Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis



Abstract
1. Introduction
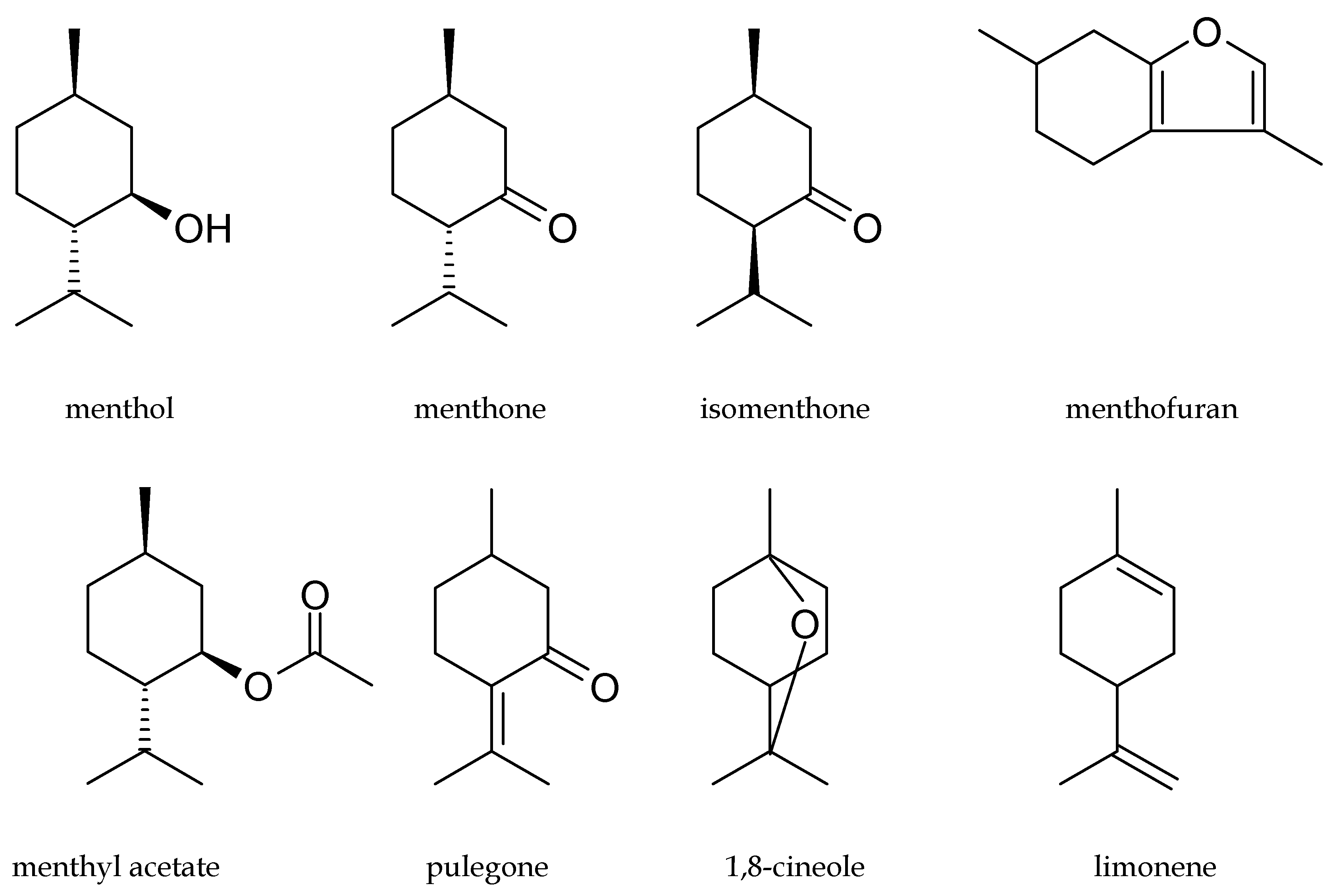
2. Content and Chemical Composition of Peppermint Oil and Cornmint Oil
3. Biological Activity and Application of Peppermint Oil and Cornmint Oil
4. Antifungal and Antibacterial Activity of Peppermint Oil and Cornmint Oil against Phytopathogens
5. Peppermint Oil and Cornmint Oil for Postharvest Decay Control
6. Insecticidal and Acaricidal Activity of Peppermint Oil and Cornmint Oil
6.1. Against Storage Insects
6.2. Against Herbivory Insects
6.3. Against Livestock Insects
6.4. Acaricidal Effects
7. Peppermint and Cornmint EOs as Candidates for Botanical Herbicides
8. Formulations of Menthol Mint Essential Oils in Agriculture—Limitations and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharopov, F.; Antolak, H.; Kręgiel, D.; Sen, S.; Sharifi-Rad, M.; Acharya, K.; Sharifi-Rad, R.; et al. Plants of Genus Mentha: From Farm to Food Factory. Plants 2018, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P. Recent advances and challenges in trichome research and essential oil biosynthesis in Mentha arvensis L. Ind. Crop. Prod. 2016, 82, 141–148. [Google Scholar] [CrossRef]
- Pushpangadan, P.; Tewari, S.K. Peppermint. In Handbook of Herbs and Spices; Elsevier: Amsterdam, The Netherlands, 2006; pp. 460–481. [Google Scholar]
- Bacon, F.J. The Botanical Origin of American Peppermint—Mentha Piperita L*. J. Am. Pharm. Assoc. (1912) 1928, 17, 1094–1096. [Google Scholar] [CrossRef]
- Maffei, M.; Sacco, T. Chemical and Morphometrical Comparison Between two Peppermint Notomorphs. Planta Medica 1987, 53, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Grzeszczuk, M.; Jadczak, D. Estimation of biological value of some species of mint (Mentha L.). Herba Polonica. 2009, 55, 193–199. [Google Scholar]
- Machiani, M.A.; Javanmard, A.; Morshedloo, M.R.; Maggi, F. Evaluation of competition, essential oil quality and quantity of peppermint intercropped with soybean. Ind. Crop. Prod. 2018, 111, 743–754. [Google Scholar] [CrossRef]
- Machiani, M.A.; Javanmard, A.; Morshedloo, M.R.; Maggi, F. Evaluation of yield, essential oil content and compositions of peppermint (Mentha piperita L.) intercropped with faba bean (Vicia faba L.). J. Clean. Prod. 2018, 171, 529–537. [Google Scholar] [CrossRef]
- Ulbrich, A.; Kahle, H.; Kramer, P.; Schulz, M. Mentha x piperita volatiles promote Brassica oleracea-A pilot study for sustainable vegetable production. Allelopath. J. 2018, 43, 93–104. [Google Scholar] [CrossRef]
- Karkanis, A.; Alexiou, A.; Katsaros, C.; Petropoulos, S. Allelopathic Activity of Spearmint (Mentha spicata L.) and Peppermint (Mentha × piperita L.) Reduces Yield, Growth, and Photosynthetic Rate in a Succeeding Crop of Maize (Zea mays L.). Agron. 2019, 9, 461. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kimbaris, A.; Michaelakis, Α.; Papachristos, D.; Polissiou, M.; Papatsakona, P.; Tsora, E. Insecticidal Activity of Plant Essential Oils Against the Vine Mealybug, Planococcus ficus. J. Insect Sci. 2013, 13, 1–13. [Google Scholar] [CrossRef]
- Ram, M.; Kumar, S. Yield and Resource Use Optimization in Late Transplanted Mint (Mentha arvensis) under Subtropical Conditions. J. Agron. Crop. Sci. 1998, 180, 109–112. [Google Scholar] [CrossRef]
- Akram, M.; Uzair, M.; Malik, N.S.; Mahmood, A.; Sarwer, N.; Madni, A.; Asif, H.M. Mentha arvensis Linn: A review article. J. Medic. Plants Res. 2011, 5, 4499–4503. [Google Scholar]
- Singh, M.; Singh, A.; Singh, S.; Tripathi, R.S.; Singh, A.K.; Patra, D.D. Cowpea (Vigna unguiculata L. Walp.) as a green manure to improve the productivity of a menthol mint (Mentha arvensis L.) intercropping system. Ind. Crops Prod. 2010, 31, 289–293. [Google Scholar] [CrossRef]
- Chand, S.; Patra, N.K.; Anwar, M.; Patra, D.D. Agronomy and uses of menthol mint (Mentha arvensis)—Indian perspective. Proc. Indian. Natl. Sci. Acad. 2004, 70, 269–297. [Google Scholar]
- Costa, A.; Chagas, J.; Bertolucci, S.; Pinto, J. Growth and production of volatiles in fertilized mint. Acta Hortic. 2018, 63–66. [Google Scholar] [CrossRef]
- Adamović, D.S.; Đalović, I.G.; Mrkovački, N.M.; Pandurevic, T.; Bjelic, D.; Tyr, S. Effect of growing season upon microbial status of peppermint (Mentha x piperita L.) rhizosphere. Acta Fytotech. Zootech. 2015, 18, 99–102. [Google Scholar] [CrossRef]
- Mousavinik, S.M.; Asgharipour, M.R.; Sardashti, S. Manure and Light Intensity Affect Growth Characteristics and Essential Oil of Peppermint (Mentha piperita L.). J. Essent. Oil Bear. Plants 2016, 19, 2029–2036. [Google Scholar] [CrossRef]
- Bajeli, J.; Tripathi, S.; Kumar, A.; Tripathi, A.; Upadhyay, R. Organic manures a convincing source for quality production of Japanese mint (Mentha arvensis L.). Ind. Crop. Prod. 2016, 83, 603–606. [Google Scholar] [CrossRef]
- Verma, R.K.; Verma, R.S.; Rahman, L.-U.; Kalra, A.; Patra, D.D. Integrated Nutrient Management on Biomass, Oil Yields and Essential Oil Composition of Peppermint (Mentha piperita L.) and Residual Fertility in a Hilly Soil. J. Essent. Oil Bear. Plants 2016, 19, 582–591. [Google Scholar] [CrossRef]
- USEPA (U.S. Environmental Protection Agency). Exemption of certain pesticide substances from federal insecticide, fungicide, and rodenticide act requirements, Washington DC, USA. 1996. Available online: http://www.nj.gov/dep/enforcement/pcp/bpo/min_risk.pdf (accessed on 23 December 2019).
- Isman, M.B. Botanical Insecticides in the Twenty-First Century-Fulfilling Their Promise? Annu. Rev. Èntomol. 2019, 65, 65. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Maggi, F.; Petrelli, R.; Nicoletti, M. Commentary: making green pesticides greener? The potential of plant products for nanosynthesis and pest control. J. Cluster Sci. 2017, 28, 3–10. [Google Scholar] [CrossRef]
- Isenring, R. Pesticides and the loss of biodiversity; Pesticide Action Network Europe: London, UK, 2010. [Google Scholar]
- Maino, J.L.; Binns, M.; Umina, P. No longer a west-side story – pesticide resistance discovered in the eastern range of a major Australian crop pest, Halotydeus destructor (Acari: Penthaleidae). Crop. Pasture Sci. 2018, 69, 216. [Google Scholar] [CrossRef]
- Balakrishnan, A. Therapeutic uses of peppermint—a review. J. Pharm. Sci. Res. 2015, 7, 474–476. [Google Scholar]
- Varshney, S.C. Indian mint oils. Perf. Flav. 2005, 30, 36–44. [Google Scholar]
- European Pharmacopoeia 5.0, 5.1–5.8. Strasbourg Council of Europe. 2005.
- Lawrence, B.M. Progress in essential oils. Perf. Flav. 1993, 18, 59–71. [Google Scholar]
- Telci, I.; Kacar, O.; Bayram, E.; Arabacı, O.; Demirtaş, I.; Yılmaz, G.; Özcan, I.; Sönmez, Ç.; Göksu, E. The effect of ecological conditions on yield and quality traits of selected peppermint (Mentha piperita L.) clones. Ind. Crop. Prod. 2011, 34, 1193–1197. [Google Scholar] [CrossRef]
- Oroian, C.; Covrig, I.; Odagiu, A.; Mălinaș, C.; Moldovan, C.; Fleșeriu, A. Effects of Cultivation Systems and Environmental Conditions on Peppermint (Mentha × piperita L.) Biomass Yield and Oil Content. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 576–581. [Google Scholar] [CrossRef]
- Santoro, M.V.; Bogino, P.C.; Nocelli, N.; Cappellari, L.D.R.; Giordano, W.F.; Banchio, E. Analysis of Plant Growth-Promoting Effects of Fluorescent Pseudomonas Strains Isolated from Mentha piperita Rhizosphere and Effects of Their Volatile Organic Compounds on Essential Oil Composition. Front. Microbiol. 2016, 7, 11. [Google Scholar] [CrossRef]
- Shahabivand, S.; Padash, A.; Aghaee, A.; Nasiri, Y.; Rezaei, P.F. Plant biostimulants (Funneliformis mosseae and humic substances) rather than chemical fertilizer improved biochemical responses in peppermint. Iran. J. Plant Physiol. 2018, 8, 2333–2344. [Google Scholar]
- Shaikh, M.N.; Kasabe, U.I.; Mokat, D.N. Influence of Rhizosphere Fungi on Essential Oil Production and Menthol Content in Mentha arvensis L. J. Essent. Oil Bear. Plants 2018, 21, 1076–1081. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential Oils: Sources of Antimicrobials and Food Preservatives. Front. Microbiol. 2017, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Sarkic, A.; Stappen, I. Essential Oils and Their Single Compounds in Cosmetics—A Critical Review. Cosmet. 2018, 5, 11. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Van de Vel, E.; Sampers, I.; Raes, K. A review on influencing factors on the minimum inhibitory concentration of essential oils. Crit. Rev. Food Sci. Nutrit. 2019, 59, 357–378. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Kumar, R.; Dubey, N.K.; Tiwari, O.P.; Tripathi, Y.B.; Sinha, K.K. Evaluation of some essential oils as botanical fungitoxicants for the protection of stored food commodities from fungal infestation. J. Sci. Food Agric. 2007, 87, 1737–1742. [Google Scholar] [CrossRef]
- Reddy, D.N.; Al-Rajab, A.J.; Sharma, M.; Moses, M.M.; Reddy, G.R.; Albratty, M. Chemical constituents, in vitro antibacterial and antifungal activity of Mentha piperita L. (peppermint) essential oils. J. King Saud Univ. Sci. 2017. [Google Scholar]
- Choi, O.; Cho, S.K.; Kim, J.; Park, C.G.; Kim, J. Antibacterial properties and major bioactive components of Mentha piperita essential oils against bacterial fruit blotch of watermelon. Arch. Phytopathol. Plant Prot. 2016, 49, 325–334. [Google Scholar] [CrossRef]
- Kumar, A.; Shukla, R.; Singh, P.; Singh, A.K.; Dubey, N.K. Use of essential oil from Mentha arvensis L. to control storage moulds and insects in stored chickpea. J. Sci. Food Agric. 2009, 89, 2643–2649. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Deans, S.G.; Eaglesham, E. Relationship between bioactivity and chemical composition of commercial essential oils. Flavour Fragr. J. 1998, 13, 98–104. [Google Scholar] [CrossRef]
- Giamperi, L.; Fraternale, D.; Ricci, D. The In Vitro Action of Essential Oils on Different Organisms. J. Essent. Oil Res. 2002, 14, 312–318. [Google Scholar] [CrossRef]
- Sokovic, M.; Van Griensven, L.J. Antimicrobial activity of essential oils and their components against the three major pathogens of the cultivated button mushroom, Agaricus bisporus. Eur. J. Plant Pathol. 2006, 116, 211–224. [Google Scholar] [CrossRef]
- Combrinck, S.; Regnier, T.; Kamatou, G. In vitro activity of eighteen essential oils and some major components against common postharvest fungal pathogens of fruit. Ind. Crop. Prod. 2011, 33, 344–349. [Google Scholar] [CrossRef]
- Chao, S.C.; Young, D.G.; Oberg, C.J. Screening for Inhibitory Activity of Essential Oils on Selected Bacteria, Fungi and Viruses. J. Essent. Oil Res. 2000, 12, 639–649. [Google Scholar] [CrossRef]
- Baruah, P.; Sharma, R.K.; Singh, R.S.; Ghosh, A.C. Fungicidal Activity of Some Naturally Occurring Essential Oils Against Fusarium moniliforme. J. Essent. Oil Res. 1996, 8, 411–412. [Google Scholar] [CrossRef]
- El-Zemity, S.R.; Radwan, M.A.; El-Monam Mohamed, S.A.; Sherby, S.M. Antibacterial screening of some essential oils, monoterpenoids and novel N-methyl carbamates based on monoterpenoids against Agrobacterium tumefaciens and Erwinia carotovora. Arch. Phytopathol. Plant Prot. 2008, 41, 451–461. [Google Scholar] [CrossRef]
- Mehrparvar, M.; Goltapeh, E.M.; Safaie, N.; Ashkani, S.; Hedesh, R.M. Antifungal activity of essential oils against mycelial growth of Lecanicillium fungicola var. fungicola and Agaricus bisporus. Ind. Crop. Prod. 2016, 84, 391–398. [Google Scholar] [CrossRef]
- Basílico, M.Z. Inhibitory effects of some spice essential oils on Aspergillus ochraceus NRRL 3174 growth and ochratoxin A production. Lett. Appl. Microbiol. 1999, 29, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Xing, F.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Inhibitory Effect of Essential Oils on Aspergillus ochraceus Growth and Ochratoxin A Production. PLOS ONE 2014, 9, e108285. [Google Scholar] [CrossRef]
- Işcan, G.; Kïrïmer, N.; Kürkcüoǧlu, M.; Başer, H.C.; Demïrcï, F. Antimicrobial Screening of Mentha piperita Essential Oils. J. Agric. Food Chem. 2002, 50, 3943–3946. [Google Scholar]
- Ahmad, A.; Darokar, M.P.; Tandon, S. Antimicrobial activity of isolates and derivatives of Indian Mentha arvensis essential oil. J. Essent. Oil Bear. Plants 2011, 14, 796–804. [Google Scholar] [CrossRef]
- Montes-Belmont, R.; Carvajal, M. Control of Aspergillus flavus in maize with plant essential oils and their components. J. Food Prot. 1998, 61, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R.; Zhou, T. Antifungal activity of monoterpenoids against postharvesr pathogens Botrytis cinerea and Monilinia fructicola. J. Essent. Oil Res. 2000, 12, 113–121. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Sharifzadeh, A.; Shokri, H.; Khosravi, A.R.; Abbaszadeh, A. Antifungal efficacy of thymol, carvacrol, eugenol and menthol as alternative agents to control the growth of food-relevant fungi. Journal de Mycologie Médicale 2014, 24, e51–e56. [Google Scholar] [CrossRef]
- Kotan, R.; Kordali, S.; Cakir, A. Screening of antibacterial activities of twenty-one oxygenated monoterpenes. Zeitschrift für Naturforschung C 2007, 62, 507–513. [Google Scholar] [CrossRef]
- Diánez, F.; Santos, M.; Parra, C.; Navarro, M.; Blanco, R.; Gea, F. Screening of antifungal activity of 12 essential oils against eight pathogenic fungi of vegetables and mushroom. Lett. Appl. Microbiol. 2018, 67, 400–410. [Google Scholar] [CrossRef]
- Pattnaik, S.; Subramanyam, V.R.; Kole, C. Antibacterial and antifungal activity of ten essential oils in vitro. Microbios 1996, 86, 237–246. [Google Scholar]
- Hossain, F.; Follett, P.; Vu, K.D.; Harich, M.; Salmieri, S.; Lacroix, M. Evidence for synergistic activity of plant-derived essential oils against fungal pathogens of food. Food Microbiol. 2016, 53, 24–30. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. Antimicrobial potential and chemical composition of Mentha piperita oil in liquid and vapour phase against food spoiling microorganisms. Food Control. 2011, 22, 1707–1714. [Google Scholar] [CrossRef]
- Bouchra, C.; Mohamed, A.; Hassani Mina, I.; Hmamouchi, M. Antifungal activity of essential oils from several medicinal plants against four postharvest citrus pathogens. Phytopathol. Mediterr. 2003, 42, 251–256. [Google Scholar]
- Moghaddam, M.; Pourbaige, M.; Tabar, H.K.; Farhadi, N.; Hosseini, S.M.A. Composition and Antifungal Activity of Peppermint (Mentha piperita) Essential Oil from Iran. J. Essent. Oil Bear. Plants 2013, 16, 506–512. [Google Scholar] [CrossRef]
- Anaruma, N.D.; Schmidt, F.L.; Duarte, M.C.T.; Figueira, G.M.; Delarmelina, C.; Benato, E.A.; Sartoratto, A. Control of Colletotrichum gloeosporioides (Penz.) Sacc. In yellow passion fruit using Cymbopogon citratus essential oil. Braz. J. Microbiol. 2010, 41, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Christian, E.J.; Goggi, A.S. Aromatic Plant Oils as Fungicide for Organic Corn Production. Crop. Sci. 2008, 48, 1941. [Google Scholar] [CrossRef]
- Edris, A.E.; Farrag, E.S. Antifungal activity of peppermint and sweet basil essential oils and their major aroma constituents on some plant pathogenic fungi from the vapor phase. Food/Nahrung 2003, 47, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.K.; Raina, A.P.; Dureja, P. Evaluation of the antifungal and phytotoxic effects of various essential oils against Sclerotium rolfsii (Sacc) and Rhizoctonia bataticola (Taub). Arch. Phytopathol. Plant Prot. 2009, 42, 65–72. [Google Scholar] [CrossRef]
- El Asbahani, A.; Jilale, A.; Voisin, S.N.; Aït Addi, E.H.; Casabianca, H.; El Mousadik, A.; Hartmann, D.J.; Renaud, F.N. Chemical composition and antimicrobial activity of nine essential oils obtained by steam distillation of plants from the Souss-Massa Re Region (Morocco). J. Essent. Oil Res. 2015, 27, 34–44. [Google Scholar] [CrossRef]
- Soković, M.D.; Vukojevic, J.; Marin, P.D.; Brkić, D.D.; Vajs, V.; Van Griensven, L.J.L.D. Chemical Composition of Essential Oils of Thymus and Mentha Species and Their Antifungal Activities. Molecules 2009, 14, 238–249. [Google Scholar] [CrossRef]
- Vieira-Brock, P.L.; Vaughan, B.M.; Vollmer, D.L. Comparison of antimicrobial activities of natural essential oils and synthetic fragrances against selected environmental pathogens. Biochim. Open 2017, 5, 8–13. [Google Scholar] [CrossRef]
- Antunes, M.D.C.; Cavaco, A.M. The use of essential oils for postharvest decay control. A review. Flavour Fragr. J. 2010, 25, 351–366. [Google Scholar] [CrossRef]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of plant essential oils on postharvest control of rot caused by fungi on four cultivars of apples in vivo. Flavour Fragr J. 2010, 25, 171–177. [Google Scholar] [CrossRef]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of Plant Essential Oils on Postharvest Control of Rots Caused by Fungi on Different Stone Fruits In Vivo. J. Food Prot. 2013, 76, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Ziedan, E.H.E.; Farrag, E.S. Fumigation of peach fruits with essential oils to control postharvest decay. Res. J. Agric. Biol. Sci. 2008, 4, 512–519. [Google Scholar]
- Varma, J. Efficacy of essential oils of Caesulia axillaris and Mentha arvensis against some storage pests causing biodeterioration of food commodities. Int. J. Food Microbiol. 2001, 68, 207–210. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N.; Banerji, R.; Chansouria, J. Evaluation of some essential oils as botanical fungitoxicants in management of post-harvest rotting of citrus fruits. World J. Microbiol. Biotechnol. 2004, 20, 317–321. [Google Scholar] [CrossRef]
- Servili, A.; Feliziani, E.; Romanazzi, G. Exposure to volatiles of essential oils alone or under hypobaric treatment to control postharvest gray mold of table grapes. Postharvest Boil. Technol. 2017, 133, 36–40. [Google Scholar] [CrossRef]
- Hadian, J.; Ghasemnezhad, M.; Ranjbar, H.; Frazane, M.; Ghorbanpour, M. Antifungal potency of some essential oils in control of postharvest decay of strawberry caused by Botrytis cinerea, Rhizopus stolonifer and Aspergillus niger. J. Essent. Oil Bear. Plants 2008, 11, 553–562. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, S.Y.; Yin, J.-J.; Parry, J.; Yu, L.L. Enhancing Antioxidant, Antiproliferation, and Free Radical Scavenging Activities in Strawberries with Essential Oils. J. Agric. Food Chem. 2007, 55, 6527–6532. [Google Scholar] [CrossRef]
- Chaemsanit, S.; Matan, N.; Matan, N. Effect of peppermint oil on the shelf-life of dragon fruit during storage. Food Control. 2018, 90, 172–179. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Serrano, M.; Bailén, G.; Guillen, F.; Zapata, P.J.; Valverde, J.M.; Castillo, S.; Fuentes, M.; Valero, D. The use of a natural fungicide as an alternative to preharvest synthetic fungicide treatments to control lettuce deterioration during postharvest storage. Postharvest Boil. Technol. 2008, 47, 54–60. [Google Scholar] [CrossRef]
- Ben Hsouna, A.; Touj, N.; Hammami, I.; Dridi, K.; Al-Ayed, A.S.; Hamdi, N. Chemical Composition and in vivo Efficacy of the Essential Oil of Mentha piperita L. in the Suppression of Crown Gall Disease on Tomato Plants. J. Oleo Sci. 2019, 68, 419–426. [Google Scholar] [CrossRef]
- Wonni, I.; Ouedraogo, S.L.; Ouedraogo, I.; Sanogo, L. Antibacterial activity of extracts of three aromatic plants from Burkina Faso againstrice pathogen, Xanthomanas oryzae. Afr. J. Microbiol. Res. 2016, 10, 681–686. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.; Seleim, M.A.; Abd-El-Moneem, K.M.; Saead, F.A. Integrated effect of Glomus mosseae and selected plant oils on the control of bacterial wilt disease of tomato. Crop. Prot. 2014, 66, 67–71. [Google Scholar] [CrossRef]
- Al-Mughrabi, K.I.; Coleman, W.K.; Vikram, A.; Poirier, R.; Jayasurija, K.E. Effectiveness of essential oils and their combinations with aluminium starch octenylsuccinate on potato storage pathogens. J. Essent. Oil Bear. Plants 2013, 16, 23–31. [Google Scholar] [CrossRef]
- Devlieghere, F.; Vermeulen, A.; Debevere, J. Chitosan: antimicrobial activity, interactions with food components and applicability as a coating on fruit and vegetables. Food Microbiol. 2004, 21, 703–714. [Google Scholar] [CrossRef]
- De Oliveira, K.; Árabe, R.; Berger, L.R.R.; De Araújo, S.A.; Câmara, M.P.S.; De Souza, E.L. Synergistic mixtures of chitosan and Mentha piperita L. essential oil to inhibit Colletotrichum species and anthracnose development in mango cultivar Tommy Atkins. Food Microbiol. 2017, 66, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Guerra, I.C.D.; De Oliveira, P.D.L.; Pontes, A.L.D.S.; Lúcio, A.S.S.C.; Tavares, J.F.; Barbosa-Filho, J.M.; Madruga, M.S.; De Souza, E.L. Coatings comprising chitosan and Mentha piperita L. or Mentha×villosa Huds essential oils to prevent common postharvest mold infections and maintain the quality of cherry tomato fruit. Int. J. Food Microbiol. 2015, 214, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Guerra, I.C.D.; De Oliveira, P.D.L.; Santos, M.M.F.; Lúcio, A.S.S.C.; Tavares, J.F.; Barbosa-Filho, J.M.; Madruga, M.S.; De Souza, E.L. The effects of composite coatings containing chitosan and Mentha piperita L. or x villosa (Huds) essential oil on postharvest mold occurrence and quality of table grape cv. Isabella. Innov. Food Sci. Emerg. Technol. 2016, 34, 112–121. [Google Scholar] [CrossRef]
- Kuorwel, K.K.; Cran, M.J.; Sonneveld, K.; Miltz, J.; Bigger, S.W. Essential Oils and Their Principal Constituents as Antimicrobial Agents for Synthetic Packaging Films. J. Food Sci. 2011, 76, R164–R177. [Google Scholar] [CrossRef]
- Serrano, M.; Martinez-Romero, D.; Guillen, F.; Valverde, J.M.; Zapata, P.J.; Salvador Castillo, S.; Valero, D. The addition of essential oils to MAP as a tool to maintain the overall quality of fruits. Trends Food Sci. Technol. 2008, 19, 464–471. [Google Scholar] [CrossRef]
- Valverde, J.M.; Guillen, F.; Martínez-Romero, D.; Castillo, S.; Serrano, M.; Valero, D. Improvement of Table Grapes Quality and Safety by the Combination of Modified Atmosphere Packaging (MAP) and Eugenol, Menthol, or Thymol. J. Agric. Food Chem. 2005, 53, 7458–7464. [Google Scholar] [CrossRef]
- Martinez-Romero, D.; Castillo, S.; Valverde, J.M.; Guillen, F.; Valero, D.; Serrano, M. The use of a natural aromatic essential oils helps to maintain post-harvest quality of “Crimson” table grapes. Acta Horticulturae 2005, 682, 1723–1730. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Insecticidal properties of Mentha species: A review. Ind. Crop. Prod. 2011, 34, 802–817. [Google Scholar] [CrossRef]
- Saeidi, K.; Mirfakhraie, S. Chemical composition and insecticidal activity Mentha piperita L. essential oil against the cowpea seed beetle Callosobruchus maculatus F. (Coleoptera: Bruchidae). J. Èntomol. Acarol. Res. 2017, 49, 127–134. [Google Scholar] [CrossRef]
- Jesser, E.N.; Werdin-González, J.O.; Murray, A.P.; Ferrero, A.A. Efficacy of essential oils to control the Indian meal moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae). J. Asia-Pacific Èntomol. 2017, 20, 1122–1129. [Google Scholar] [CrossRef]
- Somaye, A.; Khalil, T.J.; Sohrab, I.; Mohammad, K. Contact toxicity of ploy lactic acid nanofibers loaded with two essential oils against Plodia interpunctella Hub. (Lepidoptera: Pyralidae). J. Biopest. 2017, 10, 50–59. [Google Scholar]
- Vendan, S.E.; Manivannan, S.; Sunny, A.M.; Murugesan, R. Phytochemical residue profiles in rice grains fumigated with essential oils for the control of rice weevil. PLOS ONE 2017, 12, e0186020. [Google Scholar] [CrossRef]
- Rajkumar, V.; Gunasekaran, C.; Christy, I.K.; Dharmaraj, J.; Chinnaraj, P.; Paul, C.A. Toxicity, antifeedant and biochemical efficacy of Mentha piperita L. essential oil and their major constituents against stored grain pest. Pestic. Biochem. Physiol. 2019, 156, 138–144. [Google Scholar] [CrossRef]
- Khani, M.; Marouf, A.; Amini, S.; Yazdani, D.; Farashiani, M.E.; Ahvazi, M.; Khalighi-Sigaroodi, F.; Hosseini-Gharalari, A. Efficacy of Three Herbal Essential Oils Against Rice Weevil, Sitophilus oryzae (Coleoptera: Curculionidae). J. Essent. Oil Bear. Plants 2017, 20, 937–950. [Google Scholar] [CrossRef]
- Mishra, B.B.; Tripathi, S.; Tripathi, C. Sub-lethal Activity of Plant Volatile Essential Oils in Management of Red Flour Beetle Tribolium castaneum (Coleoptera: Tenebrionidae). J. Essent. Oil Bear. Plants 2014, 17, 1211–1218. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Davari, M.; Razmjou, J.; Naseri, B. Separate and Combined Effects of Mentha piperata and Mentha pulegium Essential Oils and a Pathogenic Fungus Lecanicillium muscarium Against Aphis gossypii (Hemiptera: Aphididae). J. Econ. Èntomol. 2017, 110, 1025–1030. [Google Scholar] [CrossRef]
- Pascual-Villalobos, M.; Cantó-Tejero, M.; Vallejo, R.; Guirao, P.; Rodríguez-Rojo, S.; Cocero, M. Use of nanoemulsions of plant essential oils as aphid repellents. Ind. Crop. Prod. 2017, 110, 45–57. [Google Scholar] [CrossRef]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholinesterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crop. Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- Koundal, R.; Dolma, S.K.; Chand, G.; Agnihotri, V.K.; Reddy, S.G.E. Chemical composition and insecticidal properties of essential oils against diamondback moth (Plutella xylostella L.). Toxin Rev. 2018, 1–11. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal and repellent activity of selected essential oils against of the pollen beetle, Meligethes aeneus (Fabricius) adults. Ind. Crop. Prod. 2011, 34, 888–892. [Google Scholar] [CrossRef]
- Khater, H.F.; Ramadan, M.Y.; Mageid, A.D.A. In vitro control of the camel nasal botfly, Cephalopina titillator, with doramectin, lavender, camphor, and onion oils. Parasitol. Res. 2013, 112, 2503–2510. [Google Scholar] [CrossRef]
- Lachance, S.; Grange, G. Repellent effectiveness of seven plant essential oils, sunflower oil and natural insecticides against horn flies on pastured dairy cows and heifers. Med. Veter. Entomol. 2014, 28, 193–200. [Google Scholar] [CrossRef]
- Park, J.-H.; Yang, J.-Y.; Lee, H.-S. Acaricidal Activity of Constituents Derived from Peppermint Oil against Tyrophagus putrescentiae. J. Food Prot. 2014, 77, 1819–1823. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Lee, H.-S. Chemical Composition and Acaricidal Activities of Essential Oils of Litsea cubeba Fruits and Mentha arvensis Leaves Against House Dust and Stored Food Mites. J. Essent. Oil Bear. Plants 2016, 19, 1721–1728. [Google Scholar] [CrossRef]
- Mahdavikia, F.; Saharkhiz, M.J. Secondary metabolites of peppermint change the morphophysiological and biochemical characteristics of tomato. Biocatal. Agric. Biotechnol. 2016, 7, 127–133. [Google Scholar] [CrossRef]
- Mahdavikia, F.; Saharkhiz, M.J.; Karami, A. Defensive response of radish seedlings to the oxidative stress arising from phenolic compounds in the extract of peppermint (Mentha × piperita L.). Sci. Hortic. 2017, 214, 133–140. [Google Scholar] [CrossRef]
- Możdżeń, K.; Barabasz-Krasny, B.; Stachurska-Swakoń, A.; Zandi, P.; Puła, J. Effect of Aqueous Extracts of Peppermint (Mentha × piperita L.) on the Germination and the Growth of Selected Vegetable and Cereal Seeds. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2019, 47, 412–417. [Google Scholar]
- Synowiec, A.; Kalemba, D.; Drozdek, E.; Bocianowski, J. Phytotoxic potential of essential oils from temperate climate plants against the germination of selected weeds and crops. J. Pest Sci. 2017, 90, 407–419. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxicity of Essential Oils on Selected Weeds: Potential Hazard on Food Crops. Plants 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Mahdavikia, F.; Saharkhiz, M.J. Phytotoxic activity of essential oil and water extract of peppermint (Mentha×piperita L. CV. Mitcham). J. Appl. Res. Med. Aromat. Plants 2015, 2, 146–153. [Google Scholar] [CrossRef]
- Cavalieri, A.; Caporali, F. Effects of essential oils of cinnamon, lavender and peppermint on germination of Mediterranean weeds. Allelop. J 2010, 25, 441–452. [Google Scholar]
- Jurová, J.; Matoušková, M.; Wajs-Bonikowska, A.; Kalemba, D.; Renčo, M.; Sedlák, V.; Gogaľová, Z.; Poráčová, J.; Šalamún, P.; Gruľová, D. Potential Phytotoxic Effect of Essential Oil of Non-Native Species Impatiens parviflora DC. Plants 2019, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- Rolli, E.; Marieschi, M.; Maietti, S.; Sacchetti, G.; Bruni, R. Comparative phytotoxicity of 25 essential oils on pre- and post-emergence development of Solanum lycopersicum L.: A multivariate approach. Ind. Crop. Prod. 2014, 60, 280–290. [Google Scholar] [CrossRef]
- Campiglia, E.; Mancinelli, R.; Cavalieri, A.; Caporali, F. Use of essential oils of cinnamon (Cinnamomum zeylanicum L.), lavender (Lavandula spp.) and peppermint (Mentha x piperita L.) for weed control. Ital. J. Agron. 2007, 2, 171–175. [Google Scholar] [CrossRef]
- Synowiec, A.; Smęda, A.; Adamiec, J.; Kalemba, D.; Agnieszka, S.; Aleksandra, S.; Janusz, A.; Danuta, K. The effect of microencapsulated essential oils on the initial growth of maize (Zea mays) and common weeds (Echinochloa crus-galli and Chenopodium album). Prog. Plant Prot. 2016, 56, 372–378. [Google Scholar]
- Khare, P.; Srivastava, S.; Nigam, N. Impact of essential oils of E. citriodora, O. basilicum and M. arvensis on three different weeds and soil microbial activities. Environ. Technol. Innovat. 2019, 14, 100343. [Google Scholar] [CrossRef]
- Ayala-Zavala, J.F.; González-Aguilar, G.A.; Del-Toro-Sánchez, L. Enhancing Safety and Aroma Appealing of Fresh-Cut Fruits and Vegetables Using the Antimicrobial and Aromatic Power of Essential Oils. J. Food Sci. 2009, 74, R84–R91. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Bouquillon, S.; Fauconnier, M.-L. Maes Encapsulation of Essential Oils for the Development of Biosourced Pesticides with Controlled Release: A Review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef] [PubMed]
- Adamiec, J.; Kalemba, D. Analysis of microencapsulation ability of essential oils during spray drying. Drying Technol. 2006, 24, 1127–1132. [Google Scholar] [CrossRef]
- Baranauskienė, R.; Bylaitė, E.; Žukauskaitė, J.; Venskutonis, R.P. Flavor Retention of Peppermint (Mentha piperita L.) Essential Oil Spray-Dried in Modified Starches during Encapsulation and Storage. J. Agric. Food Chem. 2007, 55, 3027–3036. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, M.; Ji, N.; Liu, J.; Xiong, L.; Sun, Q. Morphology and Characteristics of Starch Nanoparticles Self-Assembled via a Rapid Ultrasonication Method for Peppermint Oil Encapsulation. J. Agric. Food Chem. 2017, 65, 8363–8373. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Ma, Y.; Hayat, K.; Jia, C.; Xia, S.; Zhang, X. Morphology and release profile of microcapsules encapsulating peppermint oil by complex coacervation. J. Food Eng. 2011, 104, 455–460. [Google Scholar] [CrossRef]
- Yildiz, Z.I.; Celebioglu, A.; Kilic, M.E.; Durgun, T.U. Menthol/cyclodextrin inclusion complex nanofibers: enhanced water-solubility and high-temperature stability of menthol. J. Food Eng. 2018, 224, 27–36. [Google Scholar] [CrossRef]
- Donsi’, F.; Annunziata, M.; Sessa, M.; Ferrari, G. Nanoencapsulation of essential oils to enhance their antimicrobial activity in foods. LWT 2011, 44, 1908–1914. [Google Scholar] [CrossRef]
- Beyki, M.; Zhaveh, S.; Khalili, S.T.; Rahmani-Cherati, T.; Abollahi, A.; Bayat, M.; Tabatabaei, M.; Mohsenifar, A. Encapsulation of Mentha piperita essential oils in chitosan–cinnamic acid nanogel with enhanced antimicrobial activity against Aspergillus flavus. Ind. Crop. Prod. 2014, 54, 310–319. [Google Scholar] [CrossRef]


{kind=link}
| Fungi/Bacteria (B) | MIC or Total Inhibition Concentration | No. of Essential Oils Mint Oil Composition [%] | Methods Results for the Most Active Essential Oil | Ref. |
|---|---|---|---|---|
| Alternaria alternata Alternaria solani Aspergillus flavus Aspergillus niger Fusarium solani Rhizopus solani Rhizopus spp. | 117.0/57.9μg/mL1 127.1/129.0 122.0/110.7 49.5/63.5 130.7/89.8 44.11/63.9 149.7/137.1 | 4 EOs, 4 compounds MPEO menthone 28.1, menthol 4.8, menthyl acetate 9.5, limonene 7.1 MAEO, menthol 78.9, menthone 6.4 | broth microdilution, agar disc diffusion (15 μL), positive control: fluconazole 30 μg M. spicata and M. longifolia similar results as MPEO | [39] |
| Alternaria brassicae Botrytis cinerea Cladobotryum mycophilum Fusarium oxysporum Phytophthora parasitica Pythium aphanidermatum Sclerotinia sclerotiorumisolated from vegetables and mushrooms | 16.2%2 - 7.4% 15.8% 5.7% - 6% | 12 EOs MPEO menthol 42.0, menthone 28.8, 1,8-cineole 7.1 | disc diffusion 8 μL of 5–30% EO solution MPEO oil belonged to four most effective | [60] |
| Alternaria citrii Aspergillus fumigatus Aspergillus oryzae Fusarium oxysporum Fusarium solani Helminthosporium compactum Macrophomina phaseolina Sclerotium rolfsii | 0.25 μL/mL 1.0 1.0 3.0 2.0 0.5 2.0 0.25 | 4 EOs MPEO no data | disc diffusion, 5 μL, agar dilution 0.16–20 μg/mL MPEO less active than other three | [61] |
| Alternaria citrii Botrytis cinerea Colletotrichum gloeosporioides Lasiodiplodia theobromae Penicillium digitatum isolated from fruits | 3000 μL/L 3000 3000 >3000 2000 | 18 EOs MPEO menthol 40.7, menthone 21.7 | agar dilution thyme 500–1000 μL/L (3000 P. digitatum) | [47] |
| Aspergillus ochraceus | 2000 μg/L (broth) 1500 μg/L (vapor) | 5 EOs, 5 compounds MPEO menthol 50 | broth dilution/vapor phase cinnamon oil and cinnamaldehyde: 250–500 μg/L (broth), 150–250 μg/L (vapor), ochratoxin A production inhibited at 200 μg/L | [53] |
| Aspergillus ochraceus | 1000 ppm | 4 EOs MAEO, no data | broth dilution MAEO and oregano oil were the most effective in inhibition of fungal growth and ochratoxin A production | [52] |
| Aspergillus flavus Aspergillus niger Aspergillus parasiticus Penicillium chrysogenum | 10000 ppm 5000 2500 1250 | 8 EOs and EOs combinations MPEO menthol, menthone | broth dilution, vapor phase MPEO less active than thyme (312.5–1250 ppm) and oregano oils, similar activity to cinnamon oil, more active than other four oils | [62] |
| Aspergillus flavus Aspergillus niger Fusarium oxysporum Mucor spp. Penicillium digitatum | 1.13/2.25 mg/mL3 1.13/2.25 1.13/2.25 1.13/2.25 2.25/4.5 | MPEO | agar dilution (MIC), broth dilution (MFC), well diffusion, vapor phase | [63] |
| Aspergillus flavus Aspergillus fumigatus Aspergillus niger Botryodiplodia theobromae Cladosporium cladosporioides Fusarium oxysporum Helminthosporium oryzae Macrophomina sp. Sclerotium rolfsii | 0.1 mg/mL 0.1 <0.5 0.1 0.1 | 18 EOs MAEO menthol 73, menthone 6.1 | agar dilution, positive control: four synthetic fungicides MAEO was the most efficient of EOs and more efficient than synthetic fungicides at 0.1 mg/mL four fungi were inhibited totally, other 72–100% inhibition aflatoxin B1 production by A. flavus inhibited at 0.05 mg/mL | [40] |
| Alternaria alternata Aspergillus fumigatus Aspergillus candidus Aspergillus nidulans Aspergillus versicolor Cladosporium cladosporioides Curvularia lunata Fusarium nivale Fusarium oxysporum Fusarium roseum Penicillium sp. Monilia sp. Trichoderma viride | 400 μg/L | 18 EOs MAEO no data | agar dilution, positive control: nine synthetic fungicides MAEO was the most efficient of EOs and more efficient than all synthetic fungicides at 400 μg/L 11 fungi were inhibited totally, other two >84% | [43] |
| Botrytis cinerea Geotrichum citri-aurantii Phytophthora citrophthora Penicillium digitatum | no inhibition at 250 ppm | 19 EOs MPEO menthol 50, menthone 30, menthyl acetate 10 | radial growth on plate at different concentration, positive control: four synthetic fungicides Chrysanthemum viscidehirtum total inhibition at 150 ppm, synthetic fungicides at 50 ppm | [64] |
| Phytophthora cinnamomi Pyrenochaeta lycoprsici Verticillium dahliae | 800 ppm 400 800 | 8 EOs MPEO menthol 39.0, menthone 21.0, menthofuran 19.5, 1,8-cineole 7.0 | agar dilution oregano 200, 50, 50 ppm, resp. | [45] |
| Dreschlera spicifera Fusarium oxysporum f.sp. ciceris Macrophomina phaseolina | 1600 ppm >1600 800 | MPEO menthol 25.2, menthone 30.6 | agar dilution | [65] |
| Colletotrichum gloeosporioides isolated from fruits | 2.0 mg/mL | 28 EOs MPEO, no data | agar microdilution, positive control: amphotericin B 5–60 μL/mL coriander leaf, two lemongrass sp. 0.25 mg/mL (lemongrass oil evaluated on passion fruit) | [66] |
| Fusarium spp. Penicillium spp. Phythium spp. isolated from corn seeds | 1000 μL/L 1000 >1000 | 18 EOs MPEO, no data | agar dilution oregano MIC 100–200 μL/L | [67] |
| Mucor sp. Rhizopus stolonifer Sclerotinia sclerotiorum | 30 μL/400 mL air | 2 EOs, 4 compounds MPEO menthol 33.3, menthone 29.5, 1,8-cineole 7.0 | vapor phase sweet basil and menthol 30 μL/400 mL air, menthone not active | [68] |
| Rhizoctonia botaticola Sclerotium rolfsii | 1000 μg/mL | 20 EOs MAEO, no data | agar dilution 6 EOs totally inhibited both fungi’s growth at 1000 μg/mL | [69] |
| Lecanicillium fungicola var. fungicola | 750–1000 μL/L | 11 EOs MPEO menthol 39.2, menthyl acetate 20.4, menthone 15.3 | broth dilution mushroom Agaricus bisporus MPEO was similarly active against mushroom and its pest, savory and thyme oils showed the best selectivity index | [51] |
| Aspergillus niger Penicillium funiculosum | 11.4 μg/mL 11.4 | 9 EOs MPEO linalool 41.4 linalyl ac 39.5 | agar dilution Thymus letrobotrys 2.7 and 2.2 μg/mL | [70] |
| Alternaria alternata Aspergillus flavus Aspergillus fumigatus Cladosporium herbarum Fusarium oxysporum Aspergillus veriscolor Fusarium acuminatum Fusarium solani Fusarium tabacinum Monilinia fructicola Penicilliumspp. Rhizoctonia solani Sclerotinia minor Sclerotinia sclerotiorum (B) Pseudomonas syringae (B) Xanthomonas campestris | 1.50 μg/mL 10.0 0.50 1.50 1.50 10.0 2.50 10.0 1.50 5.50 1.50 1.50 10.0 10.0 2.50 80.0 | MPEO menthol 36.0, isomenthone 23.5, menthone 24.6, menthyl acetate 9.0, menthofuran 6.9 | disc diffusion 10 μL, broth microdilution, positive control: amphotericin B MIC 1–5 μg/mL menthol, menthone MIC against P. syringae 2.0, 1.0 μg/mL X. campestris 2.0, 2.0 μg/mL | [41] |
| Alternaria alternata Aspergillus flavus Aspergillus niger Aspergillus ochraceus Aspergillus terreus Aspergillus versicolor Cladosporium cladosporioides Fusarium tricinctum Penicillium funiculosum Penicillium ochrochloron | 1.5–3.0 μL/mL in ethanol 1.0–2.5 μL/mL in Tween | 4 EOs MPEO menthol 37.4, menthone 12.7, limonene 6.9, menthofuran 6.8 | agar macro- (in ethanol) and micro- (in Tween) dilution, positive control: bifonazol MIC 10–15 μL/mL thyme oil 0.125–0.5 μL/mL in ethanol, 0.05–0.25 in Tween menthol 0.25–1.5 μL/mL in ethanol, 0.05–1.0 μL/mL in Tween | [71] |
| Trichoderma harzianum Verticillium fungicola (B) Pseudomonas tolaasii | 3–4 μL/mL | 10 EOs, 10 compounds MPEO menthol 37.4, menthyl acetate 17.4, menthone 12.7 | microdilution, macrodilution, disc diffusion, vapor phase, positive control: bifonazol and prochloraz (fungi), streptomycin + penicillin (bacteria) oregano and thyme 1.5–2.0 μL/mL | [46] |
| (B) Agrobacterium tumefaciens (B) Erwinia carotovora | 13 EOs, 14 compounds MPEO, no data | agar diffusion, 50 μL solution MPEO moderately active at 200 mg/mL 6 EOs were effective, MPEO showed weak activity | [50] | |
| Aspergillus flavus Aspergillus parasiticus Fusarium solani Sclerotium rolfsii (B) Pseudomonas syringae pv. phaseolicola (B) Pseudomonas syringae pv. tomato (B) Pseudomonas syringae pv. syringae (B) Xanthomonas campestris pv. campestris (B) Xanthomonas campestris pv. phaseoli | - - - - 0.07–0.625 mg/mL 0.156–0.312 0.156–0.312 0.312–0.625 0.625–2.5 | four MPEO menthol 27.5–42.3, menthone 18.4–27.9 | fungi: agar diffusion, 50 μL, weak activity bacteria: microdilution menthol 0.07–1.25 mg/mL menthone 1.25–2.5 mg/mL | [54] |
| Fungi/Bacteria (B) | No. of essential oils Mint oil Composition [%] | Host plant Methods Results | Ref. |
|---|---|---|---|
| Botrytis cinerea Penicillium expansum isolated from apples | 11 EOs MPEO menthol 38.3, menthyl acetate 29.4, MAEO menthol 85.9, menthone 3.0 | apples wounded fruits inoculated with spore suspension, treated with 1% and 10% EO emulsion, stored at 4 °C for 30 days, positive control: tebuconazole 125 μg/mL both mint oils at 10% were less effective than thyme and oregano oils and more active than other seven oils | [74] |
| Botrytis cinerea Monilinia laxa isolated from fruits | 11 EOs MPEO menthol 42.1, menthone 20.8 MAEO menthol 33.3, menthone 20.8, isomenthone 10.7 | apricots, nectarines, plums wounded fruits inoculated with spore suspension, treated with 1 and 10% EO emulsion, stored at 1 °C for 28 days, positive control: tebuconazole 125 μg/mL both mint oils at 10% were less effective than thyme and oregano oils and more active than other oils | [75] |
| Colletotrichum asianum Colletotrichum fruticola Colletotrichum tropicale Colletotrichum dianese Colletotrichum karstii | MPEO and chitosan menthol 41.3, isomenthone 23.5, menthone 10.8 | mango in agar dilution assay total inhibition of mycelial growth at 5 mg/mL of MPEO, 10 mg/mL of chitosan, and at a combination of 0.6 μL/mL oil and 5 mg/mL chitosan wounded mango inoculated, immersed in mixtures of MPEO (0.6 or 1.25 μL/mL) and chitosan (5 or 7.5 μL/mL), stored at 4 °C for 30 days, positive control: two synthetic fungicides 10 and 5 μg a.i./mL, respectively anthracnose lesion severity was lower than with synthetic fungicide | [89] |
| Aspergillus niger Monilinia fructicola Rhizopus stolonifer | 2 EOs and combinations, 4 compounds MPEO | peach in in vitro vapor test EOs at 30 μL/400 cm3 inhibited two molds wounded fruits inoculated by fungi, stored at 3 mL/box EO decay reduction and prolonged storage was observed | [76] |
| Penicillium italicum | 20 EOs MAEO, no data | oranges and limes in agar dilution assay MAEO was the most efficient at 100 ppm better than synthetic fungicide wounded fruit inoculated and stored in 100–500 ppm in air of three the most active in vitro EOs (MAEO, Ocimum, and Zingiber) MAEO enhanced storage life from three days in control to nine and 11 days | [78] |
| Botrytis cinerea | 3 EOs and combinations MPEO menthol 34.7, menthone 27.4 | table grapes fruits exposed to EOs vapor for 24 h, under atmospheric and hyperbaric condition, stored in different temperature for nine days all EOs and MPEO/rosemary oil combinations controlled gray mold | [79] |
| Aspergillus niger Botrytis cinerea Penicillium expansum Rhizopus stolonifer | MPEO and chitosan menthol 30.3, isomenthone 26.7, menthyl acetate 8.5 | table grapes fruits immersed in spore inoculum for 1 min., dried, immersed in chitosan/EO solution at different concentrations, stored in room and low temperature chitosan 4 mg/mL/MPEO 1.25 and 2.5 μL/mL delayed the mold growth and reduced infections caused by all fungi | [91] |
| Aspergillus niger Botrytis cinerea Rhizopus stolonifer | 4 EO MPEO menthol 36.2, menthone 32.4 | strawberry fruits dipped in the conidia suspension, then dipped or sprayed in 1000 μL/L EO solution, stored at 7 °C for 10 days, positive control: tiabandazol 1.5:1000 MPEO was the most efficient in inhibiting rot incidence, better than tiabandazol | [80] |
| Aspergillus flavus Aspergillus niger Penicillium spp. Rhizopus spp. | MPEO menthol 41.6, menthone 20.9, isomenthone 9.7 | dragon fruit punched fruit inoculated and stored in oil vapor at 100–1000 μL/L air at 25 °C for 21 days total inhibition of fungal growth at 700, 550, 550, 425 μL/L air, resp. menthol 650–900 μL/L air; menthone 950->1000 μL/L air | [82] |
| Aspergillus niger Botrytis cinerea Penicillium expansum Rhizopus stolonifer | MPEO and chitosan menthol 30.3, isomenthone 26.7, menthyl acetate 8.5 | cherry tomato fruit fruits immersed in spore inoculum for 1 min, dried, immersed in chitosan/EO solution at different concentrations, stored at room and low temperature chitosan 4 mg/mL/MPEO 1.25 and 2.5 μL/mL delayed the mold growth and reduced infections caused by all fungi | [90] |
| total mesophilic bacteria, molds, yeast | MPEO | lettuce in field experiment lettuce sprayed before harvest with FungastopTM that contained 0.2% MPEO and 0.4% citric acid, stored at 2 °C for 28 days treatment lowered microorganisms count at harvest, reduced their increase in storage, and increased shelf life | [83] |
| (B) Agrobacterium tumefaciens, 9 genomovars (B) Agrobacterium vitis (B) Agrobacterium rhizogenes | MPEO menthol 33.6, isomenthone 33.0 | tomato plants MIC in broth microdilution method 0.01–12.5 mg/mL pot experiment: wounded internodes treated with EO solution (100–250 mg/mL), inoculated with A. tumefaciens suspension, stored for one month MPEO at 200 mg/mL completely inhibited the formation of tumors | [84] |
| Ralstonia solanacearum isolated from tomato | 9 EOs MPEO, no data | tomato seedlings growth the most active in in vitro (disc diffusion) MPEO and thyme oils were examined in greenhouse and field growth: seedling roots treated with oil solution (1:100) at transplanting, inoculum added to the soil disease reduction in greenhouse: thyme oil 83%, MPEO 61% | [86] |
| (B) Acidovorax citrulli | 32 EOs, 4 compounds MPEO menthol 40.8, menthone 14.7, isomenthone 8.1 | watermelon seeds in disc diffusion assay only MPEO at 20 μL and basil oil at 50 μL inhibited bacterial growth seeds inoculated, soaked in EO or compound solution (0.01–0.2%) MPEO, menthol, neomenthol, and 1,8-cineole at 0.2% were effective in decontamination of watermelon seeds | [42] |
| Aspergillus flavus | 11 EOs, EOs combinations, 8 compounds MPEO, no data | maize kernels immersed in EO solution (0.1–10%), sprayed with fungal spore suspension MPEO was effective at 8% and other 5 EOs at <7% | [56] |
| Aspergillus flavus Aspergillus fumigatus Aspergillus niger Culvularia spp. Penicillium oxalicum Mucor spp. Rhizopus spp. | 2 EOs MAEO no data | wheat uninoculated and inoculated (A. flavus spore suspension) wheat treated by EO vapors at 1300 and 600 ppm, stored 12 months at room temperature 100% protection of uninoculated and 95% of inoculated wheat orange injured fruits were inoculated (P. italicum spore suspension), treated by EO vapors at 1500 and 1000 ppm, and stored; decay and storage life were enhanced from three to 10 days | [77] |
| Target Organisms | Feeding Damage | Essential oil Composition [%] | Method Formulation | Results1 | Ref. |
|---|---|---|---|---|---|
| Insecticidal | |||||
| Aphis gossypii Glover | polyphagous aphid on watermelon, cotton, and vegetables | MPEO limonene 27.3, menthol 24.7, menthone 14.0 | fumigant toxicity 1.87–50 μL/L air | ED50 15.25 μL/L air | [104] |
| Callosobruchus maculatus F. | store pest of leguminous seeds | MPEO menthone 28.9, menthol 28.5, pulegone 6.9 | fumigant toxicity 20–45 μL/L air | KT50 1.89–3.29 days | [97] |
| repellency test 90–360 μL/L air | R.I.71.6–87.8% | ||||
| Cephalopina titillator Clark | obligate parasite of camels causing Nasopharyngeal myiasis | MPEO no data | larval immersion test 0.05–60% water emulsions | ED50 2.18% after 6 h and 0.47% after 24 h | [109] |
| Drosophila suzukii Matsumura | Insect pest of stone fruits and berry crops | MPEO menthol 46.4, menthone 13.8, menthofuran 7.3 | fumigant toxicity 2.94–11.76 mg/L | LC50 males: 3.87 mg/L LC50 females: 4.10 mg/L | [106] |
| Haematobia irritans L. | livestock pest, bloodsucking fly | MPEO no data | repellent test 5% EO in sunflower oil on the side of pastured cows | 77% less flies on the side of pastured cows after 24 h, as compared with control | [110] |
| Meligethes aeneus (Fabricius) | insect pest of oilseed rape | MAEO menthol 42.5, menthone 21.6, isomenthone 13.5 | acute toxicity 50–5000 μg/cm2 | LD50 964 μg cm−2 after 6 h and 548 μg/cm2 after 24 h | [108] |
| 20 μL EO in 2 mL acetone sprayed on 10 yellow buds of oilseed rape | R.I. 24.6% after 6 h and 45.6% after 24 h | ||||
| Planococcus ficus (Signoret) | insect pest in grape vine | MPEO menthol 34.6, isomenthone 14.6, menthyl acetate 12.4, menthone 9.7, 1,8-cineole 9.3 | contact toxicity 1 mL aqueous solution with 4.5, 9, and 18 mg of EO in 9-cm Petri dish | LC50 against 3rd instar nymphs 5.4 mg/mL LC50 against female adult 8.1 mg/mL | [11] |
| Plodia interpunctella Hüb. | insect pest of diverse stored food products | MPEO isomenthone 48.0, menthone 19.2, 1,8-cineole 9.2 | contact toxicity 5–180 μg/cm2 of filter paper in Petri dish | ED50 53.8 μg/cm2 | [98] |
| fumigant toxicity 40 μL per 8.5-cm Petri dish | KT50 27.1 min. | ||||
| residual toxicity assay 0.05, 0.10 and 0.15% (w/w) to 20 g of whole grain wheat | KT50 3.08–11.3 days | ||||
| Plodia interpunctella Hüb. | -- | MPEO Not assessed | contact toxicity polylactic acid nanofibers loaded with 14% (w/w) EO | ED50 30.3 μL/L of air | [99] |
| Plutella xylostella L. | insect pest of cruciferous crops | MPEO menthol 40.1, menthone 31.1 | residual toxicity water solutions 0.625–5.0 mg/mL | ED50 1.37 mg/mL | [107] |
| repellent activity leaf disks soaked with water solutions (+0.05% tritone) containing three concentrations of EO: 2.5–10.0 mg/mL | ED50 1.33 mg/mL | ||||
| growth inhibition activity 2.5–10 mg/mL after 48 h of feeding on treated leaves | pooled mean inhibition: 37.46% | ||||
| Rhopalosiphum padi L. | aphid pest of temperate cereal crops | MPEO menthol 40.4, menthone 23.5, menthyl acetate 8.3 | repellency EO in acetone at 0.15 μL/cm2 in choice bioassay | ED50 0.13 μL/cm2 | [105] |
| repellency water nanoemulsions, 5% Tween with 0.02, 0.29 and 0.5 μL of the EO per cm2; or 1% Tween with 0.06 or 0.12 μL of the EO per cm2 | R.I. 40 (at a dose 0.02 μL/cm2) to R.I. 90 (at a dose 0.5 μL/cm2) | ||||
| Sitophilus oryzae L. | insect pest of stored cereals | MPEO neoisocarvomenthol 40.6, menthone 27.6, 1,8-cineole 10.6, menthyl acetate 7.5 | fumigant toxicity 20–400 μL/L air, without or with food condition | ED50 47.8 μL/L without food and 45.2 μL/L with food | [100] |
| Sitophilus oryzae L. | MPEO menthol 44.0, menthone 8.3, 1,8-cineole 7.1 | fumigant toxicity 74.4–428.6 μL/L air | ED50 299.5 μL/L of air | [102] | |
| repellency test 2–16 μL/30 cm2 | R.I. 85% | ||||
| Sitophilus oryzae L. | MPEO menthone 31.7, menthol 17.9, piperitone oxide 9.9 | fumigant toxicity 0.05–0.2 μL/mL air | ED50 43.17 μL/L of air | [101] | |
| antifeedant activity ED50 43.17 μL/L of air | FDI 100% | ||||
| Tribolium castaneum Herbst | insect pest of stored grains | MPEO menthone 31.7, menthol 17.9, piperitone oxide 9.9 | fumigant toxicity 0.05–0.2 μL/mL air | ED50 48.68 μL/L of air | |
| antifeedant activity ED50 48.68 μL/L of air | FDI 100% | ||||
| MAEO No data | fumigant toxicity sublethal concentrations 40, 60 or 80% of 24 h LC50 (equal to 18.482 μL of MAEO) | 51% less eggs produced by the insects and 33% lower pupation at an 80% of 24 h LC50 dose | [103] | ||
| contraction method 30% and 60% of 24 h LC50 MAEO in acetone | 61.2% less damaged grain at an 60% of 24 h LC50 dose | ||||
| Acaricidal | |||||
| Tyrophagus putrescentiae Schrank | mite of stored food | MPEO menthol 55.2, menthone 19.5, isomenthone 7.3 | fumigant bioassay 0.63–80.0 μg/cm2 | ED50 2.72 μg/cm2 | [111] |
| filter paper bioassay 0.32–80.0 μg/ cm2 | ED50 1.87 μg/cm2 | ||||
| MAEO menthol 60.0, menthone 20.0 | fumigant bioassay 1.25–80.0 μg/cm2 | ED50 3.41 μg/cm2 | [112] | ||
| Weed/Crop Species | ED100 or ED50 | Main Compounds of EO and Their Content (%) | Ref. |
|---|---|---|---|
| Amaranthus retroflexus Avena sativa Avena fatua Brassica napus Bromus secalinus Centaurea cyanus Zea mays | ED50: 0.21 g/L ED50 0.53 ED50 0.72 ED50 0.63 ED50 0.16 ED50 0.06 ED50 1.03 | menthone 36.8, menthol 24.0 | [116] |
| Portulaca oleracea Lolium multiflorum Echinochloa crus-galli Maize Rice Tomato | 70% seeds germinated at a dose of 1 μL/mL ED100 0.125 μL/mL 72% seeds germinated at a dose of 1 μL/mL 6% seeds germinated at a dose of 1 μL/mL 58% seeds germinated at a dose of 1 μL/mL 3% seeds germinated at a dose of 1 μL/mL | menthone 23.3, iso-menthone 16.3, menthol 48.2 | [117] |
| Radish Tomato Jungle rice Convolvulus arvensis Portulaca oleracea | ED100 1.8 mL/L ED100 0.6 ED100 0.9 60% seeds germinated at a dose of 1.8 mL/L ED100 1.8 mL/L | menthol 35.1, menthone 17.5, menthofuran 11.7 | [118] |
| Amaranthus retroflexus Chenopodium album Lolium spp Portulaca oleracea Sinapis arvensis Solanum nigrum Vicia sativa | ED100 1.8 mg/L ED100 5.4 ED100 5.4 ED100 1.8 ED100 5.4 ED100 5.4 ED100 1.8 | No data | [119] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalemba, D.; Synowiec, A. Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis. Molecules 2020, 25, 59. https://doi.org/10.3390/molecules25010059
Kalemba D, Synowiec A. Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis. Molecules. 2020; 25(1):59. https://doi.org/10.3390/molecules25010059
Chicago/Turabian StyleKalemba, Danuta, and Agnieszka Synowiec. 2020. "Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis" Molecules 25, no. 1: 59. https://doi.org/10.3390/molecules25010059
APA StyleKalemba, D., & Synowiec, A. (2020). Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis. Molecules, 25(1), 59. https://doi.org/10.3390/molecules25010059