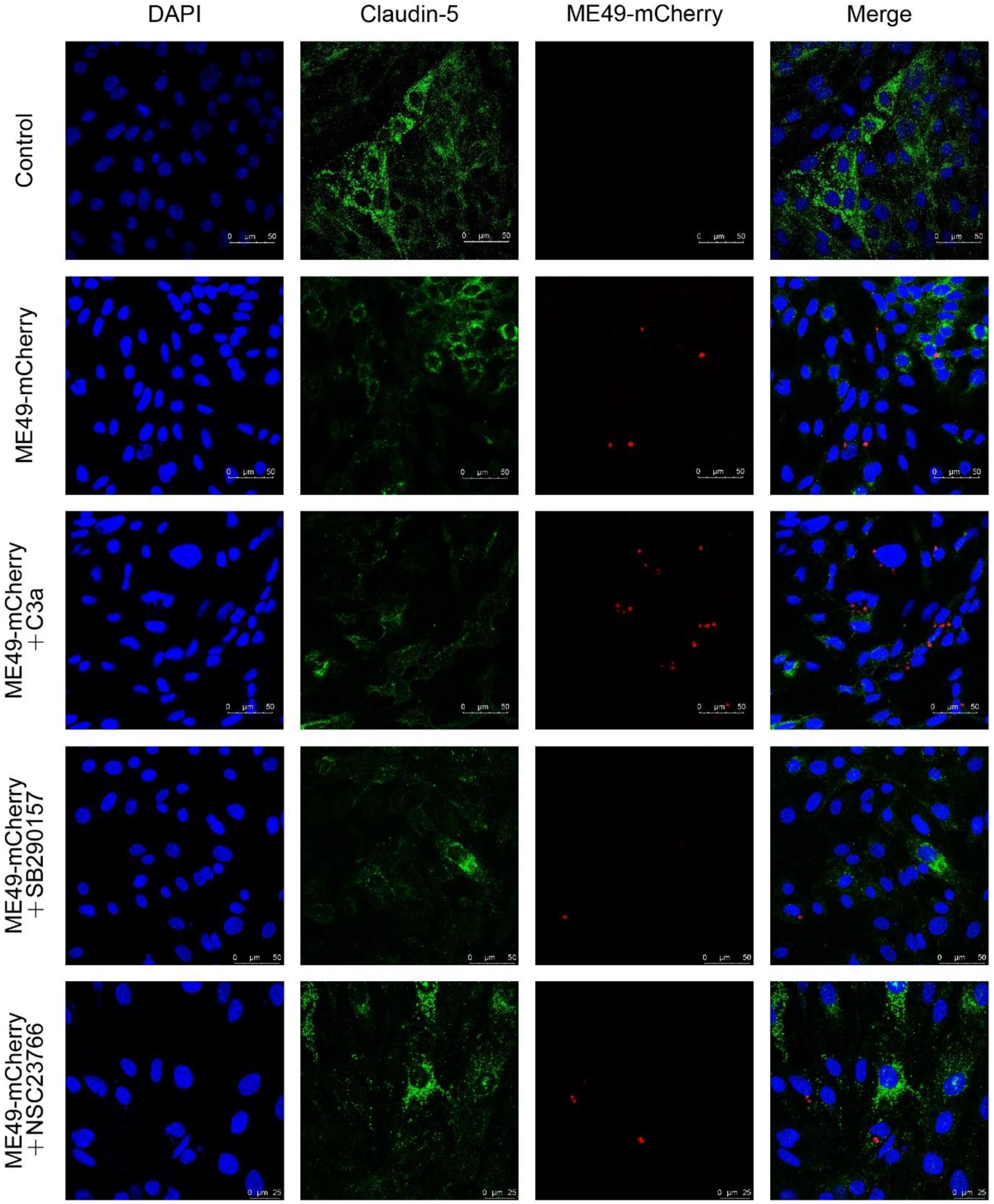
2.4. SB290157 and NSC23766 Inhibited T. gondii Invasion into bEnd3 Cells
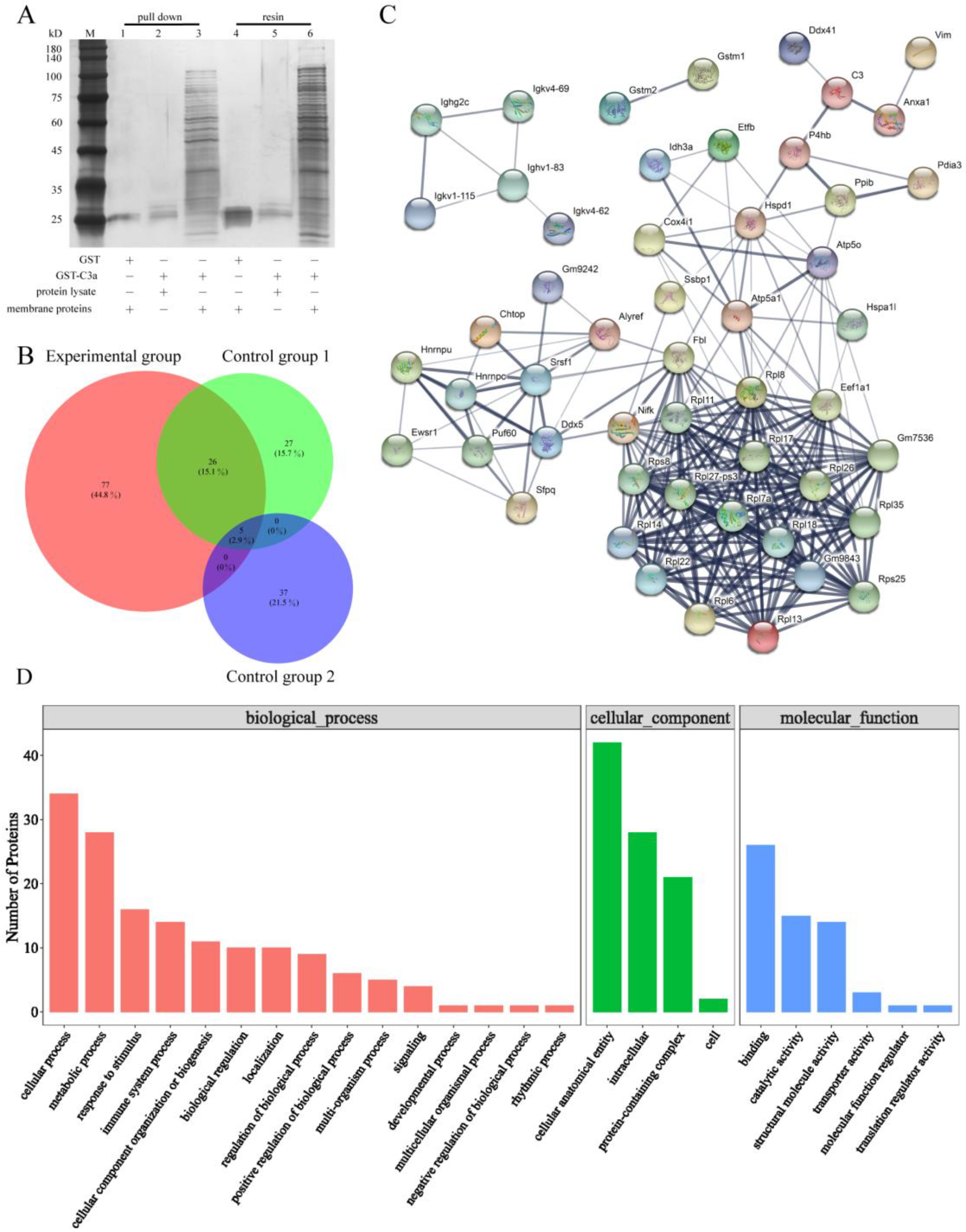
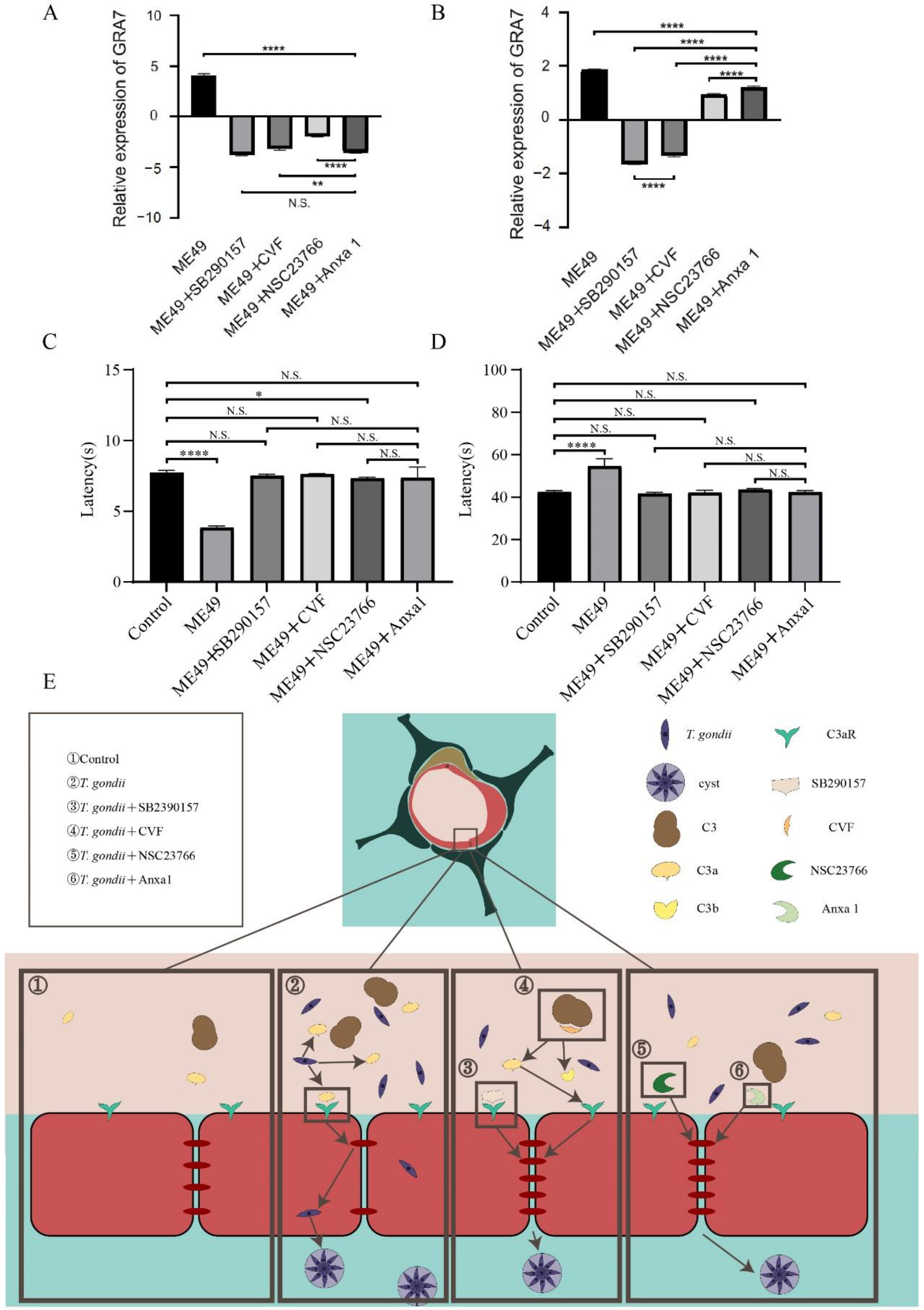
2.5. Anxa1 Was Picked out as a Protein Which Could Directly Act on C3a
2.6. Four Chemotherapeutic Compounds Inhibited T. gondii from Entering the Mice Brains, Protecting Mice from Mental Cognition Impairment
3. Discussion
4. Materials and Methods
4.1. Experimental Animal
4.2. Chemotherapeutic Drugs
4.3. Parasites and Cells
4.4. ELISA and Electron Microscopy
4.5. Western Blotting
4.6. Measurement of Nitric Oxide
4.7. Pull down and LC-MS
4.8. Immunofluorescence
4.9. Quantitative PCR
4.10. Rotarod Test and Morris Water Maze
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Courret, N.; Darche, S.; Sonigo, P.; Milon, G.; Buzoni-Gâtel, D.; Tardieux, I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006, 107, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Pittman, K.J.; Knoll, L.J. Long-Term Relationships: The Complicated Interplay between the Host and the Developmental Stages of Toxoplasma gondii during Acute and Chronic Infections. Microbiol. Mol. Biol. Rev. 2015, 79, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Roberts, C.W.; Alexander, J. Kinetics of cytokine mRNA production in the brains of mice with progressive toxoplasmic encephalitis. Eur. J. Immunol. 1992, 22, 2317–2322. [Google Scholar] [CrossRef]
- Wilson, E.H.; Hunter, C.A. The role of astrocytes in the immunopathogenesis of toxoplasmic encephalitis. Int. J. Parasitol. 2004, 34, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Sibley, L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors. Nat. Rev. Microbiol. 2012, 10, 766–778. [Google Scholar] [CrossRef]
- Luft, B.J.; Remington, J.S. Toxoplasmic encephalitis in AIDS. Clin. Infect. Dis. 1992, 15, 211–222. [Google Scholar] [CrossRef]
- Xiao, J.; Kannan, G.; Jones-Brando, L.; Brannock, C.; Krasnova, I.N.; Cadet, J.L.; Pletnikov, M.; Yolken, R.H. Sex-specific changes in gene expression and behavior induced by chronic Toxoplasma infection in mice. Neuroscience 2012, 206, 39–48. [Google Scholar] [CrossRef]
- Vyas, A.; Kim, S.K.; Giacomini, N.; Boothroyd, J.C.; Sapolsky, R.M. Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc. Natl. Acad. Sci. USA 2007, 104, 6442–6447. [Google Scholar] [CrossRef]
- Webster, J.P. The effect of Toxoplasma gondii on animal behavior: Playing cat and mouse. Schizophr. Bull. 2007, 33, 752–756. [Google Scholar] [CrossRef]
- Hamdani, N.; Daban-Huard, C.; Lajnef, M.; Richard, J.R.; Delavest, M.; Godin, O.; Le Guen, E.; Vederine, F.E.; Lépine, J.P.; Jamain, S.; et al. Relationship between Toxoplasma gondii infection and bipolar disorder in a French sample. J. Affect. Disord. 2013, 148, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Chien, W.C.; Chung, C.H.; Su, R.Y.; Lai, C.Y.; Yang, C.C.; Tzeng, N.S. Risk of dementia in patients with toxoplasmosis: A nationwide, population-based cohort study in Taiwan. Parasit. Vectors 2021, 14, 435. [Google Scholar] [CrossRef]
- Wei, H.X.; Wei, S.S.; Lindsay, D.S.; Peng, H.J. A Systematic Review and Meta-Analysis of the Efficacy of Anti-Toxoplasma gondii Medicines in Humans. PLoS ONE 2015, 10, e0138204. [Google Scholar] [CrossRef]
- Hajj, R.E.; Tawk, L.; Itani, S.; Hamie, M.; Ezzeddine, J.; El Sabban, M.; El Hajj, H. Toxoplasmosis: Current and Emerging Parasite Druggable Targets. Microorganisms 2021, 9, 2531. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, P.M.; Commodaro, A.G.; Grigg, M.E. A Protective and Pathogenic Role for Complement During Acute Toxoplasma gondii Infection. Front. Cell. Infect. Microbiol. 2021, 11, 634610. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Reis, E.S.; Mastellos, D.C.; Gros, P.; Lambris, J.D. Complement component C3—The “Swiss Army Knife” of innate immunity and host defense. Immunol. Rev. 2016, 274, 33–58. [Google Scholar] [CrossRef]
- Fuhrman, S.A.; Joiner, K.A. Toxoplasma gondii: Mechanism of resistance to complement-mediated killing. J. Immunol. 1989, 142, 940–947. [Google Scholar]
- Sikorski, P.M.; Commodaro, A.G.; Grigg, M.E. Toxoplasma gondii Recruits Factor H and C4b-Binding Protein to Mediate Resistance to Serum Killing and Promote Parasite Persistence in vivo. Front. Immunol. 2020, 10, 3105. [Google Scholar] [CrossRef]
- Huang, W.Y.; Wang, Y.P.; Mahmmod, Y.S.; Wang, J.J.; Liu, T.H.; Zheng, Y.X.; Zhou, X.; Zhang, X.X.; Yuan, Z.G. A Double-Edged Sword: Complement Component 3 in Toxoplasma gondii Infection. Proteomics 2019, 19, e1800271. [Google Scholar] [CrossRef]
- Ricklin, D.; Lambris, J.D. Compstatin: A complement inhibitor on its way to clinical application. Adv. Exp. Med. Biol. 2008, 632, 273–292. [Google Scholar] [CrossRef]
- Köhl, J. Drug evaluation: The C5a receptor antagonist PMX-53. Curr. Opin. Mol. Ther. 2006, 8, 529–538. [Google Scholar] [PubMed]
- Vogel, C.W.; Fritzinger, D.C. Cobra venom factor: Structure, function, and humanization for therapeutic complement depletion. Toxicon 2010, 56, 1198–1222. [Google Scholar] [CrossRef] [PubMed]
- Kock, M.A.; Hew, B.E.; Bammert, H.; Fritzinger, D.C.; Vogel, C.W. Structure and function of recombinant cobra venom factor. J. Biol. Chem. 2004, 279, 30836–30843. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, C.G.; Müller-Eberhard, H.J.; Aikin, B.S. Depletion of plasma complement in vivo by a protein of cobra venom: Its effect on various immunologic reactions. J. Immunol. 1970, 105, 55–69. [Google Scholar] [PubMed]
- Maillard, J.L.; Zarco, R.M. Decomplementization by a factor extracted from cobra venom. Effect on several immune reactions of the guinea pig and rat. Ann. L’institut Pasteur 1968, 114, 756–774. [Google Scholar]
- Ames, R.S.; Lee, D.; Foley, J.J.; Jurewicz, A.J.; Tornetta, M.A.; Bautsch, W.; Settmacher, B.; Klos, A.; Erhard, K.F.; Cousins, R.D.; et al. Identification of a selective nonpeptide antagonist of the anaphylatoxin C3a receptor that demonstrates antiinflammatory activity in animal models. J. Immunol. 2001, 166, 6341–6348. [Google Scholar] [CrossRef]
- Bao, L.; Osawe, I.; Haas, M.; Quigg, R.J. Signaling through up-regulated C3a receptor is key to the development of experimental lupus nephritis. J. Immunol. 2005, 175, 1947–1955. [Google Scholar] [CrossRef]
- Ahmad, S.; Pandya, C.; Kindelin, A.; Bhatia, K.; Chaudhary, R.; Dwivedi, A.K.; Eschbacher, J.M.; Liu, Q.; Waters, M.F.; Hoda, M.N.; et al. C3a receptor antagonist therapy is protective with or without thrombolysis in murine thromboembolic stroke. Br. J. Pharmacol. 2020, 177, 2466–2477. [Google Scholar] [CrossRef]
- Veluthakal, R.; Sidarala, V.; Kowluru, A. NSC23766, a Known Inhibitor of Tiam1-Rac1 Signaling Module, Prevents the Onset of Type 1 Diabetes in the NOD Mouse Model. Cell Physiol. Biochem. 2016, 39, 760–767. [Google Scholar] [CrossRef]
- Wei, H.; Zhou, L.; Wu, S.; Li, D.; Deng, S.; Peng, H. Host cell Rac1 GTPase facilitates Toxoplasma gondii invasion. Sci. China Life Sci. 2020, 63, 610–612. [Google Scholar] [CrossRef]
- Cardoso, F.L.; Brites, D.; Brito, M.A. Looking at the blood-brain barrier: Molecular anatomy and possible investigation approaches. Brain Res. Rev. 2010, 64, 328–363. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.L.; Staddon, J.M. The cell biology of the blood-brain barrier. Annu. Rev. Neurosci. 1999, 22, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Dohgu, S.; Takata, F.; Nishioku, T.; Nakashima, A.; Futagami, K.; Yamauchi, A.; Kataoka, Y. Paracellu-lar barrier and tight junction protein expression in the immortalized brain endothelial cell lines bEND.3, bEND.5 and mouse brain endothelial cell 4. Biol. Pharm. Bull. 2013, 36, 492–495. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Butt, A.M.; Jones, H.C.; Abbott, N.J. Electrical resistance across the blood-brain barrier in anaesthetized rats: A developmen-tal study. J. Physiol. 1990, 429, 47–62. [Google Scholar] [CrossRef]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Itallie, C.M.V.; Anderson, J.M. Claudins and epithelial paracellular transport. Annu. Rev. Physiol. 2006, 68, 403–429. [Google Scholar] [CrossRef]
- Balda, M.S.; Matter, K. Epithelial cell adhesion and the regulation of gene expression. Trends Cell Biol. 2003, 13, 310–318. [Google Scholar] [CrossRef]
- Halonen, S.K.; Weidner, E. Overcoating of Toxoplasma parasitophorous vacuoles with host cell vimentin type intermediate filaments. J. Eukaryot. Microbiol. 1994, 41, 65–71. [Google Scholar] [CrossRef]
- Selseleh, M.; Keshavarz, H.; Mohebali, M.; Shojaee, S.; Selseleh, M.; Eshragian, M.R.; Mansouri, F.; Modarressi, M.H. Production and evaluation of Toxoplasma gondii recombinant GRA7 for serodiagnosis of human infections. Korean J. Parasitol. 2012, 50, 233–238. [Google Scholar] [CrossRef]
- Fox, B.A.; Gigley, J.P.; Bzik, D.J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. Int. J. Parasitol. 2004, 34, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I. Opening of the blood-brain barrier by acute hypertension. Exp. Neurol. 1976, 52, 467–479. [Google Scholar] [CrossRef]
- Adams, L.B.; Hibbs, J.B., Jr.; Taintor, R.R.; Krahenbuhl, J.L. Microbiostatic effect of murine-activated macrophages for Toxoplasma gondii. Role for synthesis of inorganic nitrogen oxides from L-arginine. J. Immunol. 1990, 144, 2725–2729. [Google Scholar] [PubMed]
- Luo, Z.Z.; Gao, Y.; Sun, N.; Zhao, Y.; Wang, J.; Tian, B.; Shi, J. Enhancing the interaction between annexin-1 and formyl peptide receptors regulates microglial activation to protect neurons from ischemia-like injury. J. Neuroimmunol. 2014, 276, 24–36. [Google Scholar] [CrossRef] [PubMed]
- McArthur, S.; Cristante, E.; Paterno, M.; Christian, H.; Roncaroli, F.; Gillies, G.E.; Solito, E. Annexin A1: A central player in the anti-inflammatory and neuroprotective role of microglia. J. Immunol. 2010, 185, 6317–6328. [Google Scholar] [CrossRef]
- Klenow, M.B.; Iversen, C.; Lund, F.W.; Mularski, A.; Heitmann, A.S.B.; Dias, C.; Nylandsted, J.; Simonsen, A.C. Annexins A1 and A2 Accumulate and Are Immobilized at Cross-Linked Membrane–Membrane Interfaces. Biochemistry 2021, 60, 1248–1259. [Google Scholar] [CrossRef]
- McNeil, A.K.; Rescher, U.; Gerke, V.; McNeil, P.L. Requirement for Annexin A1 in Plasma Membrane Repair. J. Biol. Chem. 2006, 281, 35202–35207. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhu, Y.; Liu, Z.J.; Ouyang, S. The emerging roles of the DDX41 protein in immunity and diseases. Protein Cell 2017, 8, 83–89. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, J.; Zhang, Q.; Xu, Q.; Lu, L.; Wang, J.; Xia, W. P4HB knockdown induces human HT29 colon cancer cell apoptosis through the generation of reactive oxygen species and inactivation of STAT3 signaling. Mol. Med. Rep. 2019, 19, 231–237. [Google Scholar] [CrossRef]
- Dyrskjøt, L.; Kruhøffer, M.; Thykjaer, T.; Marcussen, N.; Jensen, J.L.; Møller, K.; Ørntoft, T.F. Gene expression in the urinary bladder: A common carcinoma in situ gene expression signature exists disregarding histopathological classification. Cancer Res. 2004, 64, 4040–4048. [Google Scholar] [CrossRef]
- Sanchez-Carbayo, M.; Socci, N.D.; Lozano, J.; Saint, F.; Cordon-Cardo, C. Defining molecular profiles of poor outcome in patients with invasive bladder cancer using oligonucleotide microarrays. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.B.; Sapinoso, L.M.; Su, A.I.; Kern, S.G.; Wang-Rodriguez, J.; Moskaluk, C.A.; Frierson, H.F., Jr.; Hampton, G.M. Analysis of gene expression identifies candidate markers and pharmacological targets in prostate cancer. Cancer Res. 2001, 61, 5974–5978. [Google Scholar] [PubMed]
- Wang, Z.; Li, Y.; Cai, S.; Li, R.; Cao, G. Cannabinoid receptor 2 agonist attenuates blood-brain barrier damage in a rat model of intracerebral hemorrhage by activating the Rac1 pathway. Int. J. Mol. Med. 2018, 42, 2914–2922. [Google Scholar] [CrossRef]
- Jiang, Z.S.; Zhang, J.R. LncRNA SNHG5 enhances astrocytes and microglia viability via upregulating KLF4 in spinal cord injury. Int. J. Biol. Macromol. 2018, 120, 66–72. [Google Scholar] [CrossRef]
- Mu, K.; Yu, S.; Kitts, D.D. The Role of Nitric Oxide in Regulating Intestinal Redox Status and Intestinal Epithelial Cell Functionality. Int. J. Mol. Sci. 2019, 20, 1755. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Kumar, V.; Clark, R.J.; Lee, J.D.; Woodruff, T.M. The “C3aR Antagonist” SB290157 is a Partial C5aR2 Agonist. Front. Pharmacol. 2021, 11, 591398. [Google Scholar] [CrossRef] [PubMed]
- Parente, L.; Solito, E. Annexin 1: More than an anti-phospholipase protein. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2004, 53, 125–132. [Google Scholar] [CrossRef]
- Cristante, E.; McArthur, S.; Mauro, C.; Maggioli, E.; Romero, I.A.; Wylezinska-Arridge, M.; Couraud, P.O.; Lopez-Tremoleda, J.; Christian, H.C.; Weksler, B.B.; et al. Identification of an essential endogenous regulator of blood-brain barrier integrity, and its pathological and therapeutic implications. Proc. Natl. Acad. Sci. USA 2013, 110, 832–841. [Google Scholar] [CrossRef]
- Dobrowolski, J.M.; Sibley, L.D. Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell 1996, 84, 933–939. [Google Scholar] [CrossRef]
- Harun, M.S.R.; Taylor, M.; Zhu, X.; Elsheikha, H.M. Transcriptome Profiling of Toxoplasma gondii-Infected Human Cerebromicrovascular Endothelial Cell Response to Treatment with Monensin. Microorganisms 2020, 8, 842. [Google Scholar] [CrossRef]
- de Oliveira Cardoso, M.F.; Moreli, J.B.; Gomes, A.O.; de Freitas Zanon, C.; Silva, A.E.; Paulesu, L.R.; Ietta, F.; Mineo, J.R.; Ferro, E.A.; Oliani, S.M. Annexin A1 peptide is able to induce an anti-parasitic effect in human placental explants infected by Toxoplasma gondii. Microb. Pathog. 2018, 123, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|
| C3a | ATGACTCCCAGCACAAAGGG | CTCTCTTGCGGACCATCTCC |
| GRA7 | CTAACCACCGGCCAGAATGT | TACGTCCTCGTGAGACCCAT |
| iNOS | AGCTATGTGACCACGTCCAC | CACCGGGGGATGAGGAATAG |
| ZO-1 | TCTTGCAAAGTATCCCTTCTGT | GAAATCGTGCTGATGTGCCA |
| Claudin-5 | GTTAAGGCACGGGTAGCACT | CAACGATGTTGGCGAACCAG |
| β-Actb | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Z.; Yuan, H.; Wang, J.; Yang, Z.; Zhang, P.; Mahmmod, Y.S.; Wang, X.; Liu, T.; Song, Y.; Ren, Z.; et al. Four Chemotherapeutic Compounds That Limit Blood-Brain-Barrier Invasion by Toxoplasma gondii. Molecules 2022, 27, 5572. https://doi.org/10.3390/molecules27175572
Yan Z, Yuan H, Wang J, Yang Z, Zhang P, Mahmmod YS, Wang X, Liu T, Song Y, Ren Z, et al. Four Chemotherapeutic Compounds That Limit Blood-Brain-Barrier Invasion by Toxoplasma gondii. Molecules. 2022; 27(17):5572. https://doi.org/10.3390/molecules27175572
Chicago/Turabian StyleYan, Zijing, Hao Yuan, Junjie Wang, Zipeng Yang, Pian Zhang, Yasser S. Mahmmod, Xiaohu Wang, Tanghui Liu, Yining Song, Zhaowen Ren, and et al. 2022. "Four Chemotherapeutic Compounds That Limit Blood-Brain-Barrier Invasion by Toxoplasma gondii" Molecules 27, no. 17: 5572. https://doi.org/10.3390/molecules27175572
APA StyleYan, Z., Yuan, H., Wang, J., Yang, Z., Zhang, P., Mahmmod, Y. S., Wang, X., Liu, T., Song, Y., Ren, Z., Zhang, X.-X., & Yuan, Z.-G. (2022). Four Chemotherapeutic Compounds That Limit Blood-Brain-Barrier Invasion by Toxoplasma gondii. Molecules, 27(17), 5572. https://doi.org/10.3390/molecules27175572