
Novel Putative Positive Modulators of α4β2 nAChRs Potentiate Nicotine Reward-Related Behavior




Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Reagents and Dose Selection
4.3. Conditioned Place Preference (CPP) Assays
4.4. Neuro-2a Cell Culture and Transient Transfections
4.5. Calcium 6 Assay (Calcium Accumulation Assay)
4.6. Calculations
4.7. Statistical Analysis
4.8. Supplemental Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Li, M.D.; Beuten, J.; Ma, J.Z.; Payne, T.J.; Lou, X.Y.; Garcia, V.; Duenes, A.S.; Crews, K.M.; Elston, R.C. Ethnic- and gender-specific association of the nicotinic acetylcholine receptor alpha4 subunit gene (CHRNA4) with nicotine dependence. Hum. Mol. Genet. 2005, 14, 1211–12199. [Google Scholar] [CrossRef] [Green Version]
- Ehringer, M.A.; Clegg, H.V.; Collins, A.C.; Corley, R.P.; Crowley, T.; Hewitt, J.K.; Hopfer, C.J.; Krauter, K.; Lessem, J.; Rhee, S.H.; et al. Association of the neuronal nicotinic receptor beta2 subunit gene (CHRNB2) with subjective responses to alcohol and nicotine. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144, 596–604. [Google Scholar] [CrossRef]
- Mihalak, B.K.; Carroll, F.I.; Luetje, C.W. Varenicline is a partial agonist at α4β2 and a full agonist at α7 neuronal nicotinic receptors. Mol. Pharm. 2006, 70, 801–805. [Google Scholar] [CrossRef] [Green Version]
- Coe, J.W.; Brooks, P.R.; Vetelino, M.G.; Wirtz, M.C.; Arnold, E.P.; Huang, J.; Sands, S.B.; Davis, T.I.; Lebel, L.A.; Fox, C.B.; et al. Varenicline: An a4b2 nicotinic receptor partial agonist for smoking cessation. J. Med. Chem. 2005, 48, 3474–3477. [Google Scholar] [CrossRef] [PubMed]
- Richmond, R.; Zwar, N. Review of bupropion for smoking cessation. Drug Alcohol. Rev. 2003, 22, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Slemmer, J.E.; Martin, B.R.; Damaj, M.I. Bupropion is a nicotinic antagonist. J. Pharm. Exp. 2000, 295, 321–327. [Google Scholar]
- Benli, A.R.; Erturhan, S.; Oruc, M.A.; Kalpakci, P.; Sunay, D.; Demirel, Y. A comparison of the efficacy of varenicline and bupropion and an evaluation of the effect of the medications in the context of the smoking cessation programme. Tob. Induc. Dis. 2017, 15, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, K.; Gees, M.; Karashima, Y.; Meseguer, V.M.; Vanoirbeek, J.A.J.; Damann, N.; Everaerts, W.; Benoit, M.; Janssens, A.; Vennekens, R.; et al. Nicotine activates the chemosensory cation channel TRPA1. Nat. Neurosci. 2009, 12, 1293–1299. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Li, W.; Ward, A.; Piggott, B.J.; Larkspur, E.R.; Sternberg, P.; Xu, X.S. A C. elegans Model of Nicotine-Dependent Behavior: Regulation by TRP-Family Channels. Cell 2006, 127, 621–633. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-Maia, A.J.; Stapleton-Kotloski, J.R.; Lyall, V.; Phan, T.-H.T.; Mummalaneni, S.; Melone, P.; DeSimone, J.A.; Nicolelis, M.A.L.; Simon, S.A. Nicotine activates TRPM5-dependent and independent taste pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 1596–1601. [Google Scholar] [CrossRef] [Green Version]
- Willis, D.N.; Liu, B.; Ha, M.A.; Jordt, S.; Morris, J.B. Menthol attenuates respiratory irritation responses to multiple cigarette smoke irritants. FASEB J. 2011, 25, 4434–4444. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.A.; Smith, G.J.; Cichocki, J.A.; Fan, L.; Liu, Y.-S.; Caceres, A.I.; Jordt, S.E.; Morris, J.B. Menthol Attenuates Respiratory Irritation and Elevates Blood Cotinine in Cigarette Smoke Exposed Mice. PLoS ONE 2015, 10, e0117128. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Balakrishna, S.; Jabba, S.V.; Bonner, P.; Taylor, S.R.; Picciotto, M.R.; Jordt, S.-E. Menthol decreases oral nicotine aversion in C57BL/6 mice through a TRPM8-dependent mechanism. Tob. Control. 2016, 25, ii50–ii54. [Google Scholar] [CrossRef]
- Liu, B.-Y.; Lin, Y.-J.; Lee, H.-F.; Ho, C.-Y.; Ruan, T.; Kou, Y.R. Menthol suppresses laryngeal C-fiber hypersensitivity to cigarette smoke in a rat model of gastroesophageal reflux disease: The role of TRPM8. J. Appl. Physiol. 2015, 118, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.-H.; Liu, M.-H.; Ko, H.-K.B.; Perng, D.-W.; Lee, T.-S.; Kou, Y.R. Inflammatory Effects of Menthol vs. Non-menthol Cigarette Smoke Extract on Human Lung Epithelial Cells: A Double-Hit on TRPM8 by Reactive Oxygen Species and Menthol. Front. Physiol. 2017, 8, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, B.; Chen, H. Menthol facilitates the intravenous self-administration of nicotine in rats. Front. Behav. Neurosci. 2014, 8, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benowitz, N.L.; Herrera, B.; Jacob, P. Mentholated Cigarette Smoking Inhibits Nicotine Metabolism. J. Pharmacol. Exp. Ther. 2004, 310, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Teichert, R.W.; Memon, T.; Aman, J.W.; Olivera, B.M. Using constellation pharmacology to define comprehensively a somatosensory neuronal subclass. Proc. Natl. Acad. Sci. USA 2014, 111, 2319–2324. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.J.; Wall, T.R.; Henley, B.M.; Kim, C.H.; McKinney, S.; Lester, H.A. Menthol Enhances Nicotine Reward-Related Behavior by Potentiating Nicotine-Induced Changes in nAChR Function, nAChR Upregulation, and DA Neuron Excitability. Neuropsychopharmacology 2017, 42, 2285–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brody, A.L.; Mukhin, A.G.; La Charite, J.; Ta, K.; Farahi, J.; Sugar, C.A.; Mamoun, M.S.; Vellios, E.; Archie, M.; Kozman, M.; et al. Up-regulation of nicotinic acetylcholine receptors in menthol cigarette smokers. Int. J. Neuropsychopharmacol. 2013, 16, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, S.Y.; Akers, A.T.; Henderson, B.J. Flavors Enhance Nicotine Vapor Self-administration in Male Mice. Nicotine. Tob. Res. 2021, 23, 566–572. [Google Scholar] [CrossRef]
- Biswas, L.; Harrison, E.; Gong, Y.; Avusula, R.; Lee, J.; Zhang, M.; Rousselle, T.; Lage, J.; Liu, X. Enhancing effect of menthol on nicotine self-administration in rats. Psychopharmacology 2016, 233, 3417–3427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Harrison, E.; Biswas, L.; Tran, T.; Liu, X. Menthol facilitates dopamine-releasing effect of nicotine in rat nucleus accumbens. Pharmacol. Biochem. Behav. 2018, 175, 47–52. [Google Scholar] [CrossRef]
- Ahijevych, K.; Garrett, B.E. The Role of Menthol in Cigarettes as a Reinforcer of Smoking Behavior. Nicotine Tob. Res. 2010, 12, S110–S116. [Google Scholar] [CrossRef] [Green Version]
- Journigan, V.B.; Feng, Z.; Rahman, S.; Wang, Y.; Amin, A.R.M.R.; Heffner, C.E.; Bachtel, N.; Wang, S.; Gonzalez-Rodriguez, S.; Fernández-Carvajal, A.; et al. Structure-Based Design of Novel Biphenyl Amide Antagonists of Human Transient Receptor Potential Cation Channel Subfamily M Member 8 Channels with Potential Implications in the Treatment of Sensory Neuropathies. ACS Chem. Neurosci. 2020, 11, 268–290. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.J.; Grant, S.; Chu, B.W.; Shahoei, R.; Huard, S.M.; Saladi, S.S.M.; Tajkhorshid, E.; Dougherty, D.A.; Lester, H.A. Menthol stereoisomers exhibit different effects on a4b2 nAChR upregulation and dopamine neuron spontaneous firing. eNeuro 2018, 5, e0465-18.2018. [Google Scholar] [CrossRef] [Green Version]
- Avelar, A.J.; Akers, A.T.; Baumgard, Z.J.; Cooper, S.Y.; Casinelli, G.P.; Henderson, B.J. Why flavored vape products may be attractive: Green apple tobacco flavor elicits reward-related behavior, upregulates nAChRs on VTA dopamine neurons, and alters midbrain dopamine and GABA neuron function. Neuropharmacology 2019, 158, 107729. [Google Scholar] [CrossRef]
- Henderson, B.J.; Wall, T.; Henley, B.M.; Kim, C.H.; Nichols, W.A.; Moaddel, R.; Xiao, C.; Lester, H.A. Menthol alone upregulates midbrain nAChRs, alters nAChR sybtype stoichiometry, alters dopamine neuron firing frequency, and prevents nicotine reward. J. Neurosci. 2016, 36, 2957–2974. [Google Scholar] [CrossRef]
- Tapper, A.R.; McKinney, S.L.; Nashmi, R.; Schwarz, J.; Deshpande, P.; Labarca, C.; Whiteaker, P.; Marks, M.J.; Collins, A.C.; Lester, H.A. Nicotine activation of α4* receptors: Sufficient for reward, tolerance and sensitization. Science 2004, 306, 1029–1032. [Google Scholar] [CrossRef]
- Henderson, B.; Srinivasan, R.; Nichols, W.A.; Dilworth, C.N.; Gutierrez, D.F.; Mackey, E.D.; McKinney, S.; Drenan, R.M.; Richards, C.I.; Lester, H.A. Nicotine exploits a COPI-mediated process for chaperone-mediated up-regulation of its receptors. J. Gen. Physiol. 2013, 143, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Pantoja, R.; Moss, F.J.; Mackey, E.D.W.; Son, C.; Miwa, J.; Lester, H.A. Nicotine upregulates α4β2 nicotinic receptors and ER exit sites via stoichiometry-dependent chaperoning. J. Gen. Physiol. 2011, 137, 59–79. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.I.; Srinivasan, R.; Xiao, C.; Mackey, E.D.; Miwa, J.M.; Lester, H.A. Trafficking of α4* nicotinic receptors revealed by superecliptic phluorin: Effects of a β4 amyotrophic lateral sclerosis-associated mutation and chronic exposure to nicotine. J. Biol. Chem. 2011, 286, 31241–31249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.E.; Kuryatov, A.; Choi, C.H.; Zhou, Y.; Lindstrom, J. Alternate stoichiometries of a4b2 nicotinic acetylcholine receptors. Mol. Pharm. 2003, 63, 332–341. [Google Scholar] [CrossRef] [Green Version]
- González-Cestari, T.F.; Henderson, B.J.; Pavlovicz, R.E.; McKay, S.B.; El-Hajj, R.A.; Pulipaka, A.B.; Orac, C.M.; Reed, D.D.; Boyd, R.T.; Zhu, M.X.; et al. Effect of Novel Negative Allosteric Modulators of Neuronal Nicotinic Receptors on Cells Expressing Native and Recombinant Nicotinic Receptors: Implications for Drug Discovery. J. Pharmacol. Exp. Ther. 2008, 328, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Hans, M.; Wilhelm, M.; Swandulla, D. Menthol Suppresses Nicotinic Acetylcholine Receptor Functioning in Sensory Neurons via Allosteric Modulation. Chem. Senses 2012, 37, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.K.; Byun, N.; Bubser, M. Muscarinic and nicotinic acetylcholine receptor agonists and allosteric modulators for the treatment of schizophrenia. Neuropsychopharmacology 2012, 37, 16–42. [Google Scholar] [CrossRef] [Green Version]
- Ngolab, J.; Liu, L.; Zhao-Shea, R.; Gao, G.; Gardner, P.D.; Tapper, A.R. Functional Upregulation of α4* Nicotinic Acetylcholine Receptors in VTA GABAergic Neurons Increases Sensitivity to Nicotine Reward. J. Neurosci. 2015, 35, 8570–8578. [Google Scholar] [CrossRef] [PubMed]
- Nashmi, R.; Xiao, C.; Deshpande, P.; McKinney, S.; Grady, S.R.; Whiteaker, P.; Huang, Q.; McClure-Begley, T.; Lindstrom, J.M.; Labarca, C.; et al. Chronic nicotine cell specifically upregulates functional α4* nicotinic receptors: Basis for both tolerance in midbrain and enhanced long-term potentiation in perforant path. J. Neurosci. 2007, 27, 8202–8218. [Google Scholar] [CrossRef] [PubMed]
- Grieder, T.E.; Besson, M.; Maal-Bared, G.; Pons, S.; Maskos, U.; van der Kooy, D. beta2* nAChRs on VTA dopamine and GABA neurons separately mediate nicotine aversion and reward. Proc. Natl. Acad. Sci. USA 2019, 116, 25968–25973. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Tolentino, K.; Hopkins, C.R. Selective α7 nicotinic receptor agonists and positive allosteric modulators for the treatment of schizophrenia—a review. Expert Opin. Investig. Drugs 2020, 29, 603–610. [Google Scholar] [CrossRef]
- Timmermann, D.B.; Sandager-Nielsen, K.; Dyhring, T.; Smith, M.; Jacobsen, A.-M.; Nielsen, E.Ø.; Grunnet, M.; Christensen, J.K.; Peters, D.; Kohlhaas, K.; et al. Augmentation of cognitive function by NS9283, a stoichiometry-dependent positive allosteric modulator of α2- and α4-containing nicotinic acetylcholine receptors. Br. J. Pharmacol. 2012, 167, 164–182. [Google Scholar] [CrossRef]
- Ton, H.T.; Smart, A.E.; Aguilar, B.L.; Olson, T.T.; Kellar, K.J.; Ahern, G.P. Menthol Enhances the Desensitization of Human alpha3beta4 Nicotinic Acetylcholine Receptors. Mol. Pharm. 2015, 88, 256–264. [Google Scholar] [CrossRef]
- Ashoor, A.; Nordman, J.; Veltri, D.; Yang, K.-H.S.; Al Kury, L.; Shuba, Y.; Mahgoub, M.; Howarth, F.C.; Sadek, B.; Shehu, A.; et al. Menthol Binding and Inhibition of α7-Nicotinic Acetylcholine Receptors. PLoS ONE 2013, 8, e67674. [Google Scholar] [CrossRef] [Green Version]
- Ashoor, A.; Nordman, J.; Veltri, D.; Yang, K.-H.S.; Shuba, Y.; Al Kury, L.; Sadek, B.; Howarth, F.C.; Shehu, A.; Kabbani, N.; et al. Menthol Inhibits 5-HT3 Receptor–Mediated Currents. J. Pharmacol. Exp. Ther. 2013, 347, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Corvalan, N.A.; Zygadlo, J.A.; Garcia, D.A. Stereo-selective activity of menthol on GABA(A) receptor. Chirality 2009, 21, 525–530. [Google Scholar] [CrossRef]
- Hall, A.C.; Turcotte, C.M.; Betts, B.A.; Yeung, W.-Y.; Agyeman, A.S.; Burk, L.A. Modulation of human GABAA and glycine receptor currents by menthol and related monoterpenoids. Eur. J. Pharmacol. 2004, 506, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Sanjakdar, S.S.; Maldoon, P.P.; Marks, M.J.; Brunzell, D.H.; Maskos, U.; McIntosh, J.M.; Bowers, M.S.; Damaj, M.I. Differential roles of a6b2* and a4b2* neuronal nicotinic receptors in nicotine- and cocaine-conditioned reward in mice. Neuropsychopharmacology 2015, 40, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Yi, B.; Long, S.; González-Cestari, T.F.; Henderson, B.J.; Pavlovicz, R.E.; Werbovetz, K.; Li, C.; McKay, D.B. Discovery of benzamide analogs as negative allosteric modulators of human neuronal nicotinic receptors: Pharmacophore modeling and structure-activity relationship studies. Bioorganic. Med. Chem. 2013, 21, 4730–4743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.J.; Gonzalez-Cestari, T.F.; Yi, B.; Pavlovicz, R.E.; Boyd, R.T.; Li, C.; Bergmeier, S.C.; McKay, D.B. Defining the putative inhibitory site for a selective negative allosteric modulator of human a4b2 neuronal nicotinic receptors. ACS Chem. Neurosci. 2012, 3, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.J.; Orac, C.M.; Maciagiewicz, I.; Bergmeier, S.C.; McKay, D.B. 3D-QSAR and 3D-QSSR models of negative allosteric modulators facilitate the design of a novel selective antagonist of human a4b2 neuronal nicotinic acetylcholine receptors. Bioorg. Med. Chem. Lett. 2012, 22, 1797–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
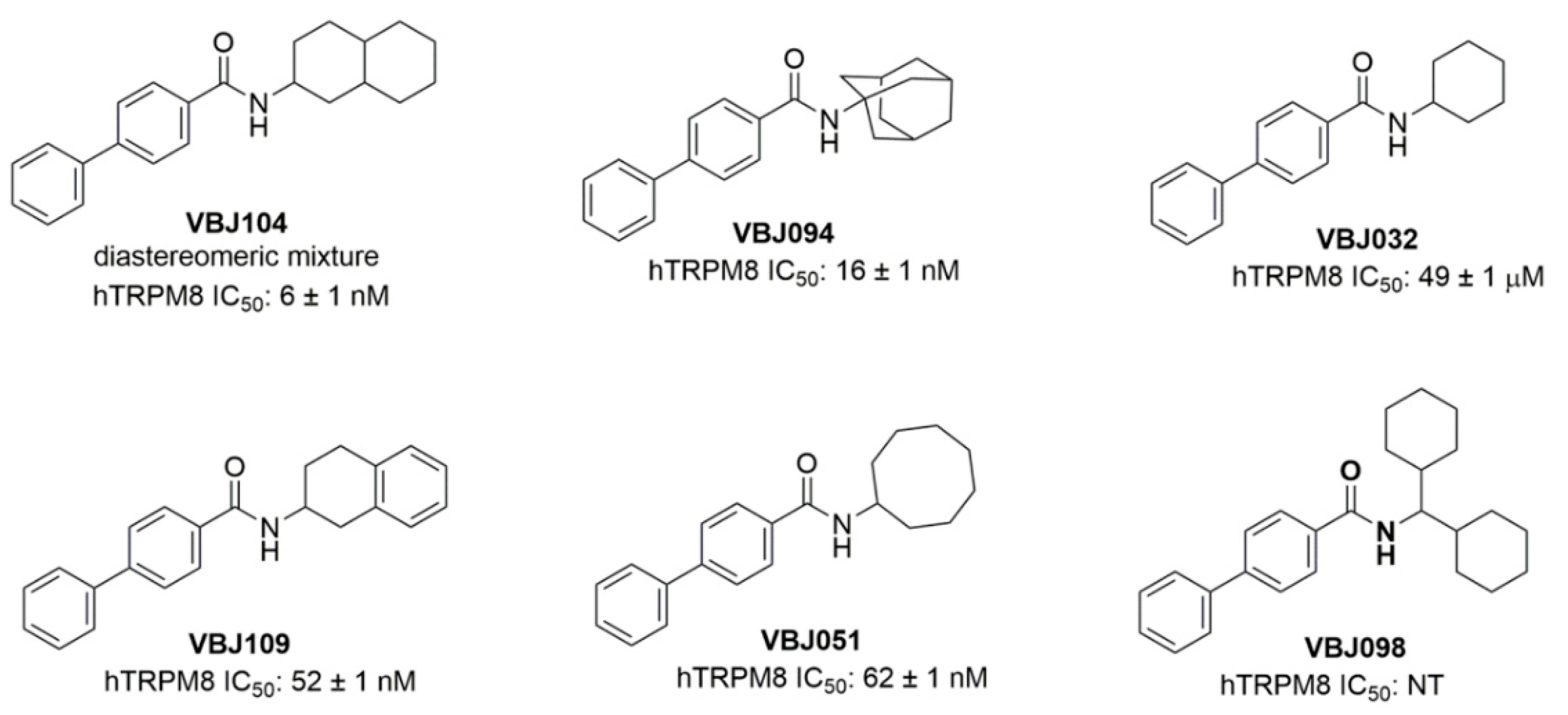




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | hTRPM8 IC50 a | α4β2 nAChR PM EC50 b | Max Efficacy (Normalized to 100 µM Nicotine) c |
|---|---|---|---|
| VBJ032 | 49 ± 1 µM | 77.6 nM | 158.5 ± 8.6% (50 µM) |
| VBJ051 | 52 ± 1 nM | 8.2 (0.8–47.2) nM | 143.6 ± 7.1% (50 µM) |
| VBJ094 | 16 ± 1 nM | 23.6 (1.8–68.2) nM | 130.7 ± 7.8% (30 µM) |
| VBJ098 | NE | NE | NE |
| VBJ104 | 6 ± 1 nM | 4.6 (2.6–8.4) µM | 360.7 ± 37.7% (100 µM) |
| VBJ109 | 52 ± 1 nM | 29.7 (4.3–69.0) nM | 125.6 ± 5.1% (50 µM) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooper, S.Y.; Akers, A.T.; Journigan, V.B.; Henderson, B.J. Novel Putative Positive Modulators of α4β2 nAChRs Potentiate Nicotine Reward-Related Behavior. Molecules 2021, 26, 4793. https://doi.org/10.3390/molecules26164793
Cooper SY, Akers AT, Journigan VB, Henderson BJ. Novel Putative Positive Modulators of α4β2 nAChRs Potentiate Nicotine Reward-Related Behavior. Molecules. 2021; 26(16):4793. https://doi.org/10.3390/molecules26164793
Chicago/Turabian StyleCooper, Skylar Y., Austin T. Akers, Velvet Blair Journigan, and Brandon J. Henderson. 2021. "Novel Putative Positive Modulators of α4β2 nAChRs Potentiate Nicotine Reward-Related Behavior" Molecules 26, no. 16: 4793. https://doi.org/10.3390/molecules26164793
APA StyleCooper, S. Y., Akers, A. T., Journigan, V. B., & Henderson, B. J. (2021). Novel Putative Positive Modulators of α4β2 nAChRs Potentiate Nicotine Reward-Related Behavior. Molecules, 26(16), 4793. https://doi.org/10.3390/molecules26164793