
Neuroprotective Effect of Quercetin during Cerebral Ischemic Injury Involves Regulation of Essential Elements, Transition Metals, Cu/Zn Ratio, and Antioxidant Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
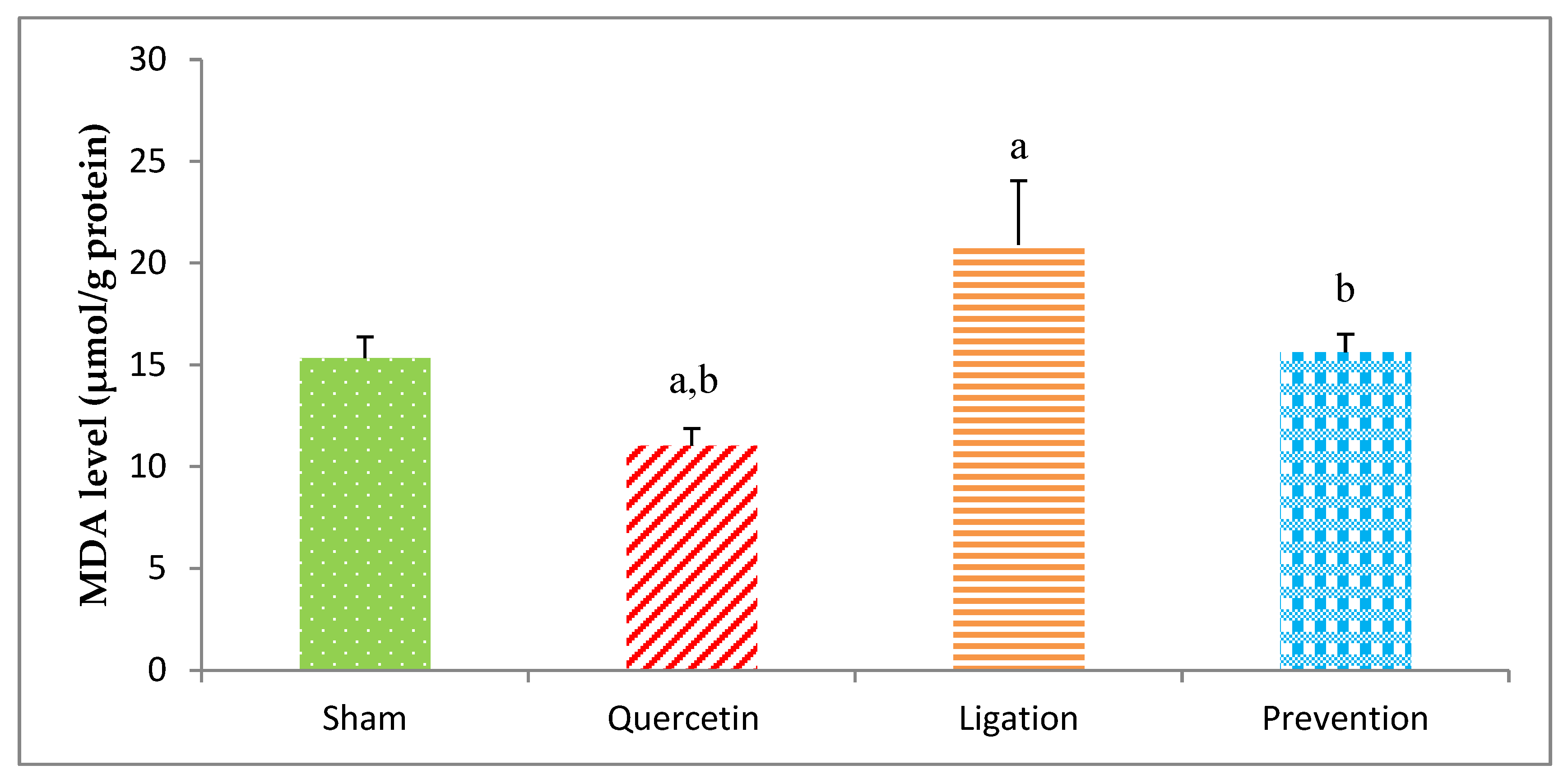
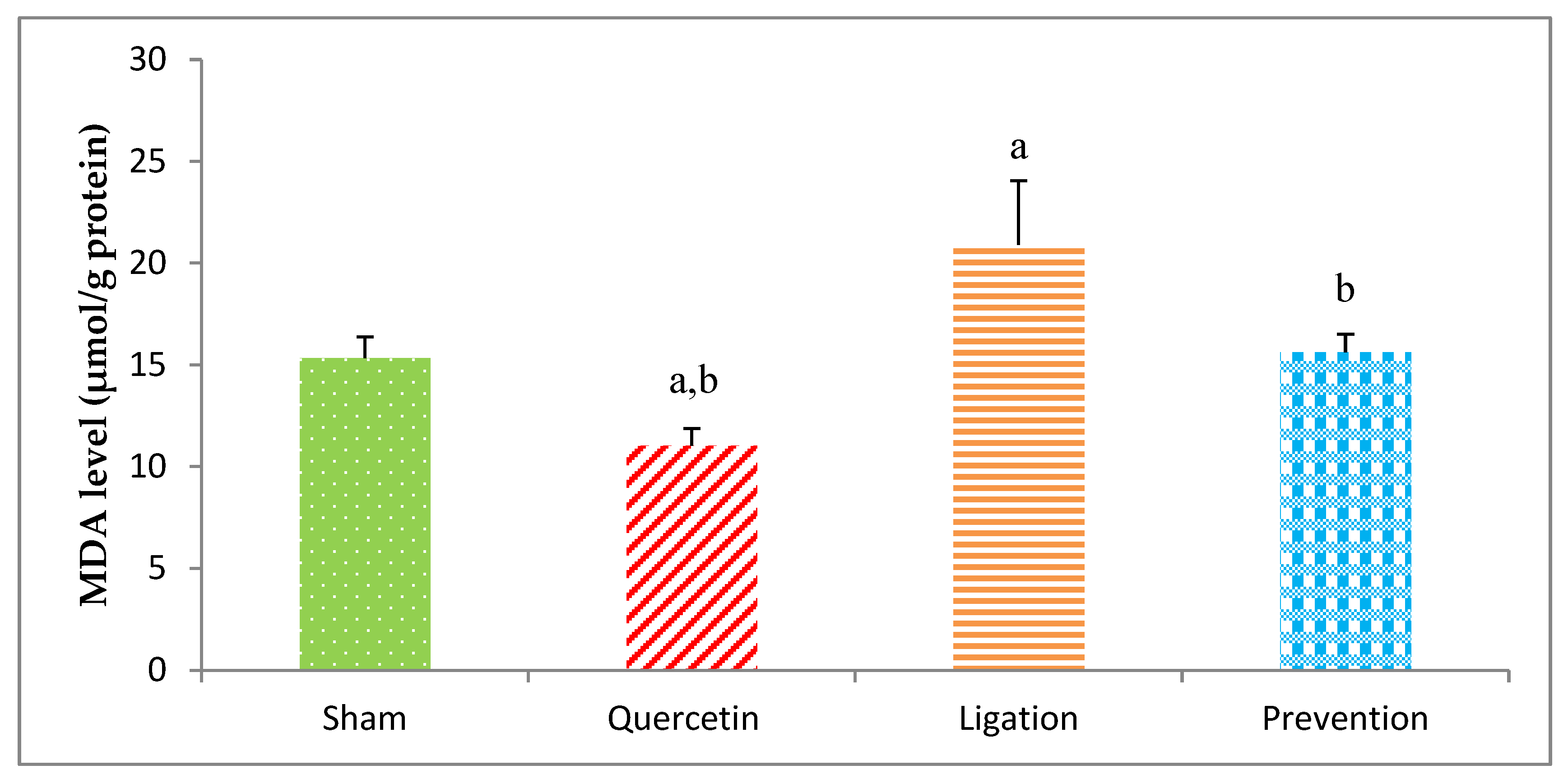
2.1. Malondialdehyde (MDA) Levels in the Ipsilateral Brain Cortex Homogenates
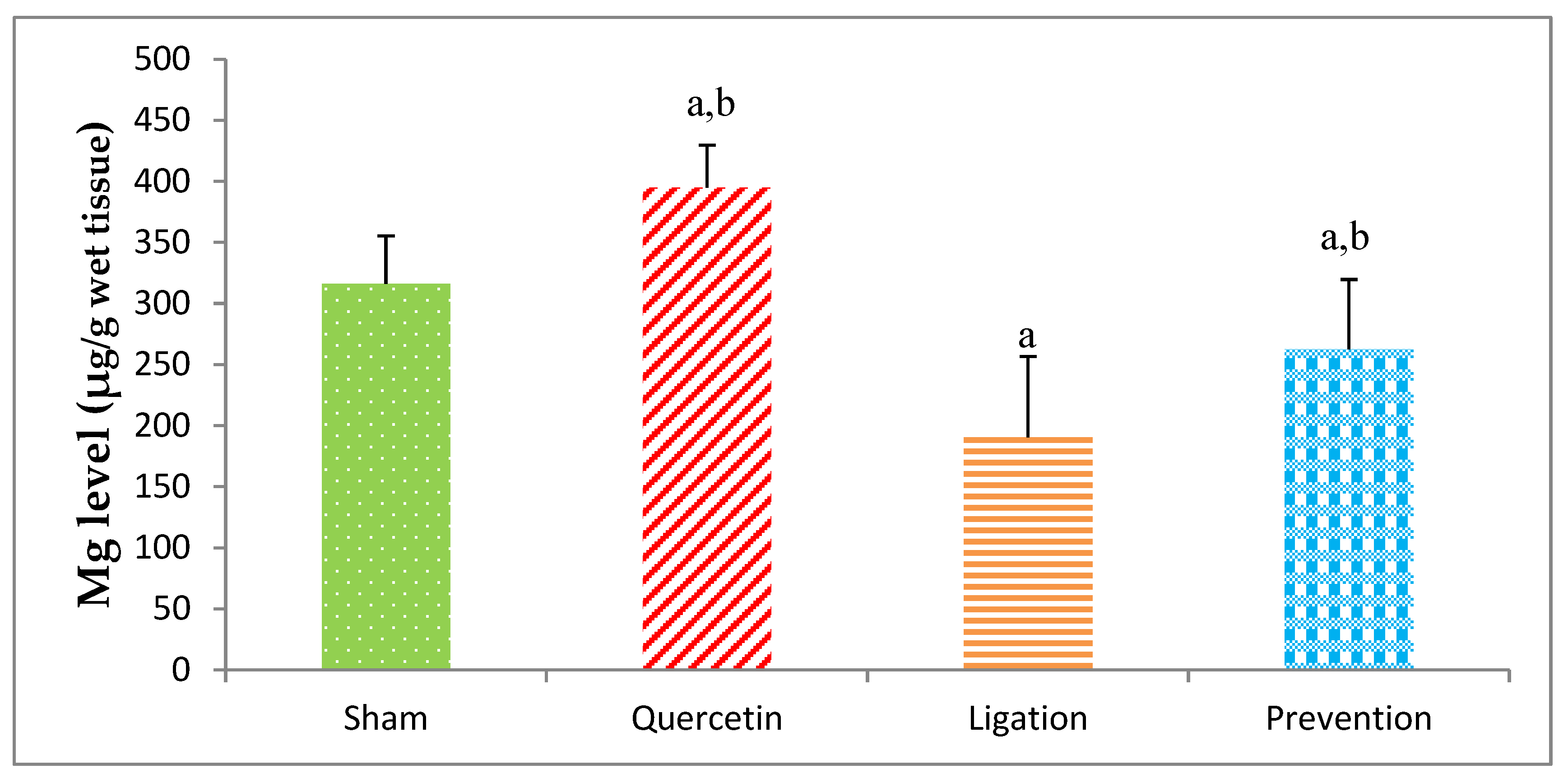
2.1.1. Essential Trace Element Mg Levels in the Ipsilateral Brain Cortex Homogenates
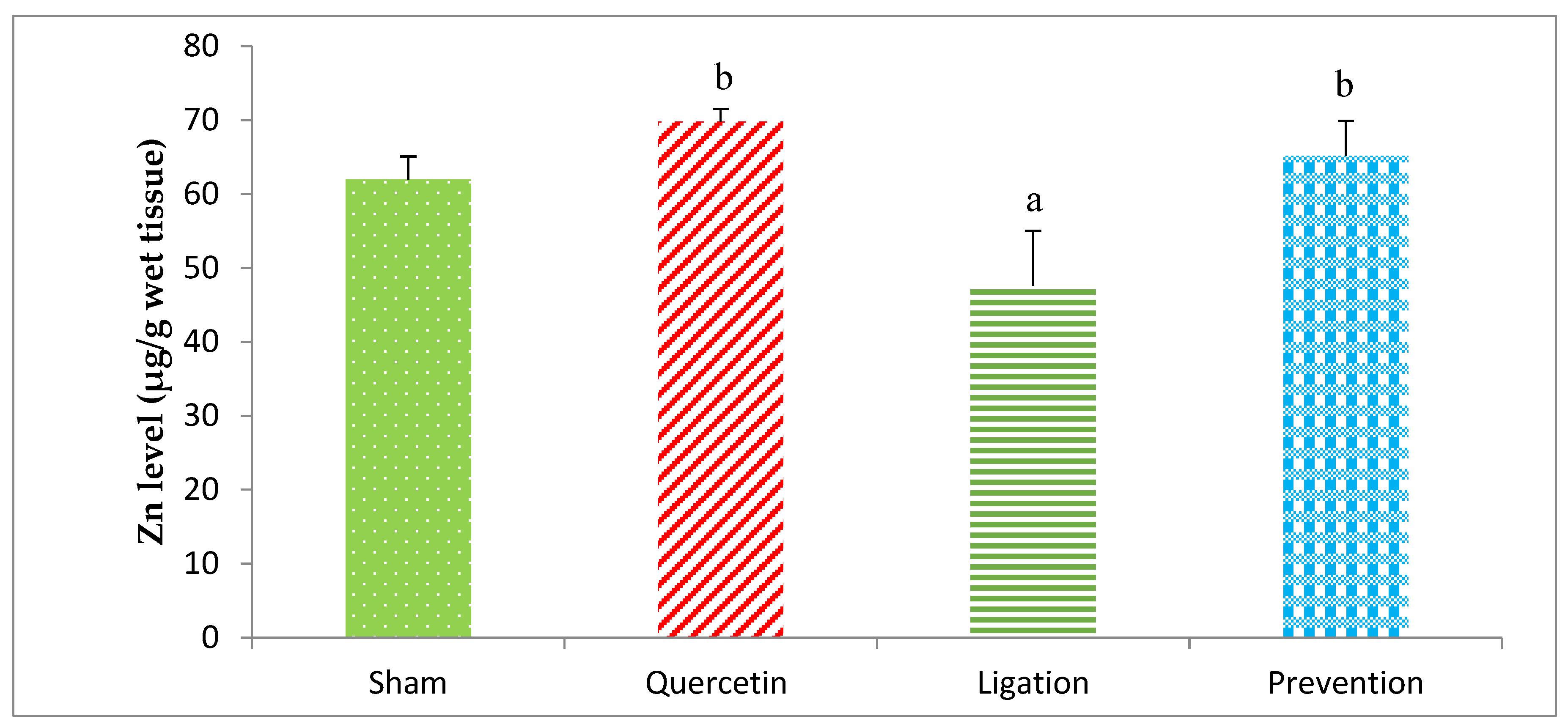
2.1.2. Essential Trace Element Zn Levels in the Ipsilateral Brain Cortex Homogenates
2.1.3. Essential Trace Element Se Levels in the Ipsilateral Brain Cortex Homogenates
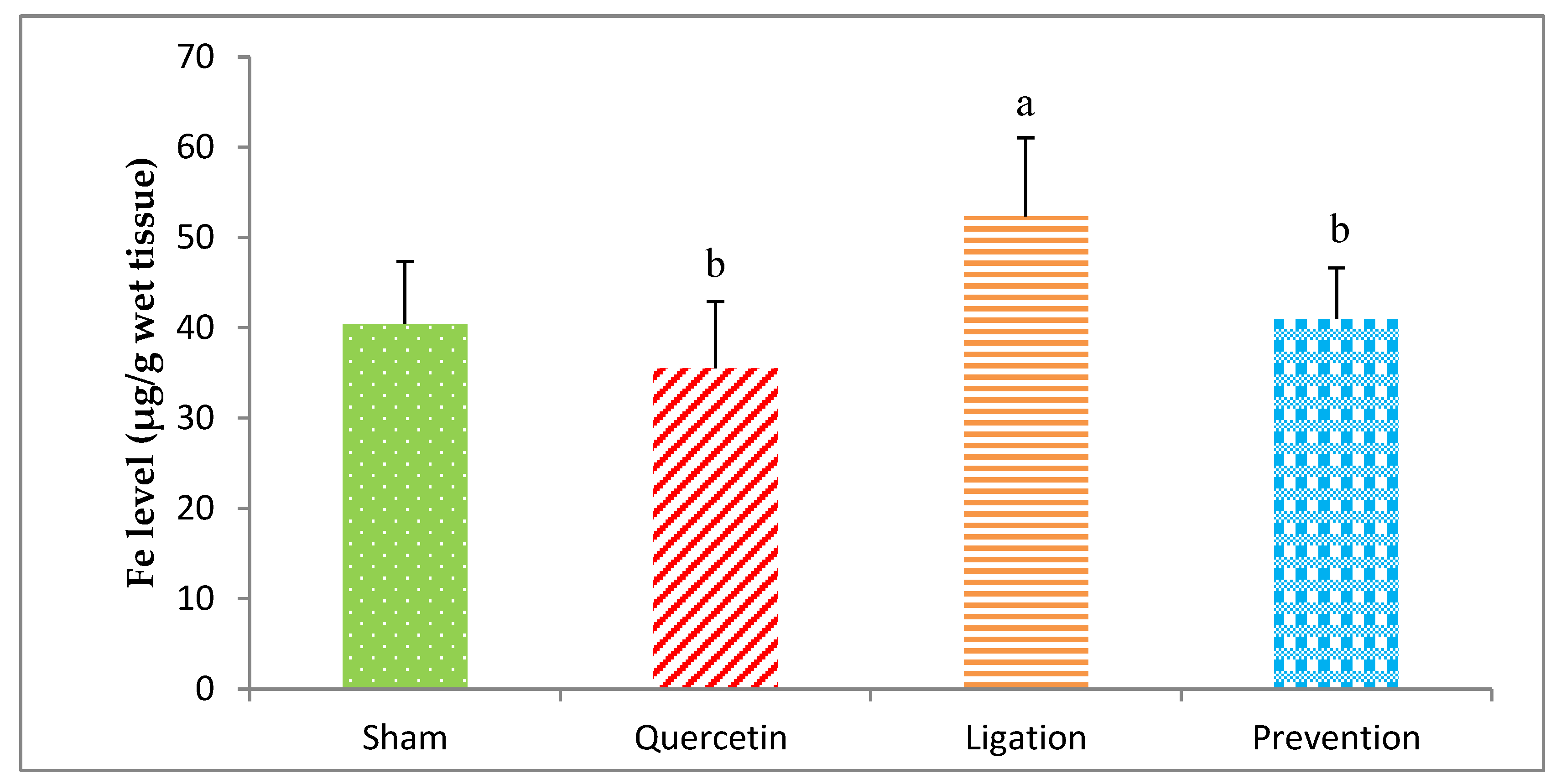
2.1.4. Essential Trace Element Fe Levels in the Ipsilateral Brain Cortex Homogenates
2.1.5. Essential Trace Element Cu Levels in the Ipsilateral Brain Cortex Homogenates
2.2. Antioxidant Activity in the Ipsilateral Brain Cortex Homogenates
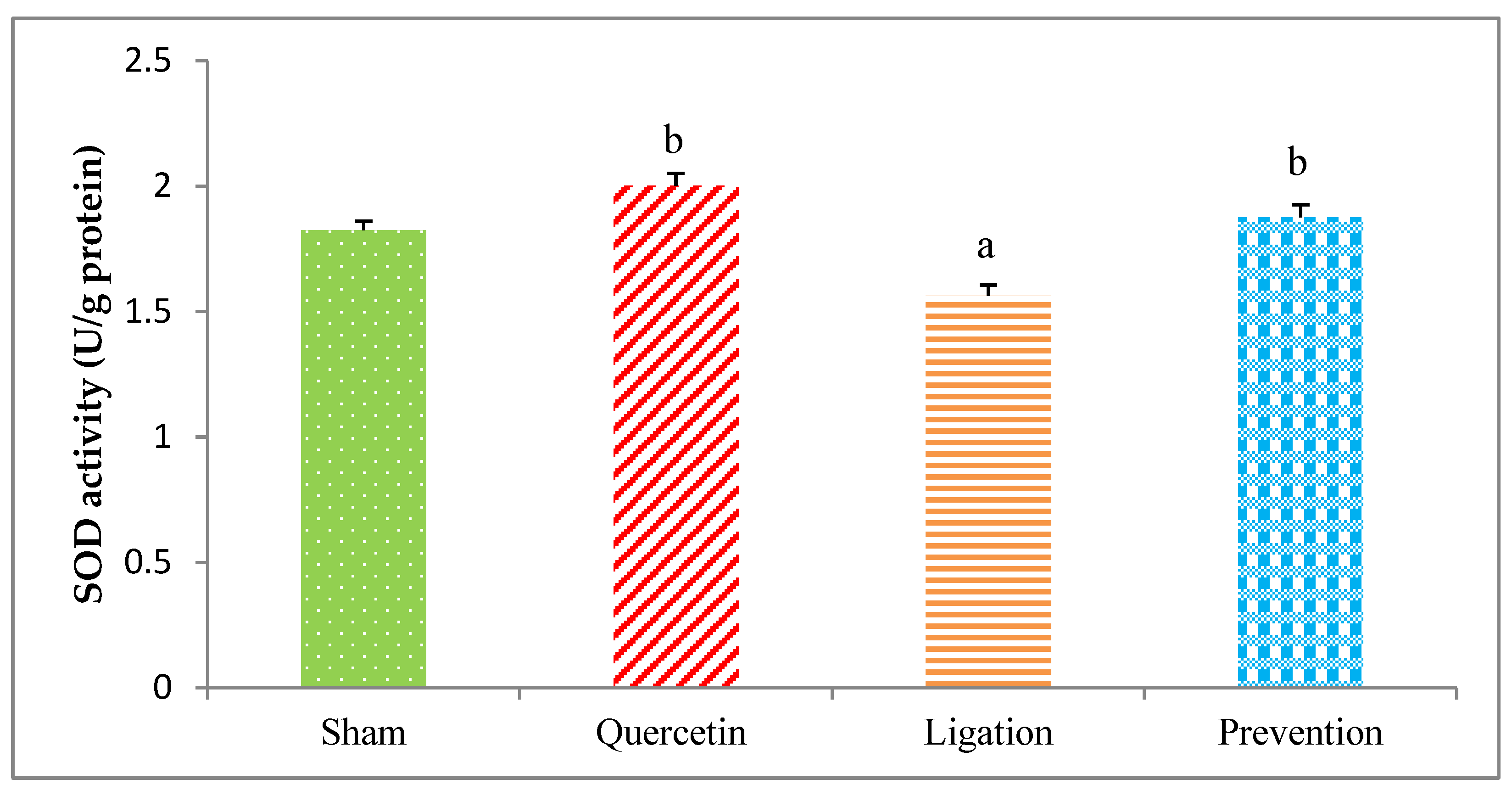
2.2.1. The SOD Activity in the Ipsilateral Brain Cortex Homogenates
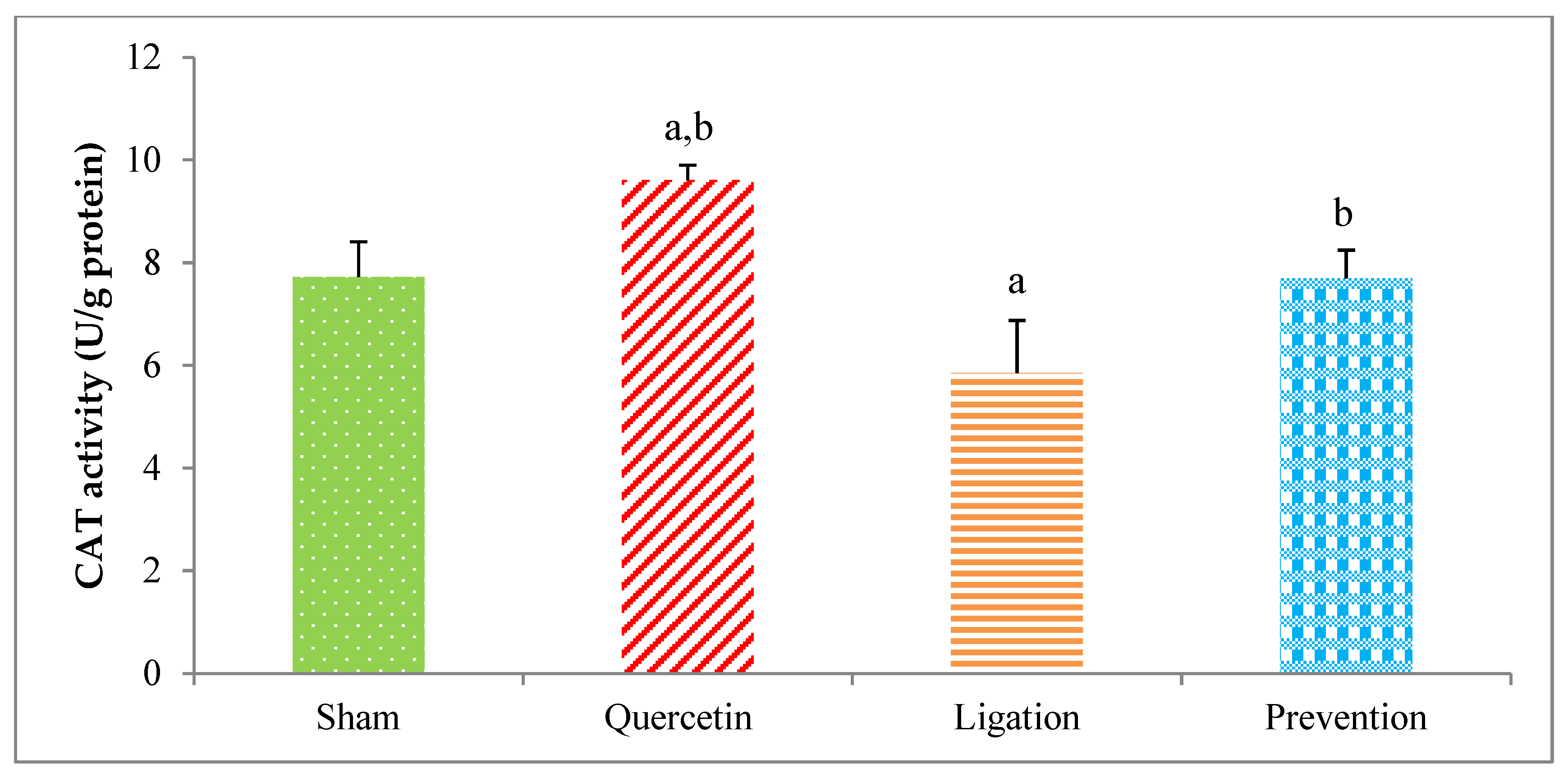
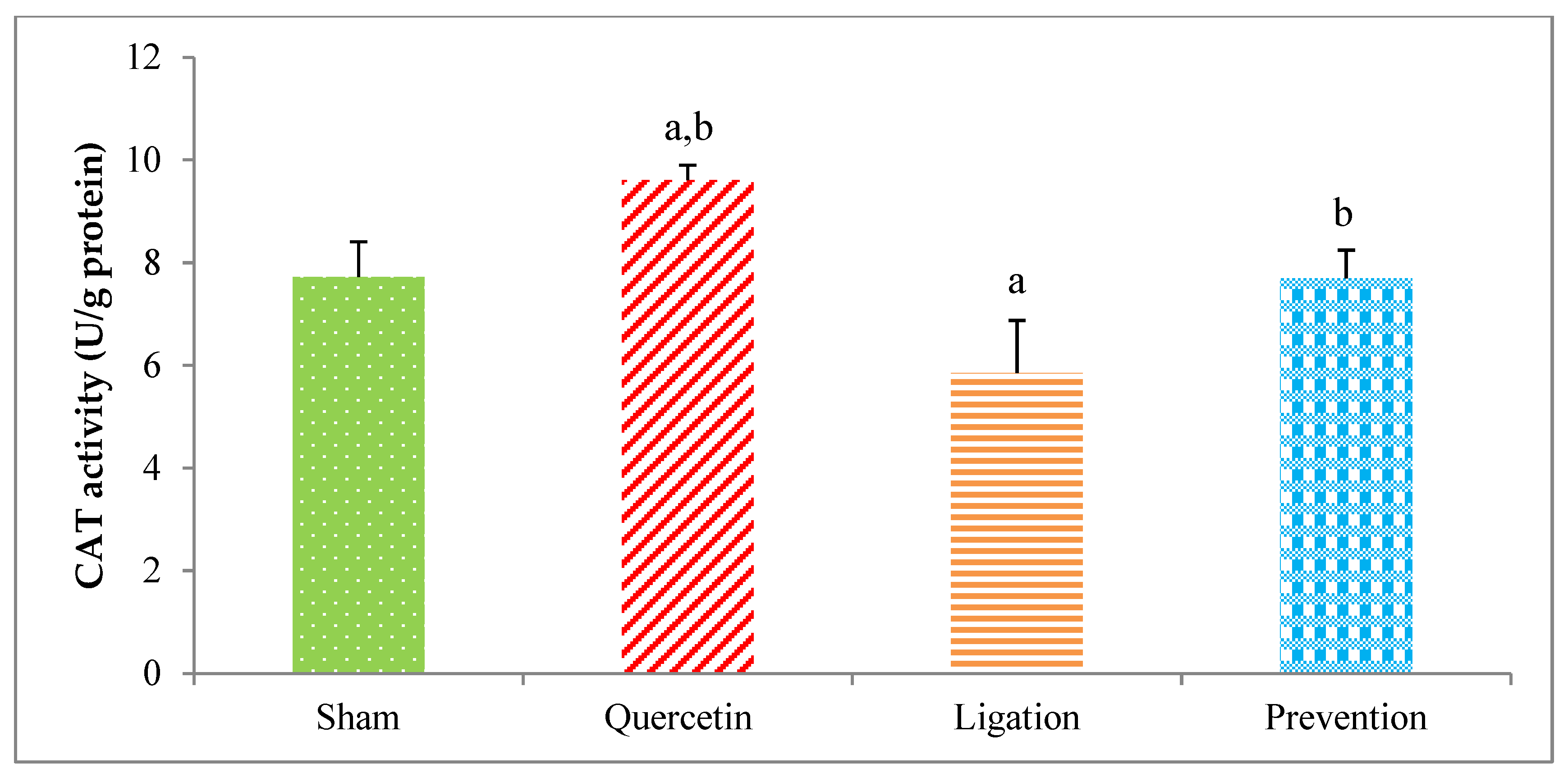
2.2.2. The CAT Activity in the Ipsilateral Brain Cortex Homogenates
2.3. The Cu/Zn Ratio in the Ipsilateral Brain Cortex Homogenates
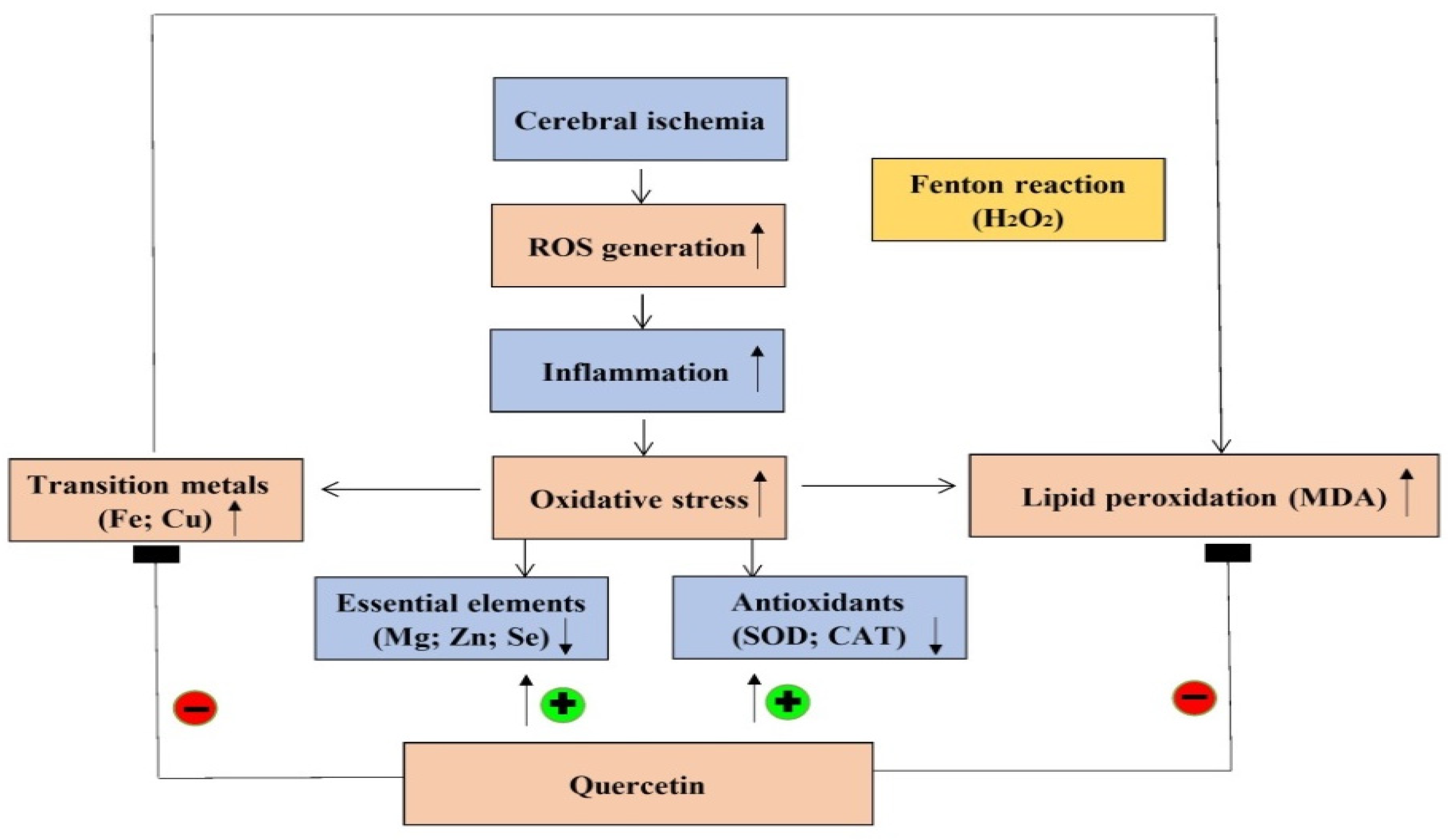
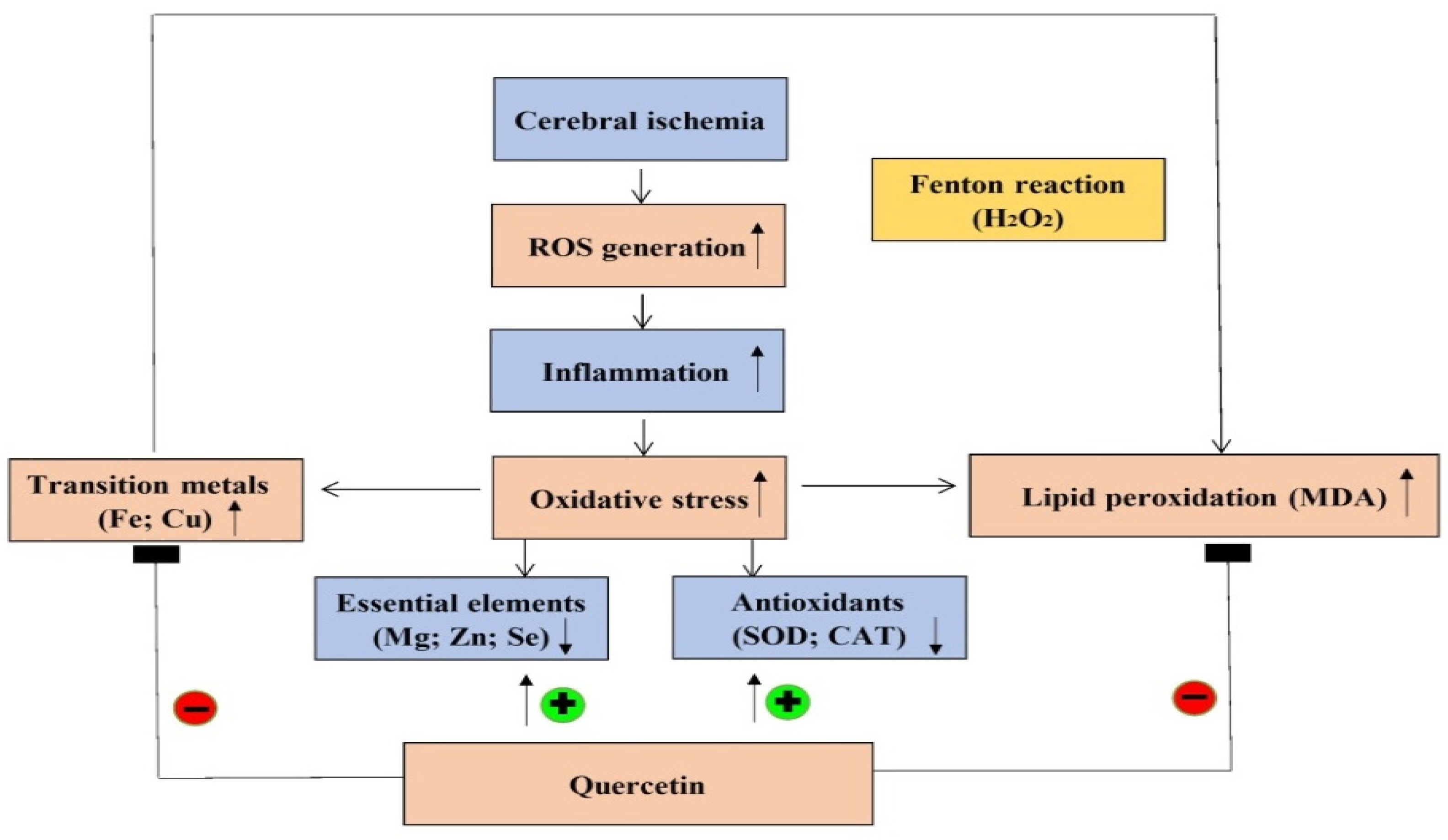
3. Discussion
4. Materials and Methods
4.1. Animal Treatment and Samples Preparation
4.2. Malondialdehyde (MDA) Concentration Analysis in the Ipsilateral Brain Cortex Homogenates
4.3. Analysis of Enzyme Activity in the Ipsilateral Brain Cortex Homogenates
4.4. Measurement of Essential Trace Element in the Ipsilateral Brain Cortex Homogenates
4.5. Protein Concentration Analysis in the Ipsilateral Brain Cortex Homogenates
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Ro, J.H.; Liu, C.C.; Lin, M.C. Resveratrol mitigates cerebral ischemic injury by altering levels of trace elements, toxic metal, lipid peroxidation, and antioxidant activity. Biol. Trace Elem. Res. 2021, 199, 3718–3727. [Google Scholar] [CrossRef]
- Fang, K.M.; Cheng, F.C.; Huang, Y.L.; Chung, S.Y.; Jian, Z.Y.; Lin, M.C. Trace element, antioxidant activity, and lipid peroxidation levels in brain cortex of gerbils after cerebral ischemic injury. Biol. Trace Elem. Res. 2013, 152, 66–74. [Google Scholar] [CrossRef]
- Uthra, C.; Shrivastava, S.; Jaswal, A.; Sinha, N.; Reshi, M.S.; Shukla, S. Therapeutic potential of quercetin against acrylamide induced toxicity in rats. Biomed. Pharmacother. 2017, 86, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Liu, M.; He, Y.; Yang, B. Quercetin protect cigarette smoke extracts induced inflammation and apoptosis in RPE cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2010–2015. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications. Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef] [PubMed]
- de Baaij, J.H.F.; Hoenderop, J.G.J.; Bindels, R.J.M. Regulation of magnesium balance: Lessons learned from human genetic disease. Clin. Kidney J. 2012, 5, i15–i24. [Google Scholar] [CrossRef]
- Kwon, W.Y.; Suh, G.J.; Kim, K.S.; Jung, Y.S.; Kim, S.H.; Lee, A.R.; You, K.M.; Park, M.J. Niacin and selenium attenuate brain injury after cardiac arrest in rats by up-regulating DJ-1-Akt signaling. Crit. Care Med. 2018, 46, e788–e796. [Google Scholar] [CrossRef]
- Allen, K.J.; Gurrin, L.C.; Constantine, C.C.; Osborne, N.J.; Delatycki, M.B.; Nicoll, A.J.; McLaren, C.E.; Bahlo, M.; Nisselle, A.E.; Vulpe, C.D.; et al. Iron-overload-related disease in HFE hereditary hemochromatosis. N. Engl. J. Med. 2008, 358, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Murphy, C.J.; Oudit, G.Y. Iron-overload cardiomyopathy: Pathophysiology, diagnosis, and treatment. J. Card. Fail. 2010, 16, 888–900. [Google Scholar] [CrossRef]
- Matos, L.; Gouveia, A.M.; Almeida, H. Resveratrol attenuates copper-induced senescence by lmproving cellular proteostasis. Oxid Med. Cell. Longev. 2017, 2017, 3793817. [Google Scholar]
- Boilan, E.; Winant, V.; Dumortier, E.; Piret, J.P.; Bonfitto, F.; Osiewacz, H.D.; Debacq-Chainiaux, F.; Toussaint, O. Role of p38MAPK and oxidative stress in copper-induced enescence. Age 2013, 35, 2255–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyama, O.; Matsuyama, T.; Michishita, H.; Nakamura, H.; Sugita, M. Protective effects of human recombinant superoxide dismutase on transient ischemic injury of CA1 neurons in gerbils. Stroke 1992, 23, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Yuan, X.; Liu, T.; Liu, L.; Hu, Y.; Wang, Z.; Zheng, Q. Neuroprotective activity of lavender oil on transient focal cerebral ischemia in mice. Molecules 2012, 17, 9803–9817. [Google Scholar] [CrossRef] [PubMed]
- Mirończuk, A.; Kapica-Topczewska, K.; Socha, K.; Soroczyńska, J.; Jamiołkowski, J.; Kułakowska, A.; Kochanowicz, J. Selenium, Copper, Zinc Concentrations and Cu/Zn, Cu/Se Molar Ratios in the Serum of Patients with Acute Ischemic Stroke in Northeastern Poland—A New Insight into Stroke Pathophysiology. Nutrients 2021, 13, 2139. [Google Scholar] [CrossRef] [PubMed]
- Wegner, K.; Saleh, D.; Degterev, A. Complex pathologic roles of RIPK1 and RIPK3: Moving beyond necroptosis. Trends Pharmacol. Sci. 2017, 38, 202–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Q.; Yin, H.; Min, Y.; Li, S. Selenium deficiency causes immune damage by activating the DUSP1/NF-κB pathway and endoplasmic reticulum stress in chicken spleen. Food Funct. 2020, 11, 6467–6475. [Google Scholar]
- Li, X.; Xing, M.; Chen, M.; Zhao, J.; Fan, R.; Zhao, X.; Cao, C.; Yang, J.; Zhang, Z.; Xu, S. Effects of selenium-lead interaction on the gene expression of inflammatory factors and selenoproteins in chicken neutrophils. Ecotoxicol. Environ. Saf. 2017, 139, 447–453. [Google Scholar] [CrossRef]
- Yang, H.; Song, Y.; Liang, Y.; Li, R. Quercetin treatment improves renal function and protects the kidney in a rat model of adenine-induced chronic kidney disease. Med. Sci. Monit. 2018, 24, 4760–4766. [Google Scholar] [CrossRef]
- Gupta, R.; Shukla, R.; Chandravanshi, L.; Srivastava, P.; Dhuriya, Y.; Shanker, J.; Singh, M.; Pant, A.; Khanna, V. Protective role of quercetin in cadmium-induced cholinergic dysfunctions in rat brain by modulating mitochondrial integrity and MAP kinase signaling. Mol. Neurobiol. 2017, 54, 4560–4583. [Google Scholar] [CrossRef]
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The Role of Magnesium in Neurological Disorders. Nutrients 2018, 10, 730. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.Y.; Chung, S.Y.; Lin, M.C.; Cheng, F.C. Effects of magnesium sulfate on energy metabolites and glutamate in the cortex during focal cerebral ischemia and reperfusion in the gerbil monitored by a dual-probe microdialysis technique. Life Sci. 2002, 71, 803–811. [Google Scholar] [CrossRef]
- Guerrero-Romero, F.; Rodríguez-Morán, M. Hypomagnesemia, oxidative stress, inflammation, and metabolic syndrome. Diabetes Metab Res. Rev. 2006, 22, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Xu, L.; Huang, Y.; Chen, T.; Zhou, T.; Yang, L. Magnesium sulfate can alleviate oxidative stress and reduce inflammatory cytokines in rat placenta of intrahepatic cholestasis of pregnancy model. Arch. Gynecol. Obs. 2018, 298, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Siegler, J.E.; Boehme, A.K.; Albright, K.C.; Bdeir, S.; Kar, A.K.; Myers, L.; Beasley, T.M.; Martin-Schild, S. Acute decrease in serum magnesium level after ischemic stroke may not predict decrease in neurologic function. J. Stroke Cerebrovasc. Dis. 2013, 22, e516–e521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malyar, R.M.; Li, H.; Liu, D.; Abdulrahim, Y.; Farid, R.A.; Gan, F.; Ali, W.; Enayatullah, H.; Banuree, S.A.H.; Huang, K.; et al. Selenium/zinc-enriched probiotics improve serum enzyme activity, antioxidant ability, inflammatory factors and related gene expression of wistar rats inflated under heat stress. Life Sci. 2020, 248, 117464. [Google Scholar] [CrossRef]
- Yang, D.Y.; Lee, J.B.; Lin, M.C.; Huang, Y.L.; Liu, H.W.; Liang, Y.J.; Cheng, F.C. The determination of brain magnesium and zinc levels by a dual-probe microdialysis and graphite furnace atomic absorption spectrometry. J. Am. Coll. Nutr. 2004, 23, 552S–555S. [Google Scholar] [CrossRef]
- Li, B.; Cui, W.; Tan, Y.; Luo, P.; Chen, Q.; Zhang, C.; Qu, W.; Miao, L.; Cai, L. Zinc is essential for the transcription function of Nrf2 in human renal tubule cells in vitro and mouse kidney in vivo under the diabetic condition. J. Cell. Mol. Med. 2014, 18, 895–906. [Google Scholar] [CrossRef]
- Schweizer, U.; Brauer, U.; Kohrle, N.R.; Savaskan, N.E. Selenium and brain function: A poorly recognized liaison. Brain Res. 2004, 45, 164–178. [Google Scholar] [CrossRef]
- Medjati, N.D.; Harek, Y.; Tarik, A.; Lahcene, L. Whole blood selenium levels in healthy adults from the west of Algeria. Biol. Trace Elem. Res. 2012, 147, 44–48. [Google Scholar] [CrossRef]
- Ognjanović, B.I.; Djordjević, N.Z.; Matić, M.M.; Obradović, J.M.; Mladenović, J.M.; Stajn, A.Š.; Saičić, Z.S. Lipid peroxidative damage on Cisplatin exposure and alterations in antioxidant defense system in rat kidneys: A possible protective effect of selenium. Int. J. Mol. Sci. 2012, 13, 1790–1803. [Google Scholar] [CrossRef] [Green Version]
- Song, E.; Su, C.; Fu, J.; Xia, X.; Yang, S.; Xiao, C.; Lu, B.; Chen, H.; Sun, Z.; Wu, S.; et al. Selenium supplementation shows protective effects against patulin-induced brain damage in mice via increases in GSH-related enzyme activity and expression. Life Sci. 2014, 109, 37–43. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.H.; Nam, S.W.; Choi, Y.H. Gastroprotective effect of selenium on ethanol-induced gastric damage in rats. Int. J. Mol. Sci. 2012, 13, 5740–5750. [Google Scholar] [CrossRef] [Green Version]
- van Eersel, J.; Ke, Y.D.; Liu, X.; Delerue, F.; Kril, J.J.; Gotz, J.; Ittner, L.M. Sodium selenate mitigates tau pathology, neurodegeneration, and functional deficits in Alzheimer’s disease models. Proc. Natl. Acad. Sci. USA 2010, 107, 13888–13893. [Google Scholar] [CrossRef] [Green Version]
- Cosín-Tomàs, M.; Senserrich, J.; Arumí-Planas, M.; Alquézar, C.; Pallàs, M.; Martín-Requero, Á.; Suñol, C.; Kaliman, P.; Sanfeliu, C. Role of resveratrol and selenium on oxidative stress and expression of antioxidant and anti-aging genes in immortalized lymphocytes from Alzheimer’s disease patients. Nutrients 2019, 11, 1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Dávalos, A.; Fernandez-Real, J.M.; Ricart, W.; Soler, S.; Molins, A.; Planas, E.; Genís, D. Iron-related damage in acute ischemic stroke. Stroke 1994, 25, 1543–1546. [Google Scholar] [CrossRef] [Green Version]
- Barbouti, A.; Doulias, P.T.; Zhu, B.Z.; Frei, B.; Galaris, D. Intracellular iron, but not copper, plays a critical role in hydrogen peroxide induced DNA damage. Free Radic. Biol. Med. 2001, 31, 490–498. [Google Scholar] [CrossRef]
- Meneghini, R. Iron homeostasis, oxidative stress, and DNA damage. Free Radic. Biol Med. 1997, 23, 783–792. [Google Scholar] [CrossRef]
- Catapano, M.C.; Tvrdý, V.; Karlíčková, J.; Migkos, T.; Valentová, K.; Křen, V.; Mladěnka, P. The Stoichiometry of Isoquercitrin Complex with Iron or Copper Is Highly Dependent on Experimental Conditions. Nutrients 2017, 9, 1193. [Google Scholar] [CrossRef] [Green Version]
- Hasumura, M.; Yasuhara, K.; Tamura, T.; Imai, T.; Mitsumori, K.; Hirose, M. Evaluation of the toxicity of enzymatically decomposed rutin with 13-weeks dietary administration to Wistar rats. Food Chem. Toxicol. 2004, 42, 439–444. [Google Scholar] [CrossRef]
- Brune, M.; Rossander, L.; Hallberg, L. Iron absorption and phenolic compounds: Importance of different phenolic structures. Eur. J. Clin. Nutr. 1989, 43, 547–557. [Google Scholar]
- Bost, M.; Houdart, S.; Oberli, M.; Kalonji, E.; Huneau, J.F.; Margaritis, I. Dietary copper and human health: Current evidence and unresolved issues. J. Trace Elem. Med. Biol. 2016, 35, 107–115. [Google Scholar] [CrossRef]
- Xiao, Y.; Yuan, Y.; Liu, Y.; Yu, Y.; Jia, N.; Zhou, L.; Wang, H.; Huang, S.; Zhang, Y.; Yang, H.; et al. Circulating multiple metals and incident stroke in Chinese adults. Stroke 2019, 50, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Zubčić, K.; Radovanović, V.; Vlainić, J.; Hof, P.R.; Oršolić, N.; Šimić, G.; Jazvinšćak Jembrek, M. PI3K/Akt and ERK1/2 signalling are involved in quercetin-mediated neuroprotection against copper-induced injury. Oxid Med. Cell. Longev. 2020, 2020, 9834742. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Huang, S.; Zhang, Y.; Zhang, H.; Zhou, L.; Li, D.; Cheng, J. Associations of multiple plasma metals with the risk of ischemic stroke: A case-control study. Environ. Int. 2019, 125, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, W.; Wang, Y.; Wang, T.; Ma, M.; Tian, C. Association between the change of serum copper and ischemic stroke: A systematic review and meta-analysis. J. Mol. Neurosci. 2020, 70, 475–480. [Google Scholar] [CrossRef]
- Skalny, A.V.; Klimenko, L.L.; Turna, A.A.; Budanova, M.N.; Baskakov, I.S.; Savostina, M.S.; Mazilina, A.N.; Deyev, A.I.; Skalnaya, M.G.; Tinkov, A.A. Serum trace elements are associated with hemostasis, lipid spectrum and inflammatory markers in men suffering from acute ischemic stroke. Metab. Brain Dis. 2017, 32, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Lee, Y.D.; Wang, H.L.; Liao, K.H.; Chen, K.B.; Poon, K.S.; Pan, Y.L.; Lai, T.W. Anesthesia-induced hypothermia attenuates early-phase blood-brain barrier disruption but not infarct volume following cerebral ischemia. PLoS ONE 2017, 12, e0170682. [Google Scholar] [CrossRef]
- Yao, R.Q.; Qi, D.S.; Yu, H.L.; Liu, J.; Yang, L.H.; Wu, X.X. Quercetin attenuates cell apoptosis in focal cerebral ischemia rat brain via activation of BDNF–TrkB–PI3K/Akt signaling pathway. Neurochem. Res. 2012, 37, 2777–2786. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, M.-C.; Liu, C.-C.; Liao, C.-S.; Ro, J.-H. Neuroprotective Effect of Quercetin during Cerebral Ischemic Injury Involves Regulation of Essential Elements, Transition Metals, Cu/Zn Ratio, and Antioxidant Activity. Molecules 2021, 26, 6128. https://doi.org/10.3390/molecules26206128
Lin M-C, Liu C-C, Liao C-S, Ro J-H. Neuroprotective Effect of Quercetin during Cerebral Ischemic Injury Involves Regulation of Essential Elements, Transition Metals, Cu/Zn Ratio, and Antioxidant Activity. Molecules. 2021; 26(20):6128. https://doi.org/10.3390/molecules26206128
Chicago/Turabian StyleLin, Ming-Cheng, Chien-Chi Liu, Chin-Sheng Liao, and Ju-Hai Ro. 2021. "Neuroprotective Effect of Quercetin during Cerebral Ischemic Injury Involves Regulation of Essential Elements, Transition Metals, Cu/Zn Ratio, and Antioxidant Activity" Molecules 26, no. 20: 6128. https://doi.org/10.3390/molecules26206128
APA StyleLin, M.-C., Liu, C.-C., Liao, C.-S., & Ro, J.-H. (2021). Neuroprotective Effect of Quercetin during Cerebral Ischemic Injury Involves Regulation of Essential Elements, Transition Metals, Cu/Zn Ratio, and Antioxidant Activity. Molecules, 26(20), 6128. https://doi.org/10.3390/molecules26206128