1. Introduction
Mussels are highly consumed in several countries. Asia and Europe are considered the leading producers, estimated to produce about 1.05 and 0.5 million tonnes of mussel per year, respectively [
1,
2]. Mussel consumption has several advantages, for both the environment and consumers. Environmentally, mussel farming can be done with minimal greenhouse gas emissions, and thus low carbon footprint and few environmental impacts [
3]. Mussels farming produces about 0.6 kg of CO
2 emission/kg edible product, while beef produces about 19.0–36.7 kg of CO
2 emission/kg edible product [
3]. For consumers, mussel meat has low fat and low calories. Still, more importantly, mussels are a rich source of sodium, selenium, vitamin B twelve, zinc [
1], and an interesting source of proteins since they are composed of about 58.7% of protein on a dry weight basis [
4]. Due to their protein-rich meat, mussels have been described as a source of bioactive peptides with relevant biological properties. Bioactive peptides are fragments that are inert when inside proteins but show different properties when broken from the original protein [
5]. Thus, enzymatic hydrolysis with proteases seems an interesting approach for obtaining bioactive extracts since these enzymes may break mussel proteins into smaller peptides, which may be associated with other biological and functional properties [
6]. Different enzymes have been used to produce bioactive peptides from mussels, such as pepsin [
7], flavourzyme [
8], papain [
8], and trypsin [
9]. Marine species have often been described as a source of bioactive peptides, and some bioactivities have been associated to mussel peptides, such as antioxidant [
10], anti-hypertensive [
11], antimicrobial [
12], anticancer [
7], anti-inflammatory [
13], anticoagulant [
14], antidiabetic [
15], and antiviral [
16]. As far as we know,
Mytilus galloprovincialis bioactive extracts are not so extensively explored, with the main studies being developed with
Mytilus coruscus and
Mytilus edulis. The farming of
M. galloprovincialis, also known as the Mediterranean mussel, has mainly been developing along the Spanish Atlantic coast and in the Mediterranean area [
2].
Mussel commercialisation generates losses and waste since they are submitted to a pre-selection before being delivered for sale, resulting in the rejection of broken mussels or those which fail to meet established criteria in the target market [
17,
18,
19,
20]. It is estimated that about 27% of produced mussels are discarded [
4,
20]. Thus, mussel meat waste can be used to produce bioactive hydrolysates with interesting properties for food, cosmetic, pharmaceutical, and nutraceutical industrial applications.
In this work, Mytilus galloprovincialis meat was submitted to different conditions according to two factorial designs to produce hydrolysates with a maximum level of soluble rich proteins/peptides and bioactive properties, more specifically antioxidant and anti-hypertensive. The hydrolysates were produced by enzymatic hydrolysis using two different enzymes, subtilisin and corolase. Therefore, this study aims to valorise mussel meat by creating bioactive hydrolysates with potential for various industries.
3. Discussion
Mussel meat has a high protein content, making it interesting to produce bioactive hydrolysates rich in proteins and bioactive peptides. However, the mussel Mytilus galloprovincialis is less exploited regarding its bioactive potential when compared to other mussel species, such as M. coruscus and M. edulis. Since we wanted to create a food-grade method, we chose two food-grade proteases to carry out enzymatic hydrolysis. Thus, to explore this mussel potential, we have performed two Box–Behken experimental designs, with two different proteases, aiming to obtain hydrolysates with interesting potential for industrial applications. Furthermore, we have not found studies with mussels from the genus Mytilus performing enzymatic hydrolysis with subtilisin or corolase. The most frequent enzymes found were mainly gastric enzymes, such as pepsin and trypsin, and non-gastric enzymes, such as papain and flavourzyme.
The mussel meat biomass used showed 70.50 ± 13.44% of protein on a dry weight (DW) basis and a moisture content of 90.30 ± 4.24%. These results show higher values of protein when compared to other studies with
Mytilus sp. from Portugal and Spain that showed protein content variation from 39.17–42.94 (DW) and moisture % of 81.71–87.59% [
23]. However, these results are in line with the possible variations in protein content that can occur in different months, as shown by Çelik [
24] in a study with
Mytilus galloprovincialis indicating higher protein levels (74.64%) in February.
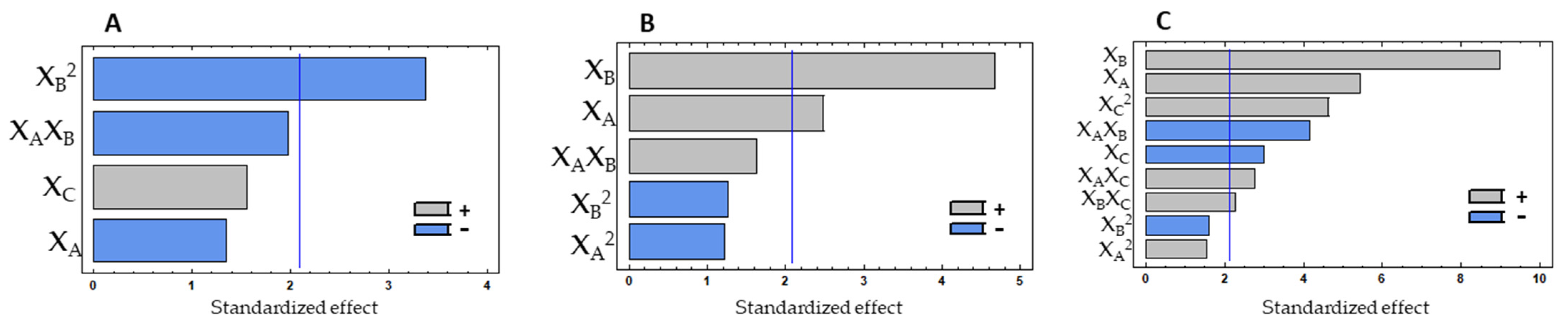
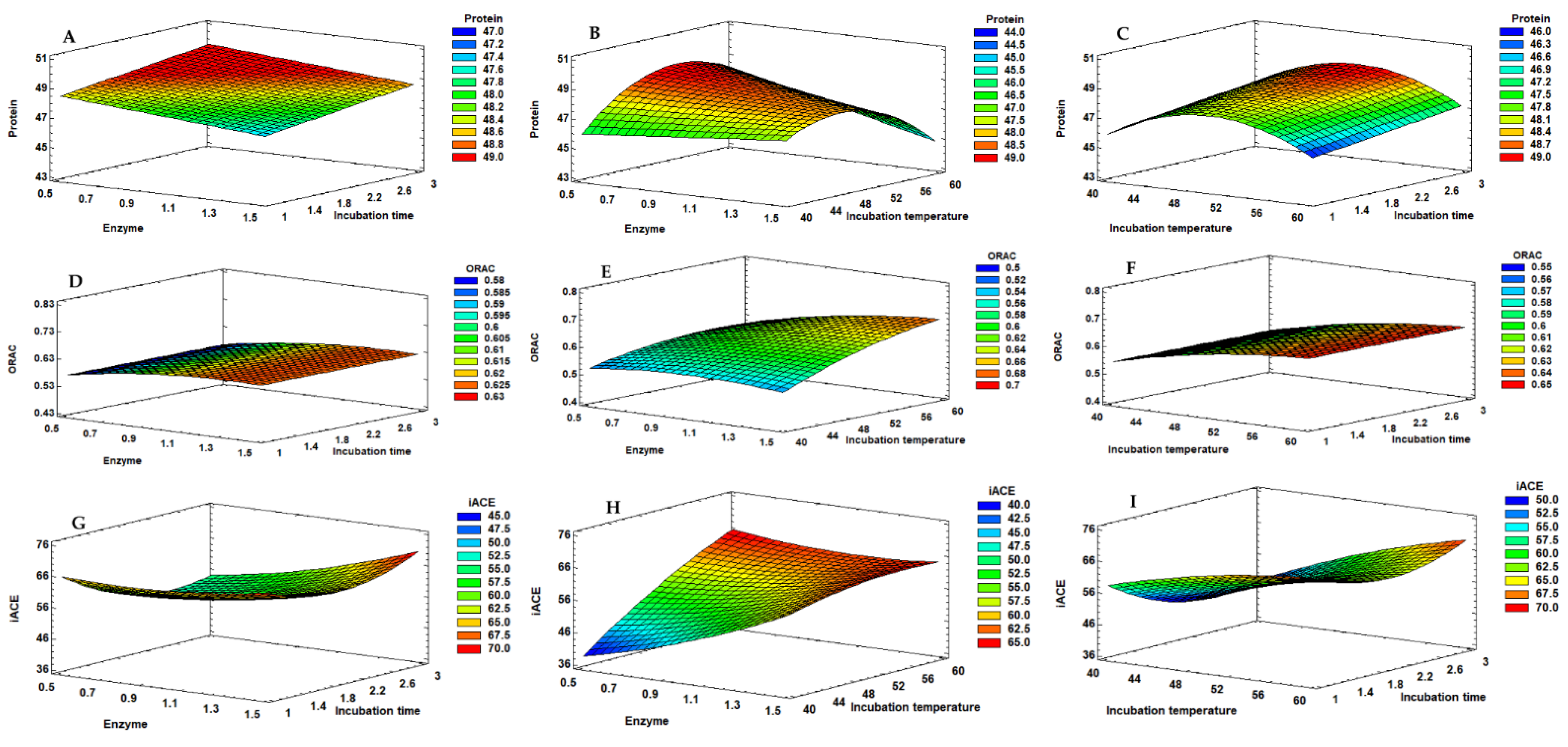
The protein % of the hydrolysates does not seem to be highly influenced by the determined models, indicating that enzymatic hydrolysis with both enzymes can produce hydrolysates with protein contents in the range of 40–48% (DW). So, to obtain mussel hydrolysates with a content of above 40%, the most economical and fastest conditions can probably be used.
The subtilisin protease optimised method was an enzymatic hydrolysis with 1.5% of enzyme with a duration of 3 h at 52 °C. In a scale-up test with these conditions, the final hydrolysates showed protein content, ORAC, and IC50 for ACE inhibition values of 45.0 ± 0.38, 0.49 ± 0.061 245 μmol TE/mg hydrolysate, and 1.0 ± 0.56 mg protein/mL, respectively. With the corolase, the optimised method was an enzymatic hydrolysis with 3.0% of enzyme with a duration of 3 h at 40 °C, obtaining scale-up hydrolysates with protein content, ORAC, and IC
50 for ACE inhibition values of 46.35 ± 1.12, 0.389 ± 0.021 μmol TE/mg hydrolysate, and 3.7 ± 0.33 mg protein/mL, respectively. The experimental design responses were not highly explained by the models, indicating that the system is highly variable, as necessary to enhance the process, or a plateau may have been quickly reached, which challenges the explanation of the variability in the models. However, the hydrolysates showed potential as proteins/peptides sources with antioxidant properties, bringing interest to the results. In both experiments, interesting protein values were obtained with a few hours of hydrolysis, which is in line with other studies showing that enzymatic hydrolysis with papain for 2 h was enough for achieving the maximum protein extraction [
25]. The obtained protein content (450 and 463 mg protein/g hydrolysate) was close to those obtained for
Mytilus edulis by Vareltzis and Undeland (430 and 580 mg protein/g with acid and alkaline process, respectively) [
26], but lower than those obtained by Neves et al. (735.45 ± 11.45 mg protein/g hydrolysate) [
15]. The subtilisin method needs a lower enzyme% to obtain higher bioactive properties than the corolase, with the main difference being observed for the anti-hypertensive potential. Even though the corolase hydrolysate shows a higher protein %, this does not bring much potential for this hydrolysate due to the small difference compared to the subtilisin hydrolysate. So, mussel meat hydrolysate produced with the subtilisin protease appears to have more potential for further studies as an active ingredient, at least regarding the antioxidant and anti-hypertensive potential. However, it is important to highlight that the obtained values for the anti-hypertensive property are not very significant since IC
50 ≥ 1000 µg protein/mL [
27]. The hydrolysate produced with corolase shows the lowest potential with an IC
50 = 3700 µg protein/mL. The subtilisin hydrolysate seems to be more promising, with an IC
50 = 1000 µg protein/mL. Bioactive peptides usually have a molecular weight (MW) less than 6 KDa [
28], and the most efficient anti-hypertensive peptides are usually associated with MW lower than 3 KDa [
29]. Several marine derived peptides with MW lower than 3 KDa have been described, such as the microalgae
Chlorella vulgaris VECYGPNRPQF peptide (1.3 KDa; IC
50 of 29.6 µM) [
30] and
C. ellipsoidea VEGY peptide (467 Da; IC
50 of 128.4 µM) [
31]; the macroalgae
Gracilariopsis lemaneiformis TGAPCR peptide (604 Da; IC
50 of 23.94 µM) [
32] and
Nannochloropsis oculata LEQ peptide (369 Da; IC
50 of 173 µM) [
33]. Thus, to increase the anti-hypertensive potential of the produced hydrolysates, a future approach may be to submit them to a ultrafiltration system using 3-KDa cut-offs, to concentrate peptides with lower MW [
34]. Furthermore, the production of low MW peptides may also increase the antimicrobial potential of hydrolysates, thus presenting new possible applications.
All the hydrolysis performed used the same mussel batch, initially minced and stored at −20 °C. The main goal was to assure that all the hydrolysis were performed with minimum mussel internal variations, since we wanted to compare a large number of extracts to optimize the hydrolysate production. The validation and scaled-up hydrolysis were also performed with the same batch, allowing us to precisely compare these extracts with those obtained using the experimental design, excluding possible mussel internal chemical variations. However, it is important to point out that mussel meat biochemical composition varies with the harvesting season, due to their reproductive cycle, environmental conditions, growth, and food availability [
24]. Çelik et al. [
24] showed that mussel protein content is highly related with the spawning seasons, with decreased protein levels being observed during this season, which increases after spawning time. So, different harvesting seasons lead to variations in the biochemical composition, which may be reflected in differences in mussel protein and amino acids, not only in terms of quantity, but also quality. Consequently, the enzyme action will produce different peptides over the seasons. Therefore, it would be expected that the ORAC and iACE results obtained for hydrolysates produced with the presented methods may differ between different mussel batches, depending on their harvesting season and other external factors. Furthermore, the mussel’s digestive gland produces proteases, which also seems to be influenced by their diet [
35], and mussels seem able to modulate their digestive enzyme activities in response to limited feeding and thermal stress [
36]. Since endogenous proteases may also have either a proteolytic effect or serve as an enzymatic substrate in the hydrolysis, the amount of endogenous proteases may also contribute to the variability of results. Thus, in the future, it would be of great interest to perform the same hydrolysis in different mussel batches, harvested in different months, and perhaps from different locations, to examine the variability of the produced hydrolysates when influenced by the expected biochemical composition differences.
The production of multifunctional extracts from mussels may be an interesting approach for food applications since they are not only a source of proteins, but also present bioactivities that can enhance consumer health, useful for the creation of functional food. Moreover, they may also be used as nutraceuticals or as cosmetic ingredients. Antioxidant food and nutraceuticals may help reduce levels of radical oxygen species that are constantly produced by the human organism, especially during high exposure to external factors, such as alcohol, tobacco smoke, and stress [
37]. Hypertension has been associated as one of the main causes of cardiovascular diseases [
38], with the angiotensin-converting enzyme (ACE) being one of the major enzymes involved in the process of blood pressure regulation [
39]. Thus, multifunctional extracts may be incorporated in food matrices with health claims, to facilitate sale as functional food. However, for claiming health benefits, it is important to study the bioavailability of food matrices incorporating these hydrolysates [
40], by analysing their resistance to the gastrointestinal (GI) tract enzymes and conditions, to verify if their properties are maintained throughout the GI tract passage [
41]. In cosmetics, antioxidants are especially important for anti-ageing purposes since free radicals are highly associated with skin ageing. Thus, natural antioxidant hydrolysates used as active cosmetic ingredients may help decrease free radical damage and work as an alternative for synthetic antioxidant ingredients.
Furthermore, mussel protein and peptide hydrolysates are frequently associated with other properties, especially antimicrobial properties [
42,
43], but also anticancer [
44], anti-inflammatory [
45], anticoagulant [
14], antidiabetic [
15], and antiviral [
46]. Thus, in the future, it would be interesting to study these hydrolysates for other bioactivities. Additionally, the water-soluble nature of these extracts makes it easy to incorporate them in several matrices. Although the freeze-drying process may lead to a loss of bioactivity, it is important for a better preservation of the hydrolysates, facilitating their incorporation in both solid and liquid matrices.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}