
An Overview of Peptide-Based Molecules as Potential Drug Candidates for Multiple Sclerosis




Abstract
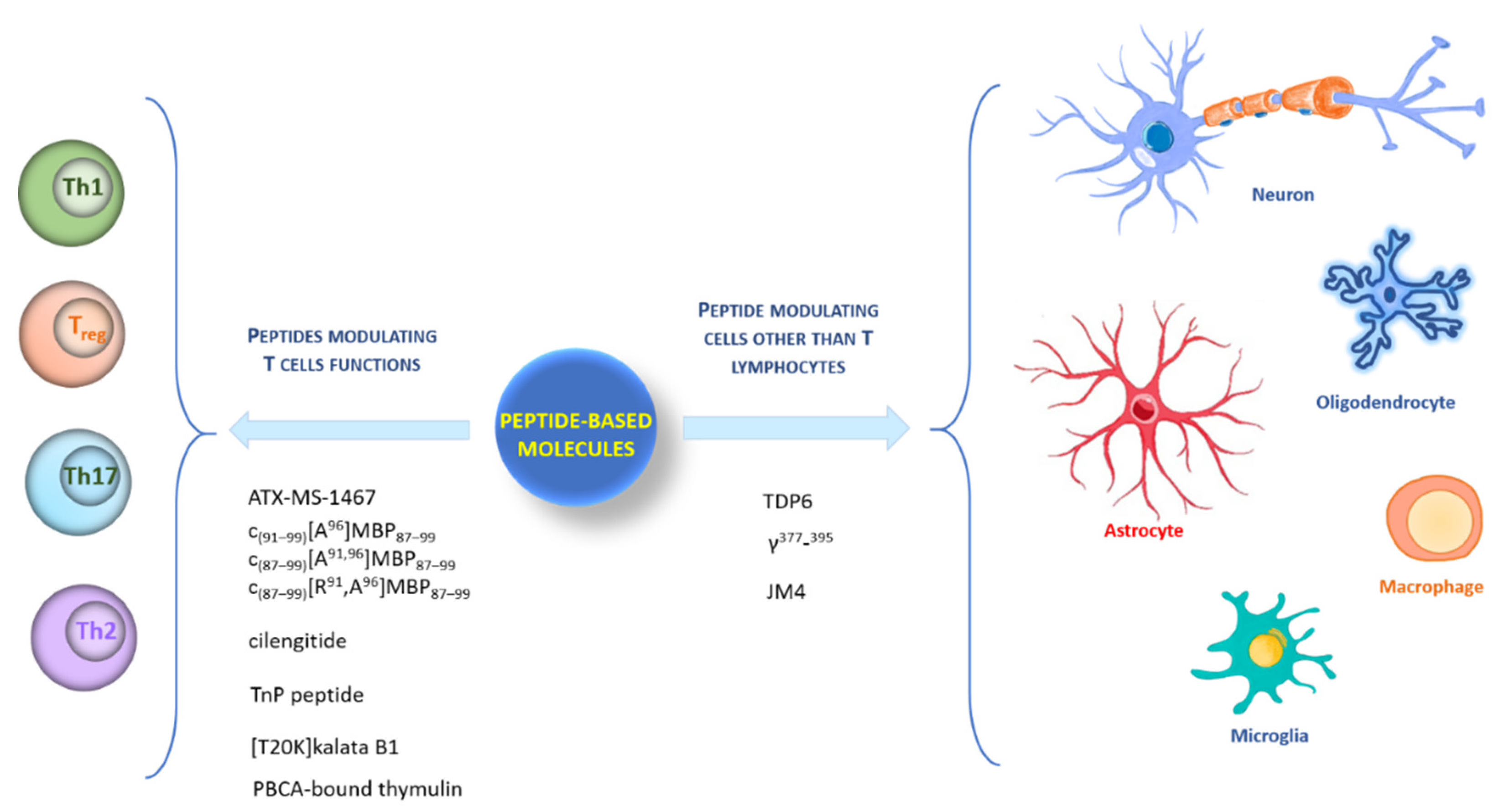
1. Introduction
1.1. Immunopathogenesis of MS
1.2. Current Available Therapeutic Approaches
2. Peptides Modulating T Cells’ Functions
2.1. Myelin-Based Autoantigen Peptides: ATX-MS-1467, c(91–99)[A96]MBP87–99, c(87–99)[A91,96]MBP87–99, and c(87–99)[ R91, A96]MBP87–99
2.2. Peptides Modulating Th Cell Subsets: Cilengitide, TnP Peptide, [T20K] Kalata B1, Thymulin
3. Peptides Modulating Other Cells besides T Lymphocytes
3.1. Remyelination-Enhancing Peptide: TDP6
3.2. Peptide Inhibiting Microglia Activation: γ377–395
3.3. Peptide Regulating T Cell Subsets and CNS Resident Cells: JM4

{kind=link}
| Name | Sequence | Ref |
|---|---|---|
| ATX-MS-1467 | ATX-MS1: PRHRDTGILDSIGRF | [34] |
| [ATX-MS4]: GFKGVDAQGTLSKIF | ||
| [ATX-MS6]: GFKGVDAQGTLSKIF | ||
| [ATX-MS7]: ENPVVHFFKNIVTPRTP | ||
| cyclo (91–99) [Ala96] MBP87–99 |  | [44] |
| cyclo (87–99) [Ala91,96] MBP87–99 |  | |
| cyclo (87–99) [Arg91,Ala96] MBP87–99 |  | |
| Cilengitide | c(RGDf(NMe)V) | [56] |
| TnP peptide |  | [59] |
| [T20K]kalata B1 |  | [65] |
| Thymulin | EAKSQGGSD | [74,75] |
| TDP6 |  | [77] |
| γ377-395 | YSMKKTTMKIIPFNRLTIG | [79] |
| JM4 | GCAEHCSLNENITVPDTKV | [88] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ransohoff, R.M. Immune-cell crosstalk in multiple sclerosis. Nature 2018, 563, 194–195. [Google Scholar] [CrossRef] [PubMed]
- Willekens, B.; Cools, N. Beyond the Magic Bullet: Current Progress of Therapeutic Vaccination in Multiple Sclerosis. CNS Drugs 2018, 32, 401–410. [Google Scholar] [CrossRef]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545–558. [Google Scholar] [CrossRef]
- Grigoriadis, N.; van Pesch, V. A basic overview of multiple sclerosis immunopathology. Eur. J. neurol. 2015, 22, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Nuyts, A.H.; Lee, W.P.; Bashir-Dar, R.; Berneman, Z.N.; Cools, N. Dendritic cells in multiple sclerosis: Key players in the immunopathogenesis, key players for new cellular immunotherapies? Mult. Scler. 2013, 19, 995–1002. [Google Scholar] [CrossRef]
- McFarland, H.F.; Martin, R. Multiple sclerosis: A complicated picture of autoimmunity. Nat. Immunol. 2007, 8, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Baldassari, L.E.; Feng, J.; Clayton, B.L.L.; Oh, S.H.; Sakaie, K.; Tesar, P.J.; Wang, Y.; Cohen, J.A. Developing therapeutic strategies to promote myelin repair in multiple sclerosis. Expert Rev. Neurother. 2019, 19, 997–1013. [Google Scholar] [CrossRef]
- Gholamzad, M.; Ebtekar, M.; Ardestani, M.S.; Azimi, M.; Mahmodi, Z.; Mousavi, M.J.; Aslani, S. A comprehensive review on the treatment approaches of multiple sclerosis: Currently and in the future. Inflamm. Res. 2019, 68, 25–38. [Google Scholar] [CrossRef]
- Compston, A. The genetic epidemiology of multiple sclerosis. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1999, 354, 1623–1634. [Google Scholar] [CrossRef]
- Lincoln, M.R.; Montpetit, A.; Cader, M.Z.; Saarela, J.; Dyment, D.A.; Tiislar, M.; Ferretti, V.; Tienari, P.J.; Sadovnick, A.D.; Peltonen, L.; et al. Hudson, A predominant role for the HLA class II region in the association of the MHC region with multiple sclerosis. Nat. Genet. 2005, 37, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef] [PubMed]
- Fujinami, R.S.; von Herrath, M.G.; Christen, U.; Whitton, J.L. Molecular mimicry, bystander activation, or viral persistence: Infections and autoimmune disease. Clin. Microbiol. Rev. 2006, 19, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Gilden, D.H. Infectious causes of multiple sclerosis. Lancet. Neurol. 2005, 4, 195–202. [Google Scholar] [CrossRef]
- Pender, M.P. Does Epstein-Barr virus infection in the brain drive the development of multiple sclerosis? Brain 2009, 132, 3196–3198. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bagert, B.A. Epstein-Barr virus in multiple sclerosis. Curr. Neurol. Neurosci. Rep. 2009, 9, 405–410. [Google Scholar] [CrossRef]
- Gaydos, C.A. Chlamydia pneumoniae and its proposed link to multiple sclerosis: To be or not to be? Neurology 2001, 56, 1126–1127. [Google Scholar] [CrossRef]
- Jacobson, S.; Cross, A. Association of Chlamydia pneumoniae and multiple sclerosis: Stage two? Neurology 2001, 56, 1128–1129. [Google Scholar] [CrossRef]
- Yao, S.Y.; Ljunggren-Rose, A.; Stratton, C.W.; Mitchell, W.M.; Sriram, S. Regulation by IFN-beta of inducible nitric oxide synthase and interleukin-12/p40 in murine macrophages cultured in the presence of Chlamydia pneumoniae antigens. J. Interferon. Cytokine Res. 2001, 21, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kenealy, S.J.; Pericak-Vance, M.A.; Haines, J.L. The genetic epidemiology of multiple sclerosis. J. Neuroimmunol. 2003, 143, 7–12. [Google Scholar] [CrossRef]
- Hafler, D.A.; Benjamin, D.S.; Burks, J.; Weiner, H.L. Myelin basic protein and proteolipid protein reactivity of brain- and cerebrospinal fluid-derived T cell clones in multiple sclerosis and postinfectious encephalomyelitis. J. Immunol. 1987, 139, 68–72. [Google Scholar]
- Ota, K.; Matsui, M.; Milford, E.L.; Mackin, G.A.; Weiner, H.L.; Hafler, D.A. T-cell recognition of an immunodominant myelin basic protein epitope in multiple sclerosis. Nature 1990, 346, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Zamvil, S.; Nelson, P.; Trotter, J.; Mitchell, D.; Knobler, R.; Fritz, R.; Steinman, L. T-cell clones specific for myelin basic protein induce chronic relapsing paralysis and demyelination. Nature 1985, 317, 355–358. [Google Scholar] [CrossRef]
- Zamvil, S.S.; Nelson, P.A.; Mitchell, D.J.; Knobler, R.L.; Fritz, R.B.; Steinman, L. Encephalitogenic T cell clones specific for myelin basic protein. An unusual bias in antigen recognition. J. Exp. Med. 1985, 162, 2107–2124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Markovic-Plese, S.; Lacet, B.; Raus, J.; Weiner, H.L.; Hafler, D.A. Increased frequency of interleukin 2-responsive T cells specific for myelin basic protein and proteolipid protein in peripheral blood and cerebrospinal fluid of patients with multiple sclerosis. J. Exp. Med. 1994, 179, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; De Rosa, V.; Carrieri, P.B.; Montella, S.; Bruzzese, D.; Porcellini, A.; Procaccini, C.; La Cava, A.; Matarese, G. Regulatory T cell proliferative potential is impaired in human autoimmune disease. Nat. Med. 2014, 20, 69–74. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, V.; Galgani, M.; Porcellini, A.; Colamatteo, A.; Santopaolo, M.; Zuchegna, C.; Romano, A.; De Simone, S.; Procaccini, C.; La Rocca, C.; et al. Glycolysis controls the induction of human regulatory T cells by modulating the expression of FOXP3 exon 2 splicing variants. Nat. Immunol. 2015, 16, 1174–1184. [Google Scholar] [CrossRef]
- Batoulis, H.; Addicks, K.; Kuerten, S. Emerging concepts in autoimmune encephalomyelitis beyond the CD4/T(H)1 paradigm. Ann. Anat. 2010, 192, 179–193. [Google Scholar] [CrossRef]
- Rayatpour, A. and Javan, M. Targeting the brain lesions using peptides: A review focused on the possibility of targeted drug delivery to multiple sclerosis lesions. Pharmacol. Res. 2021, 167, 105441. [Google Scholar] [CrossRef]
- Badawi, A.H.; Siahaan, T.J. Immune modulating peptides for the treatment and suppression of multiple sclerosis. Clin. Immunol. 2012, 144, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. Immunology: A tolerant approach. Nature 2014, 507, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Ponsford, M.; Mazza, G.; Coad, J.; Campbell, M.J.; Zajicek, J.; Wraith, D.C. Differential responses of CD45+ve T-cell subsets to MBP in multiple sclerosis. Clin. Exp. Immunol. 2001, 124, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Ponsford, M.; Lowrey, P.; Campbell, M.J.; Zajicek, J.; Wraith, D.C. Diversity and dynamics of the T-cell response to MBP in DR2+ve individuals. Clin. Exp. Immunol. 2002, 128, 538–547. [Google Scholar] [CrossRef]
- Etzensperger, R.; McMahon, R.M.; Jones, E.Y.; Fugger, L. Dissection of the multiple sclerosis associated DR2 haplotype. J. Autoimmun. 2008, 31, 201–207. [Google Scholar] [CrossRef]
- Ramagopalan, S.V.; Ebers, G.C. Multiple sclerosis: Major histocompatibility complexity and antigen presentation. Genome. Med. 2009, 1, 105. [Google Scholar] [CrossRef] [PubMed]
- Streeter, H.B.; Rigden, R.; Martin, K.F.; Scolding, N.J.; Wraith, D.C. Preclinical development and first-in-human study of ATX-MS-1467 for immunotherapy of MS. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, 93. [Google Scholar] [CrossRef] [PubMed]
- De Souza, A.L.S.; Rudin, S.; Chang, R.; Mitchell, K.; Crandall, T.; Huang, S.; Choi, J.K.; Okitsu, S.L.; Graham, D.L.; Tomkinson, B.; et al. ATX-MS-1467 Induces Long-Term Tolerance to Myelin Basic Protein in (DR2 x Ob1)F1 Mice by Induction of IL-10-Secreting iTregs. Neurol. Ther. 2018, 7, 103–128. [Google Scholar] [CrossRef] [PubMed]
- Tselios, T.; Daliani, I.; Deraos, S.; Thymianou, S.; Matsoukas, E.; Troganis, A.; Gerothanassis, I.; Mouzaki, A.; Mavromoustakos, T.; Probert, L.; et al. Treatment of experimental allergic encephalomyelitis (EAE) by a rationally designed cyclic analogue of myelin basic protein (MBP) epitope 72-85. Bioorg. Med. Chem. Lett. 2000, 10, 2713–2717. [Google Scholar] [CrossRef]
- Tselios, T.; Apostolopoulos, V.; Daliani, I.; Deraos, S.; Grdadolnik, S.; Mavromoustakos, T.; Melachrinou, M.; Thymianou, S.; Probert, L.; Mouzaki, A.; et al. Antagonistic effects of human cyclic MBP (87–99) altered peptide ligands in experimental allergic encephalomyelitis and human T-cell proliferation. J. Med. Chem. 2002, 45, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Matsoukas, J.; Apostolopoulos, V.; Kalbacher, H.; Papini, A.M.; Tselios, T.; Chatzantoni, K.; Biagioli, T.; Lolli, F.; Deraos, S.; Papathanassopoulos, P.; et al. Design and synthesis of a novel potent myelin basic protein epitope 87-99 cyclic analogue: Enhanced stability and biological properties of mimics render them a potentially new class of immunomodulators. J. Med. Chem. 2005, 48, 1470–1480. [Google Scholar] [CrossRef]
- Deraos, G.; Rodi, M.; Kalbacher, H.; Chatzantoni, K.; Karagiannis, F.; Synodinos, L.; Plotas, P.; Papalois, A.; Dimisianos, N.; Papathanasopoulos, P.; et al. Properties of myelin altered peptide ligand cyclo (87-99)(Ala91,Ala96) MBP87-99 render it a promising drug lead for immunotherapy of multiple sclerosis. Eur. J. Med. Chem. 2015, 101, 13–23. [Google Scholar] [CrossRef]
- Katsara, M.; Deraos, G.; Tselios, T.; Matsoukas, J.; Apostolopoulos, V. Design of novel cyclic altered peptide ligands of myelin basic protein MBP83-99 that modulate immune responses in SJL/J mice. J. Med. Chem. 2008, 51, 3971–3978. [Google Scholar] [CrossRef]
- Katsara, M.; Deraos, G.; Tselios, T.; Matsoukas, M.T.; Friligou, I.; Matsoukas, J.; Apostolopoulos, V. Design and synthesis of a cyclic double mutant peptide (cyclo (87-99)[A91,A96] MBP87-99) induces altered responses in mice after conjugation to mannan: Implications in the immunotherapy of multiple sclerosis. J. Med. Chem. 2009, 52, 214–218. [Google Scholar] [CrossRef]
- Gaur, A.; Boehme, S.A.; Chalmers, D.; Crowe, P.D.; Pahuja, A.; Ling, N.; Brocke, S.; Steinman, L.; Conlon, P.J. Amelioration of relapsing experimental autoimmune encephalomyelitis with altered myelin basic protein peptides involves different cellular mechanisms. J. Neuroimmunol. 1997, 74, 149–158. [Google Scholar] [CrossRef]
- Karin, N.; Mitchell, D.J.; Brocke, S.; Ling, N.; Steinman, L. Reversal of experimental autoimmune encephalomyelitis by a soluble peptide variant of a myelin basic protein epitope: T cell receptor antagonism and reduction of interferon gamma and tumor necrosis factor alpha production. J. Exp. Med. 1994, 180, 2227–2237. [Google Scholar] [CrossRef]
- Emmanouil, M.; Tseveleki, V.; Triantafyllakou, I.; Nteli, A.; Tselios, T.; Probert, L. A Cyclic Altered Peptide Analogue Based on Myelin Basic Protein 87-99 Provides Lasting Prophylactic and Therapeutic Protection Against Acute Experimental Autoimmune Encephalomyelitis. Molecules 2018, 23, 304. [Google Scholar] [CrossRef]
- Mangan, P.R.; Harrington, L.E.; O’Quinn, D.B.; Helms, W.S.; Bullard, D.C.; Elson, C.O.; Hatton, R.D.; Wahl, S.M.; Schoeb, T.R.; Weaver, C.T. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature 2006, 441, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Flavell, R.A.; Stockinger, B. Signals mediated by transforming growth factor-beta initiate autoimmune encephalomyelitis, but chronic inflammation is needed to sustain disease. Nat. Immunol. 2006, 7, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Chang, S.H.; Martinez, G.J.; Yang, X.O.; Nurieva, R.; Kang, H.S.; Ma, L.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; et al. Critical regulation of early Th17 cell differentiation by interleukin-1 signaling. Immunity 2009, 30, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Goedkoop, A.Y.; Kraan, M.C.; Picavet, D.I.; de Rie, M.A.; Teunissen, M.B.; Bos, J.D.; Tak, P.P. Deactivation of endothelium and reduction in angiogenesis in psoriatic skin and synovium by low dose infliximab therapy in combination with stable methotrexate therapy: A prospective single-centre study. Arthritis Res. Ther. 2004, 6, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Canete, J.D.; Pablos, J.L.; Sanmarti, R.; Mallofre, C.; Marsal, S.; Maymo, J.; Gratacos, J.; Mezquita, J.; Mezquita, C.; Cid, M.C. Antiangiogenic effects of anti-tumor necrosis factor alpha therapy with infliximab in psoriatic arthritis. Arthritis Rheum. 2004, 50, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Kurohori, Y.; Sato, K.; Suzuki, S.; Kashiwazaki, S. Adhesion molecule expression on peripheral blood mononuclear cells in rheumatoid arthritis: Positive correlation between the proportion of L-selectin and disease activity. Clin. Rheumatol. 1995, 14, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, G.; Mittal, A.; Weiner, H.L. Increased osteopontin expression in dendritic cells amplifies IL-17 production by CD4+ T cells in experimental autoimmune encephalomyelitis and in multiple sclerosis. J. Immunol. 2008, 181, 7480–7488. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L. Shifting therapeutic attention in MS to osteopontin, type 1 and type 2 IFN. Eur. J. Immunol. 2009, 39, 2358–2360. [Google Scholar] [CrossRef]
- Han, M.H.; Hwang, S.I.; Roy, D.B.; Lundgren, D.H.; Price, J.V.; Ousman, S.S.; Fernald, G.H.; Gerlitz, B.; Robinson, W.H.; Baranzini, S.E.; et al. Proteomic analysis of active multiple sclerosis lesions reveals therapeutic targets. Nature 2008, 451, 1076–1081. [Google Scholar] [CrossRef]
- Teesalu, T.; Hinkkanen, A.E.; Vaheri, A. Coordinated induction of extracellular proteolysis systems during experimental autoimmune encephalomyelitis in mice. Am. J. Pathol. 2001, 159, 2227–2237. [Google Scholar] [CrossRef]
- Du, F.; Garg, A.V.; Kosar, K.; Majumder, S.; Kugler, D.G.; Mir, G.H.; Maggio, M.; Henkel, M.; Lacy-Hulbert, A.; McGeachy, M.J. Inflammatory Th17 Cells Express Integrin alphavbeta3 for Pathogenic Function. Cell Rep. 2016, 16, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Comegna, D.; Zannetti, A.; Del Gatto, A.; de Paola, I.; Russo, L.; Di Gaetano, S.; Liguoro, A.; Capasso, D.; Saviano, M.; Zaccaro, L. Chemical Modification for Proteolytic Stabilization of the Selective alphavbeta3 Integrin RGDechi Peptide: In Vitro and in Vivo Activities on Malignant Melanoma Cells. J. Med. Chem. 2017, 60, 9874–9884. [Google Scholar]
- Bolzati, C.; Salvarese, N.; Carpanese, D.; Seraglia, R.; Melendez-Alafort, L.; Rosato, A.; Capasso, D.; Saviano, M.; Del Gatto, A.; Comegna, D.; et al. [(99m)Tc][Tc(N)PNP43]-Labeled RGD Peptides As New Probes for a Selective Detection of alphavbeta3 Integrin: Synthesis, Structure-Activity and Pharmacokinetic Studies. J. Med. Chem. 2018, 61, 9596–9610. [Google Scholar] [CrossRef]
- Komegae, E.N.; Souza, T.A.; Grund, L.Z.; Lima, C.; Lopes-Ferreira, M. Multiple functional therapeutic effects of TnP: A small stable synthetic peptide derived from fish venom in a mouse model of multiple sclerosis. PLoS ONE 2017, 12, 171796. [Google Scholar] [CrossRef]
- Baxter, A.G. The origin and application of experimental autoimmune encephalomyelitis. Nat. Rev. Immunol. 2007, 7, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, M.M.; Saito, Y.; Knapp, G.W.; Ting, J.P.; Suzuki, K.; Matsushima, G.K. Microglial/macrophage accumulation during cuprizone-induced demyelination in C57BL/6 mice. J. Neuroimmunol. 1998, 92, 38–49. [Google Scholar] [CrossRef]
- Praet, J.; Guglielmetti, C.; Berneman, Z.; Van der Linden, A.; Ponsaerts, P. Cellular and molecular neuropathology of the cuprizone mouse model: Clinical relevance for multiple sclerosis. Neurosci. Biobehav. Rev. 2014, 47, 485–505. [Google Scholar] [CrossRef]
- Craik, D.J. Plant cyclotides: Circular, knotted peptide toxins. Toxicon 2001, 39, 1809–1813. [Google Scholar] [CrossRef]
- Grundemann, C.; Thell, K.; Lengen, K.; Garcia-Kaufer, M.; Huang, Y.H.; Huber, R.; Craik, D.J.; Schabbauer, G.; Gruber, C.W. Cyclotides Suppress Human T-Lymphocyte Proliferation by an Interleukin 2-Dependent Mechanism. PLoS ONE 2013, 8, 68016. [Google Scholar] [CrossRef]
- Thell, K.; Hellinger, R.; Sahin, E.; Michenthaler, P.; Gold-Binder, M.; Haider, T.; Kuttke, M.; Liutkeviciute, Z.; Goransson, U.; Grundemann, C.; et al. Oral activity of a nature-derived cyclic peptide for the treatment of multiple sclerosis. Proc. Natl. Acad. Sci. USA 2016, 113, 3960–3965. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Stone, S.; Lin, W. Role of nuclear factor kappaB in multiple sclerosis and experimental autoimmune encephalomyelitis. Neural. Regen. Res. 2018, 13, 1507–1515. [Google Scholar]
- Lunin, S.M.; Khrenov, M.O.; Glushkova, O.V.; Parfenyuk, S.B.; Novoselova, T.V.; Novoselova, E.G. Immune response in the relapsing-remitting experimental autoimmune encephalomyelitis in mice: The role of the NF-kappaB signaling pathway. Cell Immunol. 2019, 336, 20–27. [Google Scholar] [CrossRef]
- Lunin, S.M.; Novoselova, E.G. Thymus hormones as prospective anti-inflammatory agents. Expert. Opin. Ther. Targets 2010, 14, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Lunin, S.M.; Khrenov, M.O.; Novoselova, T.V.; Parfenyuk, S.B.; Novoselova, E.G. Thymulin, a thymic peptide, prevents the overproduction of pro-inflammatory cytokines and heat shock protein Hsp70 in inflammation-bearing mice. Immunol. Investig. 2008, 37, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Novoselova, E.G.; Khrenov, M.O.; Glushkova, O.V.; Lunin, S.M.; Parfenyuk, S.B.; Novoselova, T.V.; Fesenko, E.E. Anti-inflammatory effects of IKK inhibitor XII, thymulin, and fat-soluble antioxidants in LPS-treated mice. Mediat. Inflamm. 2014, 2014, 724838. [Google Scholar] [CrossRef] [PubMed]
- Safieh-Garabedian, B.; Nomikos, M.; Saade, N. Targeting inflammatory components in neuropathic pain: The analgesic effect of thymulin related peptide. Neurosci. Lett. 2019, 702, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Lunin, S.M.; Glushkova, O.V.; Khrenov, M.O.; Novoselova, T.V.; Parfenyuk, S.B.; Fesenko, E.E.; Novoselova, E.G. Thymic peptides restrain the inflammatory response in mice with experimental autoimmune encephalomyelitis. Immunobiology 2013, 218, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Lunin, S.M.; Khrenov, M.O.; Novoselova, T.V.; Parfenyuk, S.B.; Glushkova, O.V.; Fesenko, E.E.; Novoselova, E.G. Modulation of inflammatory response in mice with severe autoimmune disease by thymic peptide thymulin and an inhibitor of NF-kappaB signaling. Int. Immunopharmacol. 2015, 25, 260–266. [Google Scholar] [CrossRef]
- Lunin, S.M.; Khrenov, M.O.; Glushkova, O.V.; Parfenyuk, S.B.; Novoselova, T.V.; Novoselova, E.G. Protective Effect of PBCA Nanoparticles Loaded with Thymulin Against the Relapsing-Remitting Form of Experimental Autoimmune Encephalomyelitis in Mice. Int. J. Mol. Sci. 2019, 20, 5374. [Google Scholar] [CrossRef]
- Novoselova, E.G.; Lunin, S.M.; Glushkova, O.V.; Khrenov, M.O.; Parfenyuk, S.B.; Zakharova, N.M.; Fesenko, E.E. Thymulin, free or bound to PBCA nanoparticles, protects mice against chronic septic inflammation. PLoS ONE 2018, 13, 197601. [Google Scholar] [CrossRef] [PubMed]
- Stangel, M.; Kuhlmann, T.; Matthews, P.M.; Kilpatrick, T.J. Achievements and obstacles of remyelinating therapies in multiple sclerosis. Nat. Rev. Neurol. 2017, 13, 742–754. [Google Scholar] [CrossRef]
- Fletcher, J.L.; Wood, R.J.; Nguyen, J.; Norman, E.M.L.; Jun, C.M.K.; Prawdiuk, A.R.; Biemond, M.; Nguyen, H.T.H.; Northfield, S.E.; Hughes, R.A.; et al. Targeting TrkB with a Brain-Derived Neurotrophic Factor Mimetic Promotes Myelin Repair in the Brain. J. Neurosci. 2018, 38, 7088–7099. [Google Scholar] [CrossRef]
- Minagar, A.; Alexander, J.S. Blood-brain barrier disruption in multiple sclerosis. Mult. Scler. 2003, 9, 540–549. [Google Scholar] [CrossRef]
- Adams, R.A.; Bauer, J.; Flick, M.J.; Sikorski, S.L.; Nuriel, T.; Lassmann, H.; Degen, J.L.; Akassoglou, K. The fibrin-derived gamma377–395 peptide inhibits microglia activation and suppresses relapsing paralysis in central nervous system autoimmune disease. J. Exp. Med. 2007, 204, 571–582. [Google Scholar] [CrossRef]
- Yuan, R.; Maeda, Y.; Li, W.; Lu, W.; Cook, S.; Dowling, P. Erythropoietin: A potent inducer of peripheral immuno/inflammatory modulation in autoimmune EAE. PLoS ONE 2008, 3, 1924. [Google Scholar] [CrossRef] [PubMed]
- Dicato, M. Venous thromboembolic events and erythropoiesis-stimulating agents: An update. Oncologist 2008, 13, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Ehrenreich, H.; Weissenborn, K.; Prange, H.; Schneider, D.; Weimar, C.; Wartenberg, K.; Schellinger, P.D.; Bohn, M.; Becker, H.; Wegrzyn, M.; et al. Recombinant human erythropoietin in the treatment of acute ischemic stroke. Stroke 2009, 40, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Corwin, H.L.; Gettinger, A.; Fabian, T.C.; May, A.; Pearl, R.G.; Heard, S.; An, R.; Bowers, P.J.; Burton, P.; Klausner, M.A.; et al. Efficacy and safety of epoetin alfa in critically ill patients. N. Engl. J. Med. 2007, 357, 965–976. [Google Scholar] [CrossRef]
- Yuan, R.; Wang, B.; Lu, W.; Maeda, Y.; Dowling, P. A Distinct Region in Erythropoietin that Induces Immuno/Inflammatory Modulation and Tissue Protection. Neurotherapeutics 2015, 12, 850–861. [Google Scholar] [CrossRef]
- Wang, B.; Kang, M.; Marchese, M.; Rodriguez, E.; Lu, W.; Li, X.; Maeda, Y.; Dowling, P. Beneficial Effect of Erythropoietin Short Peptide on Acute Traumatic Brain Injury. Neurotherapeutics 2016, 13, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Maeda, Y.; Yuan, R.R.; Elkabes, S.; Cook, S.; Dowling, P. Beneficial effect of erythropoietin on experimental allergic encephalomyelitis. Ann. Neurol. 2004, 56, 767–777. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Gaindh, D.; Choi, Y.B.; Marchese, M.; Dowling, P.; Cook, S.; Blumberg, B.; Park, J.H.; Lu, W. Prolonged Beneficial Effect of Brief Erythropoietin Peptide JM4 Therapy on Chronic Relapsing EAE. Neurotherapeutics 2020, 18, 401–411. [Google Scholar] [CrossRef]
- Sorensen, P.S.; Sellebjerg, F. Pulsed immune reconstitution therapy in multiple sclerosis. Ther. Adv. Neurol. Disord. 2019, 12. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Gatto, A.; Saviano, M.; Zaccaro, L. An Overview of Peptide-Based Molecules as Potential Drug Candidates for Multiple Sclerosis. Molecules 2021, 26, 5227. https://doi.org/10.3390/molecules26175227
Del Gatto A, Saviano M, Zaccaro L. An Overview of Peptide-Based Molecules as Potential Drug Candidates for Multiple Sclerosis. Molecules. 2021; 26(17):5227. https://doi.org/10.3390/molecules26175227
Chicago/Turabian StyleDel Gatto, Annarita, Michele Saviano, and Laura Zaccaro. 2021. "An Overview of Peptide-Based Molecules as Potential Drug Candidates for Multiple Sclerosis" Molecules 26, no. 17: 5227. https://doi.org/10.3390/molecules26175227
APA StyleDel Gatto, A., Saviano, M., & Zaccaro, L. (2021). An Overview of Peptide-Based Molecules as Potential Drug Candidates for Multiple Sclerosis. Molecules, 26(17), 5227. https://doi.org/10.3390/molecules26175227