2.1. Effect of SAMul Ligands Surface Groups in Polyanion Recognition
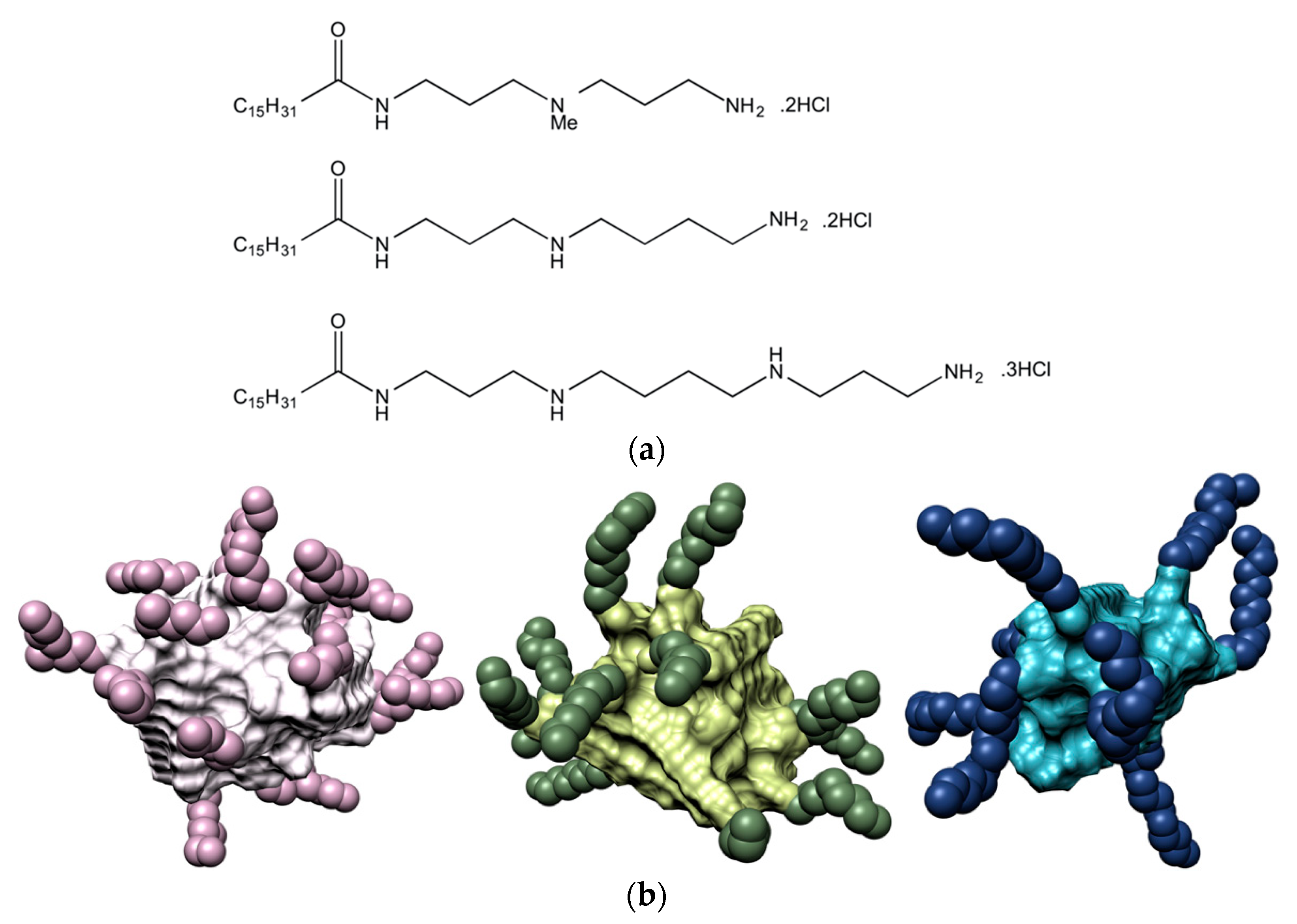
For the purpose of investigating the effect of the ligand head groups on polyanion recognition by SAMul ligands, we considered spermine (SPM), spermidine (SPD) and the shorter
N,
N-di-(3-aminopropyl)-
N-methylamine (DAPMA) as the polyanion-binding ligands [
13]. Mammalian cells naturally produce spermine and spermidine (along with their precursor putrescine), which play important roles in many cellular processes including regulation of transcription and translation, control of ion channels activity, modulation of kinase activities, effects on the cell cycle, protection from oxidative damage and maintenance of membrane structure/function. Above all, these two polyamines are known to physiologically interact with DNA in diverse and important ways, functioning as a protective agent, and a modulator of the nucleic acid secondary structure [
28]. In order to create the corresponding amphiphilic, self-assembling ligands, we initially selected palmitic acid (C
16) as the hydrophobic moiety, and connected the hydrophobe to the different amines via 2-(1H-benzotriazole-1-yl)-1,1,3,3-tetramethylaminium tetrafluoroborate (TBTU)-mediated peptide coupling with an appropriate group strategy [
13]. The synthesis yielded the three C
16-DAPMA, C
16-SPD, and C
16-SPM ligands, with nominal charge at pH = 7.4 of +2, +2, and +3, respectively (
Figure 1a). In aqueous solution at physiological pH and ionic strength (7.4, 150 mM NaCl), all three SAMul ligands were able self-assemble into almost spherical nanomicelles (
Figure 1b), whose main characteristics are listed in
Table 1.
The three different SAMul nanomicelles were next challenged for DNA and heparin binding. Experimentally, DNA binding was investigated via the ethidium bromide (EthBr) assay, during which the displacement of EthBr from DNA by each SAMul micelle is monitored by fluorimetry. Heparin binding was quantified via a heparin binding competition assay (HBCA) exploiting Mallard Blue (MB). MB is a highly charged (+5), blue-colored synthetic dye based on an arginine-functionalized thionine developed by our group as a specific heparin sensor. MB works in both physiological solutions and serum [
6,
7,
8,
10,
11,
12,
29,
30,
31,
32], and reports on heparin binding by UV−visible spectroscopy. Both displacement assays allow for the rapid determination of two binding parameters: CE
50 (i.e., the cation:anion charge excess at which 50% of each indicator dye is displaced from its complex) and EC
50 (i.e., the effective concentration at which 50% of each dye is displaced). Accordingly, the lower the values of CE
50 and EC
50 obtained wither in the MB or EthBr displacement assay, the stronger the affinity of the tested binder with respect to heparin and DNA, respectively.
In parallel, binding of the three nanomicelles to the two polyanions was investigated again experimentally - by Isothermal Titration Calorimetry (ITC)—and in silico, using atomistic molecular dynamics (MD) simulations. These two, more sophisticated techniques allow for the obtainment of the full thermodynamic characterization of each nanomicelle/polyanion interaction, i.e., the binding enthalpy ∆Hbind, binding entropy T∆Sbind, and free energy of binding ∆Gbind.
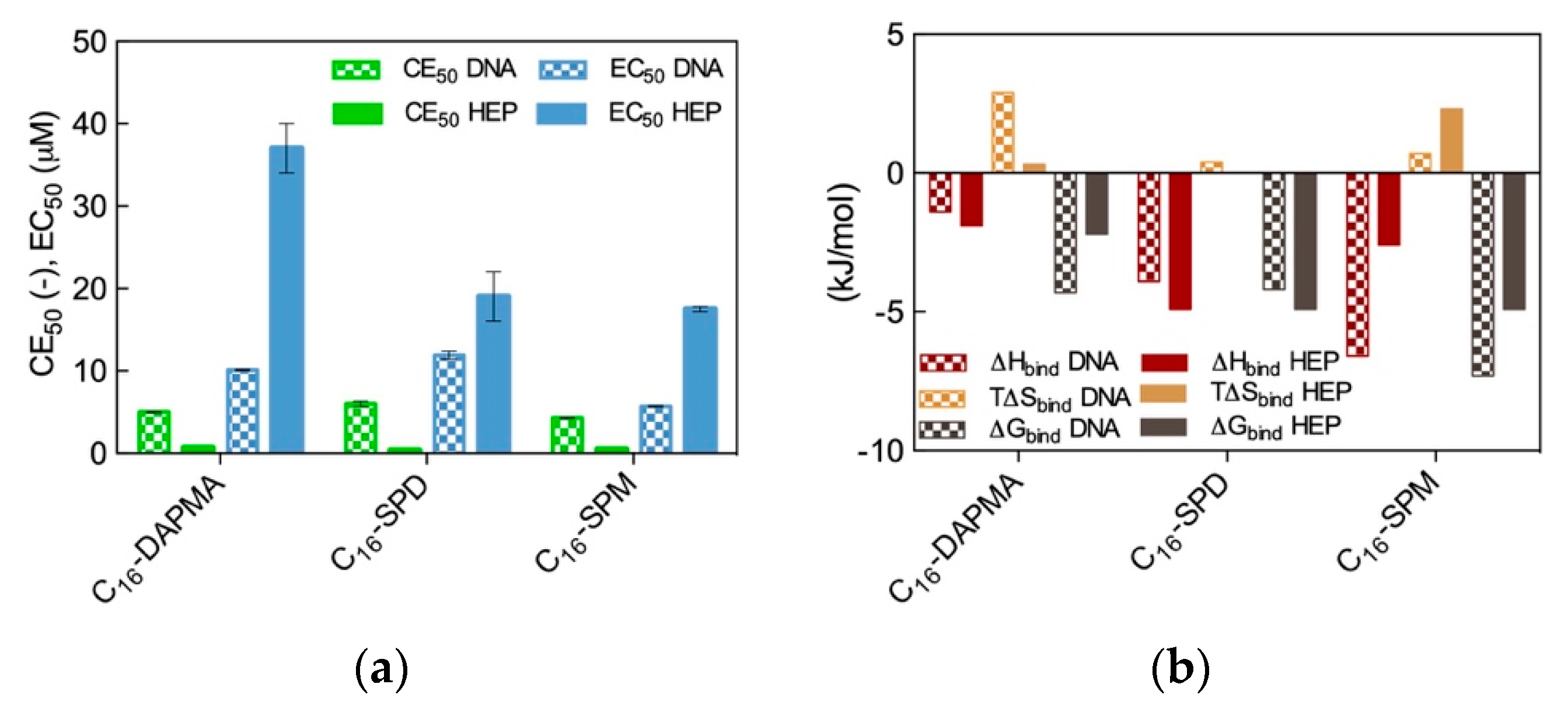
The results from the two spectrometric assays are shown in
Figure 2a, while the corresponding binding thermodynamics data are reported in
Figure 2b.
As seen from
Figure 2a, the preliminary results based on the dye displacement assays revealed that the micelles formed by the amphiphilic ligands featuring the most charged ligands (C
16-SPM) were optimized for DNA binding (CE
50 = 4.3), followed by the C
16-DAPMA (5.0) and the C
16-SPD (6.0) nanoassemblies, respectively. On the contrary, the same analysis showed that the nanomicelle/heparin binding charge-efficiency decreased in the order C
16-SPD (CE
50 = 0.34) > C
16-SPM (0.49) > C
16-DAPMA (0.69). Although these differences in CE
50 values might seem rather small, they were statistically meaningful and reproducible; as such, they provided a first clue on the fact that DNA and heparin can behave differently when facing these SAMul nanostructures as binding partners, even in the case when the nominal charge on the hydrophilic portion of the amphiphilic ligands was the same (e.g., +2 for both DAPMA and SPD).
We were somewhat taken aback by the apparent DNA/heparin selectivity difference discussed above as we had originally anticipated that both polyanion would bind the most highly charged micelles best or, at least, all three SAMul nanoobjects equally well on the basis of their predicted micellar surface density values (σ
mic = 0.026, 0.024, and 0.026 e/nm
2 for C
16-DAPMA, C
16-SPD, and C
16-SPM, respectively,
Table 1). Thus, we decided to verify this polyanion binding selectivity via a direct method (ITC). A comparison of the ITC-derived binding thermodynamics parameters (
Figure 2b) with the trends obtained from the displacement assays (
Figure 2a) revealed the broad agreement between the two data sets. Indeed, the micelles generated by the C
16-SPM SAMul ligands were again found to be the most effective DNA binders (∆G
bind = −7.3 kJ/mol), while the other two SAMul nanomicelles presented comparable affinity for the nucleic acid (∆G
bind = −4.3 kJ/mol and −4.2 kJ/mol for C
16-DAPMA and C
16-SPD, respectively). At the same time, ITC data consistently ranked the C
16-SPD micelles as the best heparin binders together with C
16-SPM (∆G
bind = −4.9 kJ/mol for both systems) and C
16-DAPMA as the SAMul nanostructures with lowest affinity towards this anionic polysaccharide (∆G
bind = −2.2 kJ/mol). When considering the per-charge-normalized binding free energy values (∆G
bind+), ITC data completely matched the dye displacement trends, particularly for heparin for which we obtained ∆G
bind+ = −2.45 kJ/mol for C
16-SPD, −1.63 kJ/mol for C
16-SPM, and −1.1 kJ/mol for C
16-DAPMA, respectively.
Having ascertained by transmission electron microscopy (TEM) that all cationic micelles remained intact upon polyanion binding, and that they were organized into hierarchical nanostructure arrays resulting in close packed interactions with both polyanions, we finally resorted again to atomistic MD simulations in order to provide an explanation for the displacement assays/ITC data and to get some insights into the binding interface between the three SAMul nanostructures and the negatively charged macromolecules. These results are shown in
Figure 3.
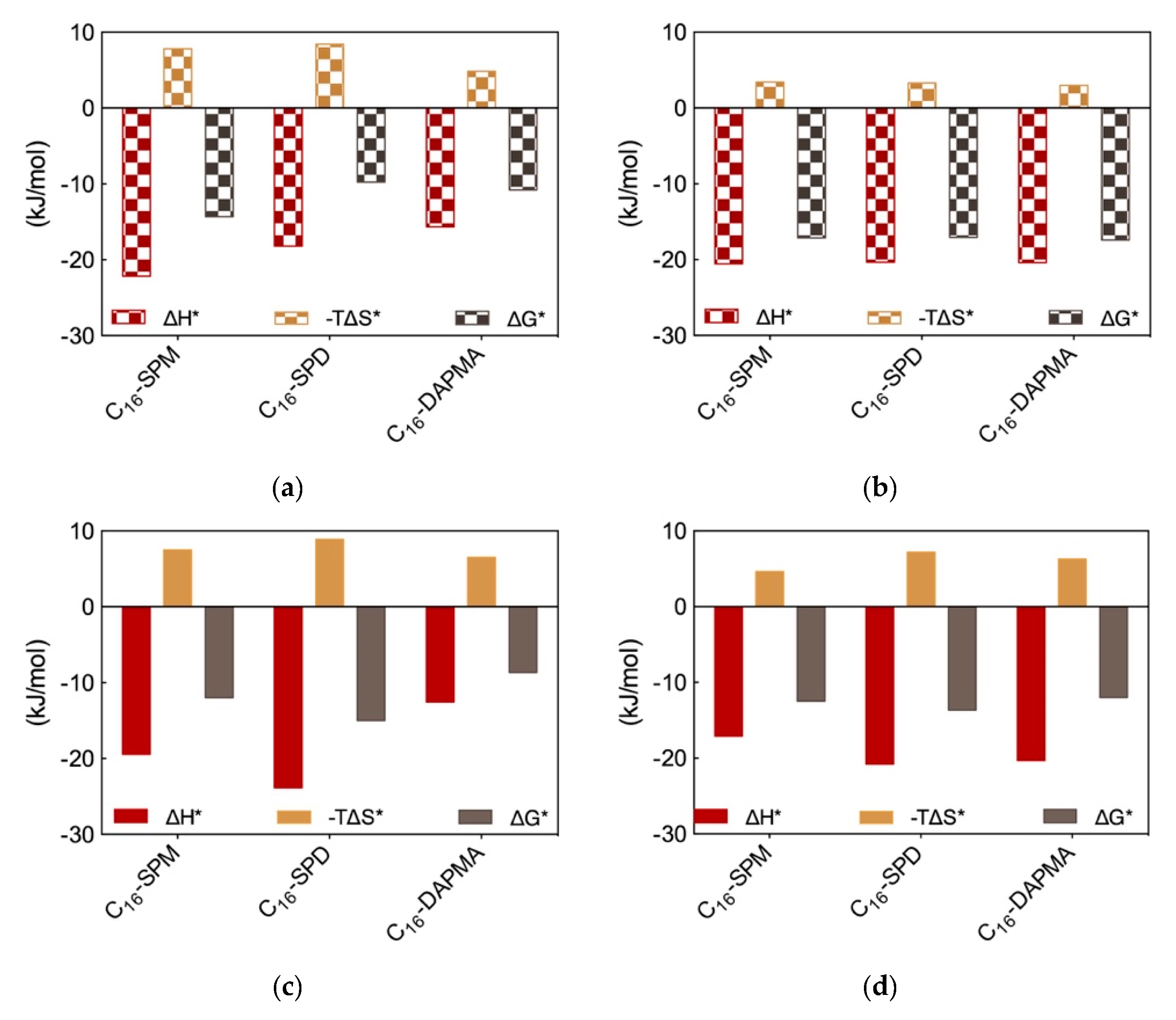
The MD trajectory analysis for each DNA/SAMul micelle complex performed from the viewpoint of the self-assembled ligands allowed us to observe that, during binding, the micelles formed by C
16-SPM contacted the nucleic acid with 9 out of 10 available SAMuls (i.e., the so-called effective residues), resulting in a favorable charge-normalized, per-effective-residue enthalpic gain ∆H*. This overcompensated the corresponding entropic cost (T∆S*) associated with ligand organization upon polyanion binding, leading to a favorable free energy of binding ∆G* of −14.32 kJ/mol (
Figure 3a), in agreement with ITC data (
Figure 2b). The other two micelles formed by the less-charged ligands (2+) both gained less in ∆H* (
Figure 3a), since the C
16-DAPMA and C
16-SPD nanostructures only used 7 and 8 effective residues (out of 16 and 13 available residues, respectively) to bind DNA. Yet, the micelles composed by the shorter, slightly more rigid ligand C
16-DAPMA payed less entropic penalty upon DNA interaction, and this slightly favored their nucleic acid affinity (∆G* = −10.80 kJ/mol) over C
16-SPD micelles (∆G* = −9.76 kJ/mol) (
Figure 3a), in line with the experimental ITC results (
Figure 2b). In aggregate, the simulated ∆G* values followed the same trend as the experimental CE
50 (
Figure 2a) and ITC values (
Figure 2b), with the DNA affinity of these SAMul micelles decreasing in the order C
16-SPM < C
16-DAPMA > C
16-SPD.
Applying the same analysis this time from the perspective of each anionic DNA residue implicated at the binding interface, the values of the three charge-normalized, per-effective-residue thermodynamic parameters ∆G*, ∆H* and T∆S* were essentially independent of the ligand—from the viewpoint of DNA, all interface interactions were equally good (
Figure 3b). This result led us to conclude that the selectivity of the SAMul micelles towards DNA can be ascribed only to ligand optimization—as such DNA appears to be a shape persistent polyanion which simply binds to, and organize the SAMul display with which it is presented.
For heparin binding, starting again from the perspective of the SAMul effective charge, the C
16-SPD micelles experienced the largest ∆H* gain upon reorganizing and optimizing the highest number of effective residues (12/13) while interacting with the polysaccharide with respect to the micelles formed by their other two counterparts C
16-DAPMA (9/16) and C
16-SPM (6/10) (
Figure 3c)—in line with the ITC data (
Figure 2b). Although the entropic loss for the C
16-SPD/heparin system was not the best in the series, the overall binding remained enthalpy-driven in nature, confirming the heparin best-binding properties of this SAMul nanosystem (∆G* = −14.98 kJ/mol) over the C
16-SPM (−11.97 kJ/mol) and C
16-DAPMA (−8.65 kJ/mol) ones (
Figure 3c), in keeping with both CE
50 (
Figure 2a) and ITC data (
Figure 2b). Considering SAMul/heparin binding from the perspective of each heparin sugar (
Figure 3d), a different behavior could also be observed, depending on the ligand nature: each heparin residue offset the entropic cost of binding C
16-SPD micelles with a greater enthalpic gain of its own. This was in stark contrast to DNA where, as discussed above, each anion behaved identically irrespective of the ligand. As such, the C
16-SPD nanomicelles induced more effective binding from each residue of the heparin chain via an enthalpy/entropy optimization, mediated through polyanion structural adaptation—i.e., heparin can be defined as an adaptive polyanion, which not only binds to the SAMul display, but importantly, is also able to adapt itself in response.
Thus, this part of our work highlighted the first misconception in ligand choice for generating selective SAMul displays at polyanion binding interfaces according to which polyanion and ligand charge density are the unique main players in driving electrostatic ion–ion binding—and introduced the two new concepts of ligand structural details and polyanion adaptability at binding interfaces as the two key parameters required for productive polyanion selectivity—as confirmed by the complementary experimental methods of competition binding assays, ITC and molecular simulations.
2.2. Effect of SAMul Hydrophobic Tails in Polyanion Recognition
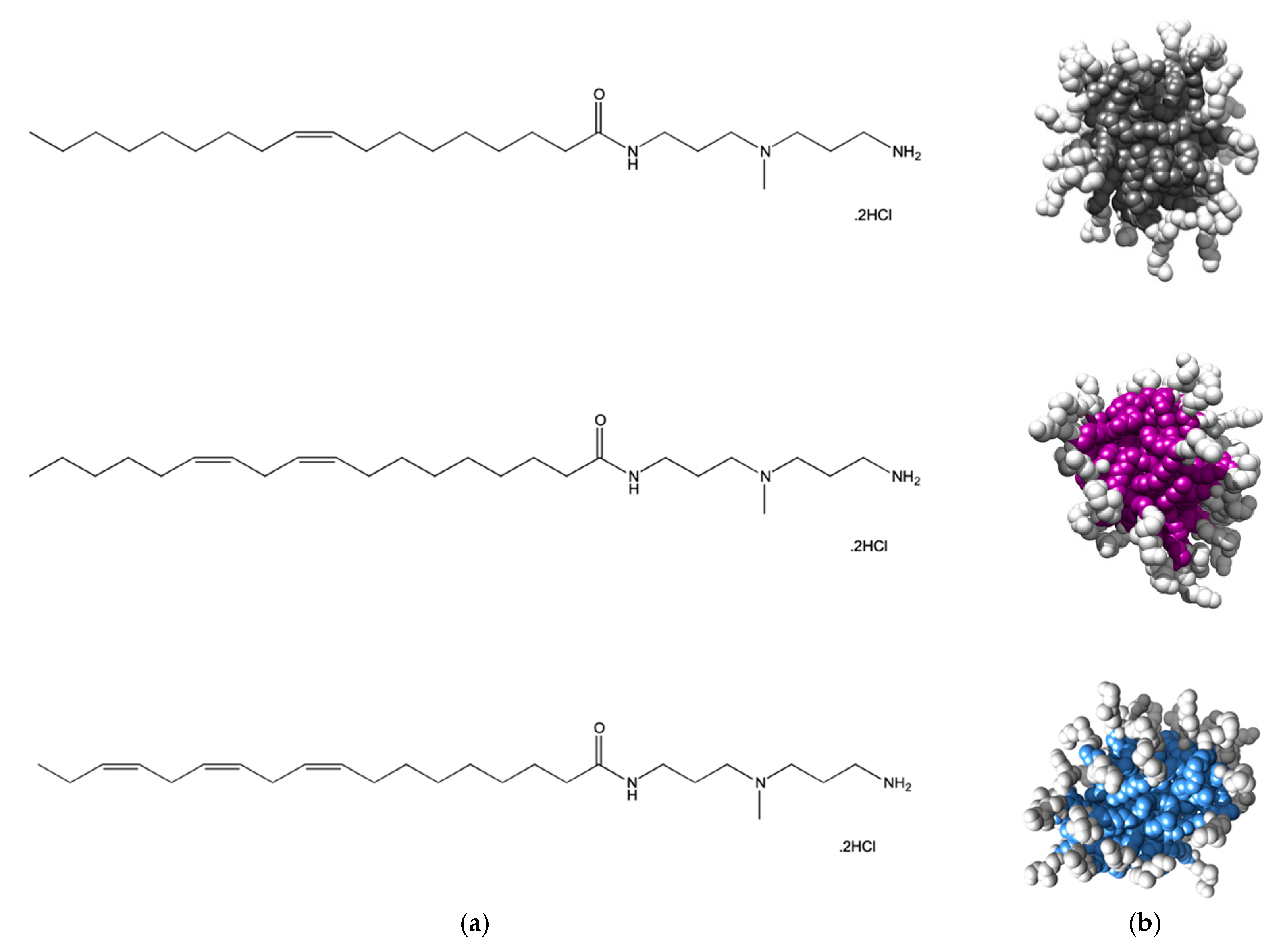
Although the hydrophobic moieties of amphiphilic molecules are responsible for driving self-assembly in aqueous environments, once the micellization process is completed they locate inside the resulting nanostructures and, as such, should not directly interfere with polyanion binding. Nonetheless, we decided to challenge the validity of this general concept and, since the self-assembled micelles of all three SAMul ligands C
16-DAPMA, C
16-SPD, and C
16-SPM were excellent systems for polyanion binding selectivity, we next decided to use closely-related ligands to explore the eventual role played by the hydrocarbon tails on their self-assembling properties and the related DNA/heparin binding [
12]. To the scope, we prepared three new molecular entities still featuring DAPMA as the polar head yet this time decorated with C
18 aliphatic chains bearing 1, 2 or 3 double bonds, respectively (see
Figure 4a). The reason underlying the choice of the C
18 instead of the C
16 fragment adopted in the study discussed above was solely due to the possibility of incorporating up to 3 double bonds in the hydrophobic portion while maintaining the identical and flexible 7-carbon-long segment within the same moiety.
Atomistic MD simulations initially predicted all compounds in
Figure 4 to self-assembly into spherical micelles of nanometric dimensions (
Figure 4b). For this nanomicelle set, the average micellar diameters (D
mic), aggregation number (N
agg) and zeta potential (ζ) values were computationally predicted to increase in passing from one to two C=C bonds in the alkyl chain while the presence of a further unsaturation resulted in a relatively confined additional effect (
Table 2). The subsequent experimental characterization of the relevant self-assembled forms fully confirmed the in silico data, as also shown in
Table 2.
Binding of the two polyanions with the nanomicelles formed by the three different SAMuls was next studied using again a combination of EthBr/MB displacement assays and MD simulations. Data the experimental techniques demonstrated that, for DNA, the SAMul micelle binding affinity decreased in the order C
18,3-DAPMA > C
18,2-DAPMA > C
18,1-DAPMA (CE
50 = 3.5, 4.3 and 5.0, respectively) while, for heparin, the opposite behavior was observed with polyanion binding ability decreasing with decreasing unsaturation level, i.e., C
18,1-DAPMA > C
18,2-DAPMA > C
18,3-DAPMA (CE
50 = 0.80, 1.8, ad 2.3, respectively) [
12]. Given the fact that the SAMul ligand head is the same for the three amphiphiles, and that the molecular-scale structural differences of these SAMul systems are buried in respective micellar core, this apparent polyanion selectivity was quite surprising. Thus, MD simulations were finally performed to explain this somewhat unanticipated findings (
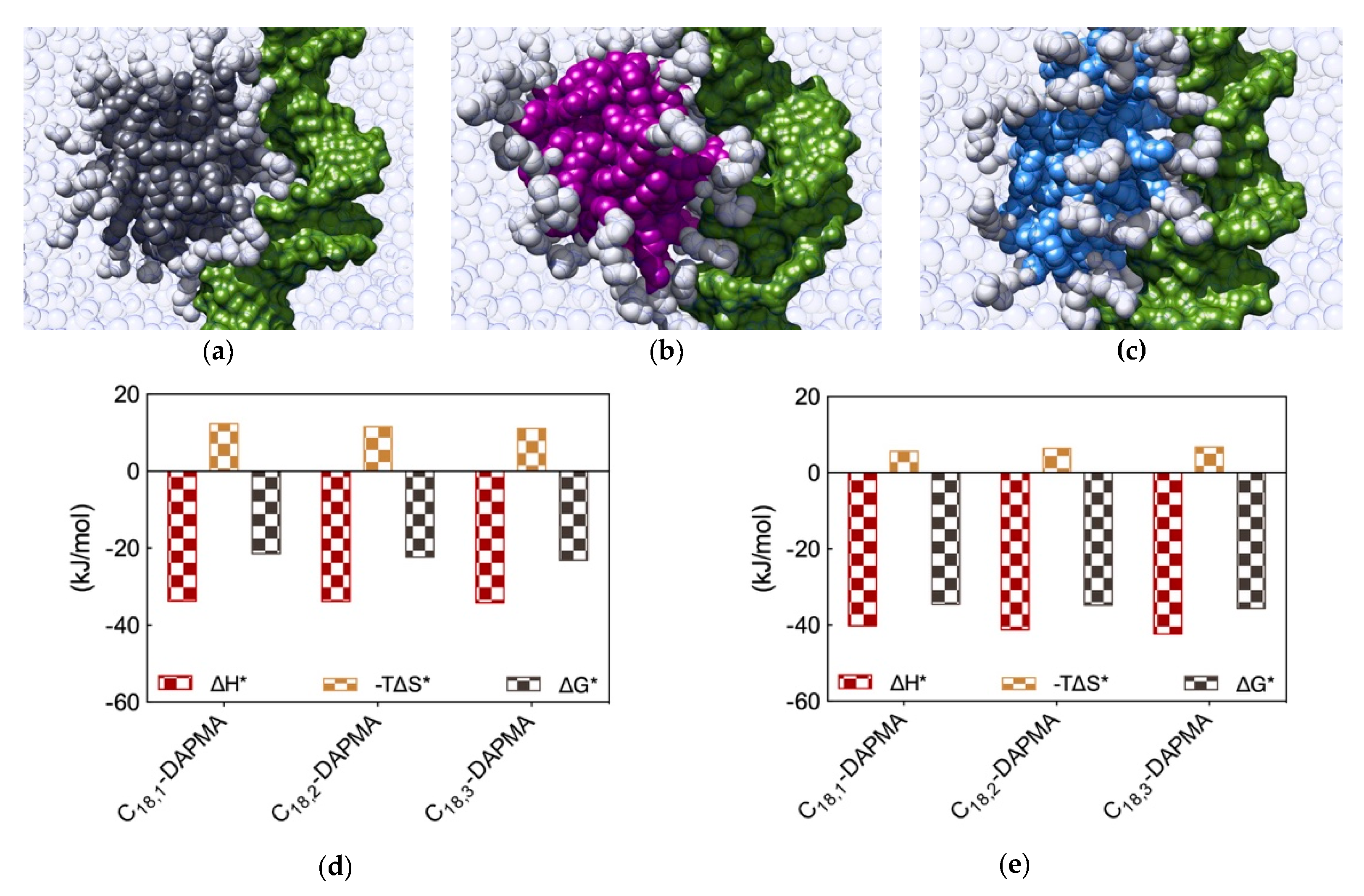
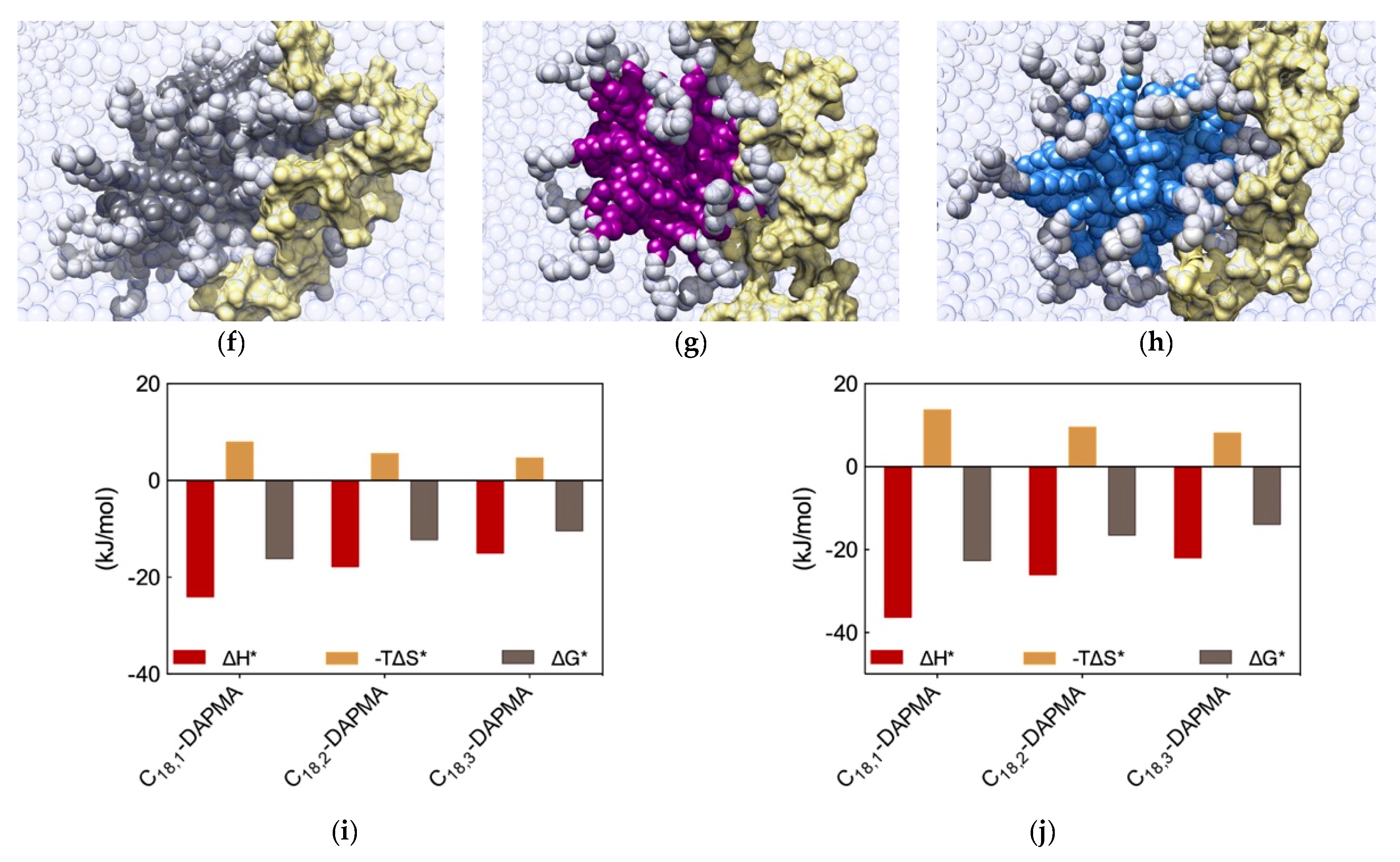
Figure 5a–c,f–h). A first, qualitative analysis of the MD data revealed that the micelles formed by C
18,1-DAPMA could exploit 19 effective residues (out of 28) in stably binding heparin whereas a progressive decreased in the number of effective residues was estimated when considering C
18,2-DAPMA (15/32) and C
18,3-DAPMA (13/35) in complex with the anionic polysaccharide, respectively. When bound to DNA, however, this difference in effective residues for each micelle type leveled off, being equal to 16, 17, and 18 for ligands bearing 1, 2, and 3 unsaturation in their aliphatic portion, respectively.
Further analysis of each SAMul/polyanion complex performed as detailed in
Section 2.1 indeed revealed the molecular reasons for these findings. Starting the discussion with heparin, and again considering polyanion binding from the viewpoint of each single effective SAMul interaction (
Figure 5d), the more flexible nature of the mono-unsaturated chain of C
18,1-DAPMA allow this SAMul micelles to maximize their interactions with the polysaccharide, resulting in the most favorable enthalpic contribution (∆H* = −24.02 kJ/mol), which overcompensate the unfavorable penalty paid upon binding (T∆S* = −7.92 kJ/mol) and ultimately leading to the largely favorable ∆G* value of −16.10 kJ/mol (
Figure 5d). As discussed a few lines above, increasing the rigidity of the hydrocarbon chain has the effect of decreasing the number of micelle effective residues involved in heparin productive binding, and this reflects into a progressive decrement of the corresponding binding enthalpic component (∆H* = −17.76 and −14.98 kJ/mol for C
18,2-DAPMA and C
18,3-DAPMA, respectively). However, the inclusion of 2 or 3 double carbon-carbon bonds in the hydrophobic moieties of the SAMul ligands has a beneficial effect on the corresponding binding entropy (T∆S* = −5.55 and −4.63 kJ/mol for C
18,2-DAPMA and C
18,3-DAPMA, respectively) since, as these molecules become progressively more rigid, they are less subjected to conformational reorganization upon binding and, as such, also suffer less entropic penalty. As a net result, the heparin binding of these two SAMul micelles however remain enthalpically driven (the corresponding ∆G* being equal to −12.21 and −10.35 kJ/mol for C
18,2-DAPMA and C
18,3-DAPMA, respectively).
Overall, the affinity of the three SAMul micelles for heparin, as expressed by the corresponding simulated values of ∆G*, indeed follow the same trend as the corresponding CE50 values, that is C18,1-DAPMA > C18,2-DAPMA > C18,3-DAPMA.
When considering DNA binding, simulation results highlight an interesting feature: the enthalpic gain for the three different SAMul micelles upon interaction with the nucleic acid is almost constant and equal to ∆H* = −33.77, −33.81, and −34.15 kJ/mol for C
18,1-DAPMA, C
18,2-DAPMA, and C
18,3-DAPMA, respectively (
Figure 5e). Thus, in absolute terms, the small difference in effective binding enthalpy between the micelles formed by the least and the most unsaturated SAMul micelle (0.38 kJ/mol) is in stark contrast with what observed in the case of heparin binding, where the same difference amounts to 9.04 kJ/mol (
Figure 5d). Also, the entropic component is decidedly less subjected to variation when these three micelles are placed in contact with DNA, the difference between C
18,1-DAPMA and C
18,3-DAPMA being again limited to 1.3 kJ/mol (
Figure 5e) while, for heparin, it is almost this value is almost the double (3.3 kJ/mol,
Figure 5d). As a result, the values of DG* for the DNA binding by these three SAMul micelles are definitely less sensitive to each SAMul molecular structure and, in agreement with their corresponding experimental CE
50 values, they rank the affinity of the micelles for the polyanion in the order C
18,3-DAPMA > C
18,2-DAPMA > C
18,1-DAPMA (∆G* = −23.07, −22.27, and −21.43 kJ/mol, respectively,
Figure 5e).
The same analysis performed on the heparin perspective showed that both the heparin sugars and the micellar ligands experienced the same trend in the variation of the three binding components (
Figure 5i). Accordingly, upon binding the micelles generated by the most flexible ligand (C
18,1-DAPMA) the polyanion can cope with a higher entropy loss via a considerably more favorable enthalpic stabilization. At the same time, this polyanion is able to gradually adjust the enthalpy/entropy compensation as the micellar rigidity increases, a phenomenon utterly analogous to that observed when studying the effect of the ligand head effect on polyanion selectivity discussed in
Section 2.1. Once again, these data confirm the concept that heparin is a relatively flexible adaptive polyanion. From the DNA viewpoint, the values of ∆G* and its components ∆H* and T∆S* are aging largely independent of the hydrophobic portion of the SAMul ligands (
Figure 5j), confirming that each DNA units binds its cationic counterpart with comparable strength. In agreement with what presented in
Section 2.1, DNA confirms to be a relatively rigid and shape-persistent polyanion.
Thus, the new concept we learned from this study is that, contrarily to an intuitive misconception, even though the molecular structural differences in these new three ligands are buried in the hydrophobic cores of the micelles they generate, with a mechanisms prototypical of SAMul materials these characteristics are transmitted through the entire nanoobjects, ultimately resulting into significantly different polyanion binding preferences, with heparin—an adaptive polyanion—being more affine to the micelles constituted by the most flexible monomers.
2.3. Effect of SAMul Chirality in Polyanion Recognition
Chiral recognition by DNA and heparin is not a new concept. For instance, the chiral discriminating capabilities of heparin have been exploited in capillary electrophoresis [
33] for the separation of several chiral drugs [
34]. Contextually, DNA is an inherently chiral molecule, as its constituents contain several stereogenic centers. As a matter of fact, DNA global handedness is responsible for the nucleic acid recognition by chiral molecules and, indeed, DNA has been exploited as a very efficient chiral stationary phase [
35] and as a chiral microenvironment or chiral template in asymmetric synthesis [
36].
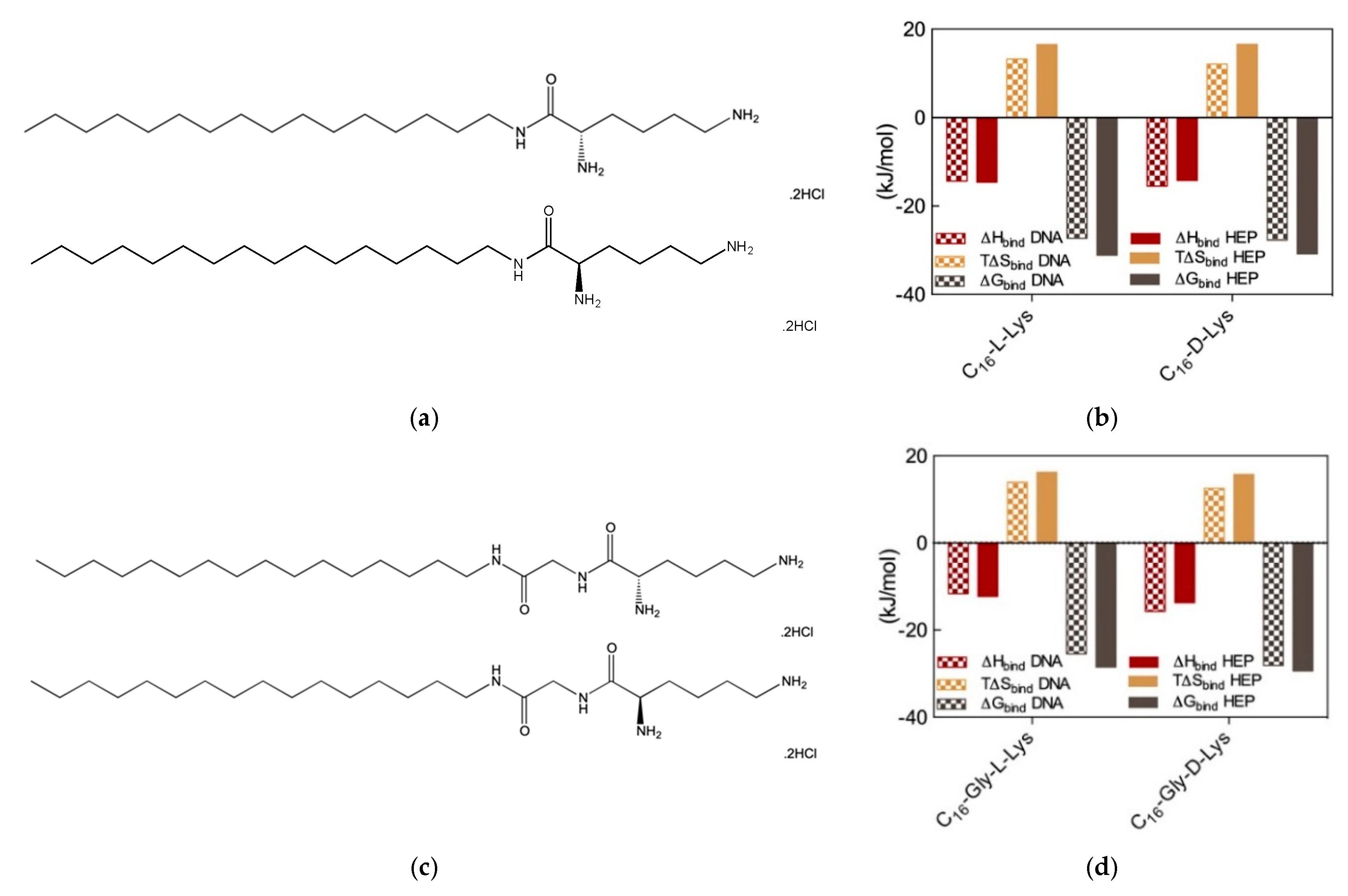
Therefore, we wondered whether chiral SAMul systems could achieve enantioselectivity in the two polyanion binding. To the purpose, we initially synthesized two new amphiphilic cationic ligands C
16-
l-Lys and C
16-
d-Lys (
Figure 6a), which are utterly identical apart from the chirality of the lysine ligands [
14]. As expected, these two chiral SAMul molecules self-assembled – at the same CMC (45 and 48 μM for the
l and
d SAMul, respectively)—into spherical nanomicelles with similar dimensions (D
mic = 6.2 ± 1.7 and 6.3 ± 1.7 nm) and surface charges (ζ = + 45.2 ± 1.6 and 39.2 ± 1.6 mV) but opposed chirality. It was therefore not surprising to find that these two nanostructures failed to show any preference in binding heparin and DNA, the corresponding CE
50 values being equal to 1.8 ± 0.1 and 1.8 ± 0.1 for heparin binding and to 1.6 ± 0.2 and 1.7 ± 0.1 for DNA binding by C
16-
l-Lys and C
16-
d-Lys, respectively. The results from MB and EthBr displacement assays were definitively confirmed by ITC (
Figure 6b), which confirmed the absence of chiral recognition in the polyanion binding thermodynamics by the two SAMul nanomicelles (e.g., for DNA ∆G
bind = −27.3 and −27.7 kK/mol and for heparin ∆G
bind = −31.1 and −30.8 kJ/mol for C
16-
l-Lys and C
16-
d-Lys, respectively).
Although these results suggested that self-assembled nanoscale chirality has no significant impact on the molecular recognition interfaces, we were still not satisfied and went on by synthesizing two closely related molecules, C
16-Gly-
l-Lys and C
16-Gly-
d-Lys, featuring a glycine spacer between the C
16 hydrophobic chain and the L/D lysine polar head of the ligand (
Figure 6c). The self-assembly of these two new chiral SAMuls also resulted in small and spherical micelles with similar characteristics (e.g., CMC = 49 and 49 μM and ζ = 40.1 ± 0.1 and 47.1 ± 0.1 mV for C
16-Gly-
l-Lys and C
16-Gly-
d-Lys, respectively). Pleasingly, however, the results from the two, preliminary dye-based assays revealed that chiral discrimination at the nanomicelle-polyanion binding interface has been switched on by the presence of the glycine spacer unit. In fact, the EthBr displacement assay indicated that the DNA binding ability of C
16-Gly-
l-Lys and C
16-Gly-
d-Lys was significantly different, the corresponding CE
50 values being equal to 3.8 ± 0.7 and 1.5 ± 0.1, respectively. At the same time, also with respect to heparin the two new chiral SAMuls displayed different performances, with CE
50 values of 1.7 ± 0.2 and 1.1 ± 0.1 or C
16-Gly-
l-Lys and C
16-Gly-
d-Lys, respectively. The chiral discrimination of the two polyanions by these SAMul nanostructures was again confirmed by ITC experiments (
Figure 6d). The relevant binding thermodynamic parameters indeed showed that DNA displays a clear preference for the micelles formed by the SAMul D-enantiomer over those generated by the amphiphilic ligand with opposed chirality, with ∆G
bind values of −28.1 and −25.5 kJ/mol for C
16-Gly-
d-Lys and C
16-Gly-
l-Lys, respectively. Furthermore, the same techniques indicated that also heparin binds the D-SAMul micelles with a slight preference (∆G
bind = −29.4 kJ/mol) over the L-SAMul ones (∆G
bind = −28.5 kJ/mol). A more detailed analysis of the polyanion binding thermodynamics by these two chiral SAMul nanosystems showed that the binding enthalpy is always negative (i.e., exothermic binding), as it could be expected from interaction driven by electrostatic forces. The ∆H
bind values for DNA binding are −15.7 and −11.6 kJ/mol while those for heparin binding are −13.7 and 12.3 kJ/mol for C
16-Gly-
d-Lys and C
16-Gly-
l-Lys, respectively. The binding entropies are also favorable (i.e., positive,
Figure 6), suggesting the combined effect of some stabilizing hydrophobic interactions between the CH
2-group of the micelle terminal Lys moieties and the bases/sugars of the polyanions and an increment in the degree of disorder of the overall system induced by the release of water molecules and counterions upon nanoscale binding interface formation. Interestingly, however, the entropic differences between the enantiomeric micelles are less pronounced, as for DNA the T∆S
bind values are equal to +15.7 and +16.2 kJ/mol and for heparin these quantities amount to +12.5 and +13.9 kJ/mol for C
16-Gly-
d-Lys and C
16-Gly-
l-Lys, respectively. In aggregate, these results clearly show that the enhanced DNA binding—and, albeit to a lesser extent, heparin binding—of the C
16-Gly-
d-Lys-based nanomicelles is an enthalpically-driven process. As such, the specific SAMul-polyanion recognition appears to be optimized on the surface of the C
16-Gly-
d-Lys micelles in comparison with the otherwise identical nanoassemblies formed by the C
16-Gly-
l-Lys amphiphilic ligands. We also reasoned that the lower degree of heparin recognition exhibited by these two micelles could be the results of the greater heparin polydispersity, which ultimately reflects into a less-well defined distribution of the anionic sites along the polysaccharide backbone. DNA, as a more rigid and less-adaptive polyanion (
Section 2.1), is able to present its anionic sited evenly and more regularly spaced down its helical structure, thereby likely benefitting more from a suitably structured binding counterpart.
In summary, the further new concept we learned from this study is that, contrarily to another intuitive misconception, chiral discrimination and differential polyanion recognition by otherwise identical nanomicelles can be switched on by the presence of specific molecular features—such as an apt spacer connecting the hydrophobic and hydrophilic portions of the micellar amphiphilic constituents—which ultimately results in the differential optimization of the relevant binding nanointerfaces. Understanding such effects in details can add important criteria for the design of new SAMul ligands with enhanced chiral recognition to be generally exploited in optimizing binding process at self-assembled bio-interfaces (e.g., cell membranes, proteins and other polyanions).
2.4. Effect of SAMul Chirality in Polyanion Recognition—Revisited
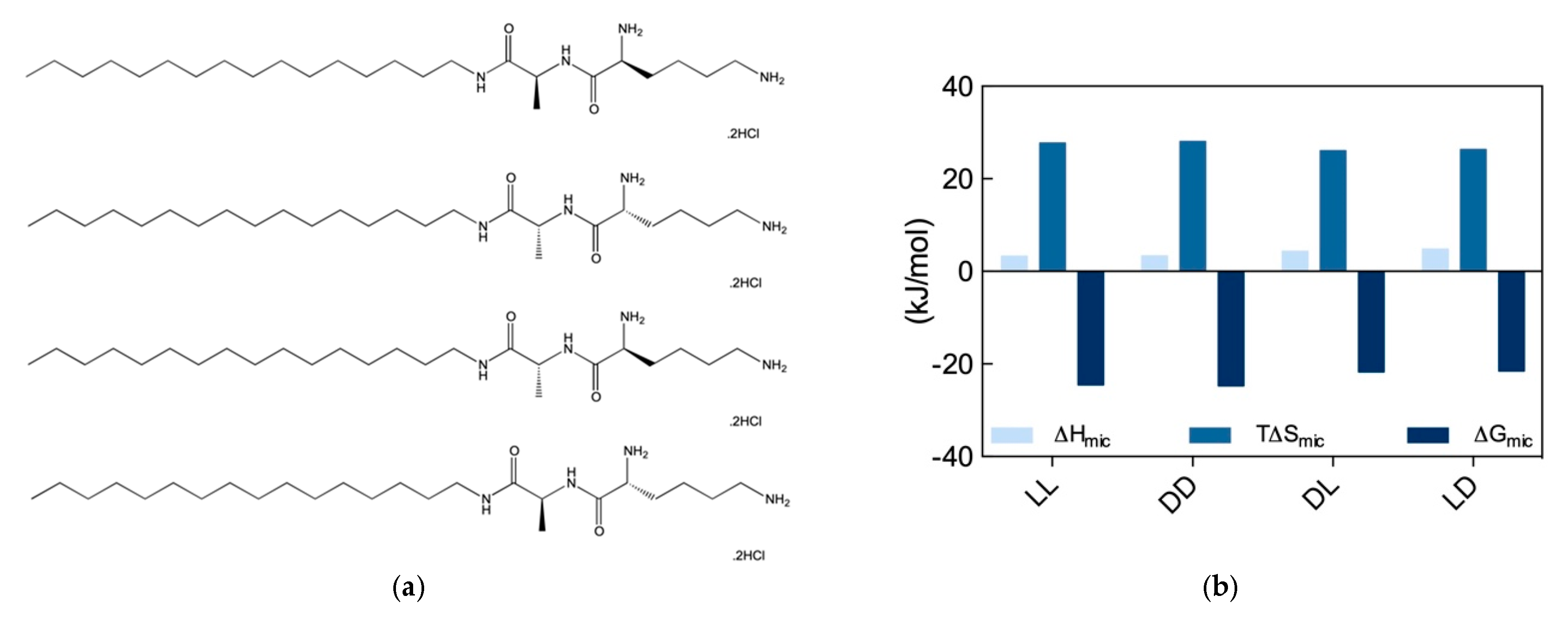
Given the novelty and the practical application potential of the results discussed in
Section 2.3, we decided to gain a deeper understanding of the recognition potential of chiral SAMul systems towards the two prototypical biological polyanions, DNA and heparin. To the purpose, another set of stereoisomeric SAMul amphiphiles were designed and synthesized, as shown in
Figure 7a. By virtue of the presence of the two amino acids Ala and Lys, each of these molecules contains two chiral centers; accordingly, they can exist in four possible stereoisomers, i.e., two enantiomer pairs with a diastereomeric relationship to each other, as follows: C
16-
l-Ala-
l-Lys and C
16-
d-Ala-
d-Lys (
ll and
dd), and C
16-
d-Ala-
l-Lys and C
16-
l-Ala-
d-lys (
dl and
ld), respectively [
8]. We reasoned that, in contrast to our previous work discussed above, the presence of a second chiral center (Ala) that is not directly involved in the binding interface, should allow us to explore both enantiomeric and diastereomeric effects on polyanion binding selectivity.
The self-assembly of these four new SAMul ligands was initially explored by ITC, according to which the
ll and
dd nanosystems were characterized by a substantially lower CMC (52 and 48 μM for the
ll and
dd, respectively) than the
dl and
ld ones (159 and 172 μM, respectively). Interesting, however, at variance with what observed in our previous work (
Section 2.3), for these new amphiphiles the micellization process was entropically driven (T∆S
mic > 0) and slightly enthalpically disfavored (∆H
mic > 0) (
Figure 7b). In particular, the analysis of the micellization thermodynamic data reported in
Figure 7b reveals that micelle formation by the
ll and
dd SAMuls are both slightly enthalpically and entropically preferred over the micellization of the diastereomeric
dl and
ld SAMuls. Accordingly, the relevant ∆G
mic values becomes more favorable, in the order:
ll (∆G
mic = −24.5 kJ/mol) ≈
dd (∆G
mic = −24.7 kJ/mol) >
dl (∆G
mic = −21.7 kJ/mol) ≈
ld (∆G
mic = −21.5 kJ/mol). The self-assembly of the four SAMuls in
Figure 7a was also monitored by CD (above the respective CMC), which indicated that the nanoassemblies had equal and opposite chirality. However, DLS measurements suggested that the diastereoisomers formed slightly different spherical micelles, the
ll and
dd assembling into smaller, better defined aggregates with lower surface charge (D
mic = 6.5 ± 2.9 and 7.2 ± 2.2 nm and ζ = 35.5 ± 3.3, 39.2 ± 2.2 mV) with respect to
dl/
ld (D
mic = 9.2 ± 2.8 and 8.6 ± 1.6 nm and ζ = 46.8 ± 0.5, and 43.3 ± 0.6 mV).
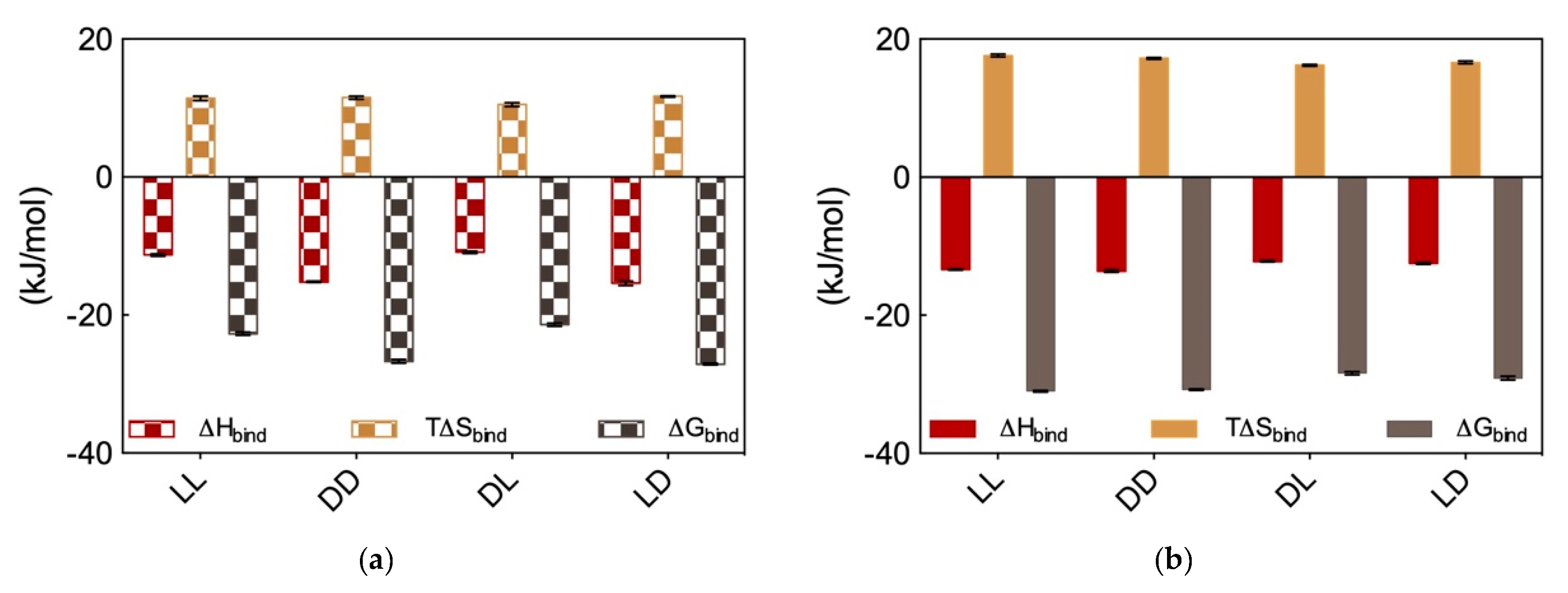
In the light of these somewhat unexpected results, we proceeded with performing DNA and heparin binding via ITC. These results are reported in
Figure 8a and
Figure 8b, respectively. Considering DNA binding first, data in
Figure 8a show that the
dd and
ld micelles displayed higher DNA affinity (∆G
bind = −26.7 ± 0.3 and −27.1 ± 0.1 kJ/mol, respectively) with respect to those formed by the alternative couple
ll and
dl (∆G
bind = −22.7 ± 0.2 and −21.4 ± 0.1 kJ/mol, respectively). Overall, the affinity for DNA of these SAMul micelles decreases in the order
ld >
dd >
ll >
dl. Importantly, the difference in ∆G
bind between the DNA best binder (
ld) and the micelles presenting the lowest affinity for this polyanion (
dl) is large (5.7 kJ/mol), and substantially larger than that measured for the related systems discussed above (
Section 2.3), for which this difference amount to 2.6 kJ/mol. Also, further analysis of the thermodynamic data in
Figure 8a reveals that enthalpy is governing the interaction between these four micelle types and DNA (∆H
bind = −15.4 ± 0.3, −15.2 ± 0.1, −11.3 ± 0.2, and −10.9 ± 0.2 for
ld, dd, ll, and
dl, respectively) although also T∆S
bind affords a favorable (i.e., positive) yet almost constant contribution to the interaction (T∆S
bind = +11.7 ± 0.1, +11.5 ± 0.2, +11.4 ± 0.3, and +10.5 ± 0.3 for
ld, dd, ll, and
dl, respectively). This last result is in line with the shape persistent nature of DNA discussed in
Section 2.1, according to which this polyanion is better able to optimize interactions for enthalpic gain by virtue of its well-organized and repetitive structure along its double helix.
Moving on to heparin binding, from the ITC data reported in
Figure 8b it appears immediately that not only all these four chiral SAMul micelles are able to bind the polysaccharide better than DNA but also, and perhaps more importantly, they show substantially less polyanion chiral recognition. Quantitatively, the ∆G
bind values for heparin binding by these nanomicelles are the following: −31.1 ± 0.1, −30.8 ± 0.1, −28.4 ± 0.2, and −29.1 ± 0.3 kJ/mol for
ll, dd; dl, and
ld, respectively. Therefore (although in this case with a considerably lower difference between the “stronger” and “weaker” binder then in the case of DNA), the affinity for the polysaccharide by these SAMuls follows the order:
ll ≥
dd >
ld ≥
dl. As it is also evident from data in
Figure 8b, and contrarily to what observed for DNA, heparin binding is entropically driven (T∆S
bind = +17.6 ± 0.2, +17.2 ± 0.1, +16.2 ± 0.1, and +16.6 ± 0.2 kJ/mol for LL, DD; DL, and LD, respectively), although also in this case enthalpy affords in all cases a favorable contribution (∆H
bind = −13.4 ± 0.1, −13.6 ± 0.2, −12.2 ± 0.1, and −12.5 ± 0.01 kJ/mol for
ll,
dd;
dl, and
ld, respectively). This is also in line with the adaptive character of the heparin polyanion (
Section 2.1) which, with respect to the shape-persistent DNA, can re-organize its conformation to achieve a greater surface contact with the SAMul nanomicelles. This, in turn, leads to a more effective binding interface stabilized by–besides the expected electrostatic interactions (∆H
bind < 0)—hydrophobic contacts between the Lys CH
2 groups and, above all, a substantial release of water molecules and counterions upon nanoscale binding interface formation (T∆S
bind > 0).
In aggregate, these results are quite important, as they led us to formulate two major concepts: (i) with respect to Gly, the presence of the chiral spacer Ala in the inner part of the SAMul ligand head assists the terminal chiral Lys residue in its pre-organization for DNA enantiomeric recognition, and (ii) while the stereochemical configuration of the inner spacer in the SAMul ligand head does not play a role in DNA recognition, that at the chiral center of the terminal binding unit does, as micelles featuring SAMul the terminal binding unit in the D configuration (e.g., ld or dd) are considerably more affine to DNA then those having the surface Lys with opposite chirality (ll or dl). On the contrary, the ability of adaptive, ill-defined heparin binding is primarily controlled by the ability of the SAMul systems to self-assemble irrespective of the chirality presented at the binding interface, with heparin wrapping around the nanosystems but not forming highly optimized electrostatic interaction with the chiral nanomicelles.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}