Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium barbarum L. Fruit

 ,
,
Abstract
1. Introduction
2. Results
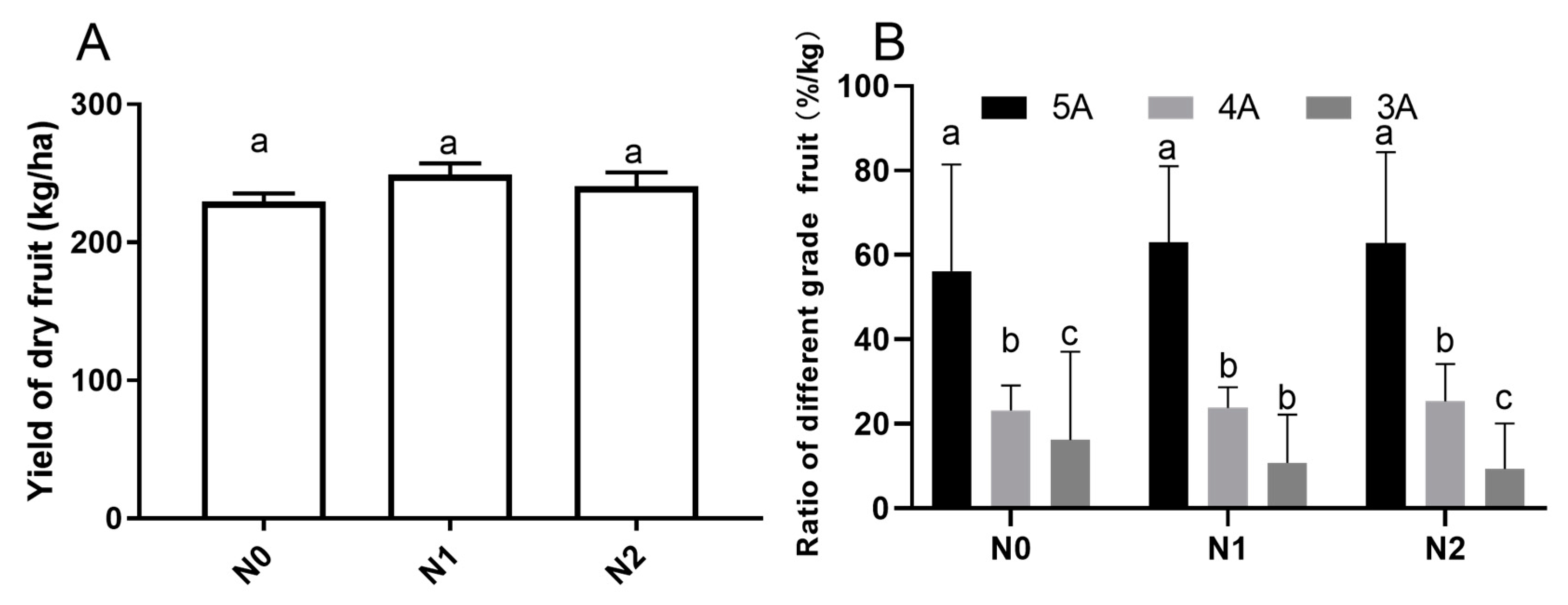
2.1. Yield and Commodity Grade of Goji Dry Fruits under Different Nitrogen Fertilizations
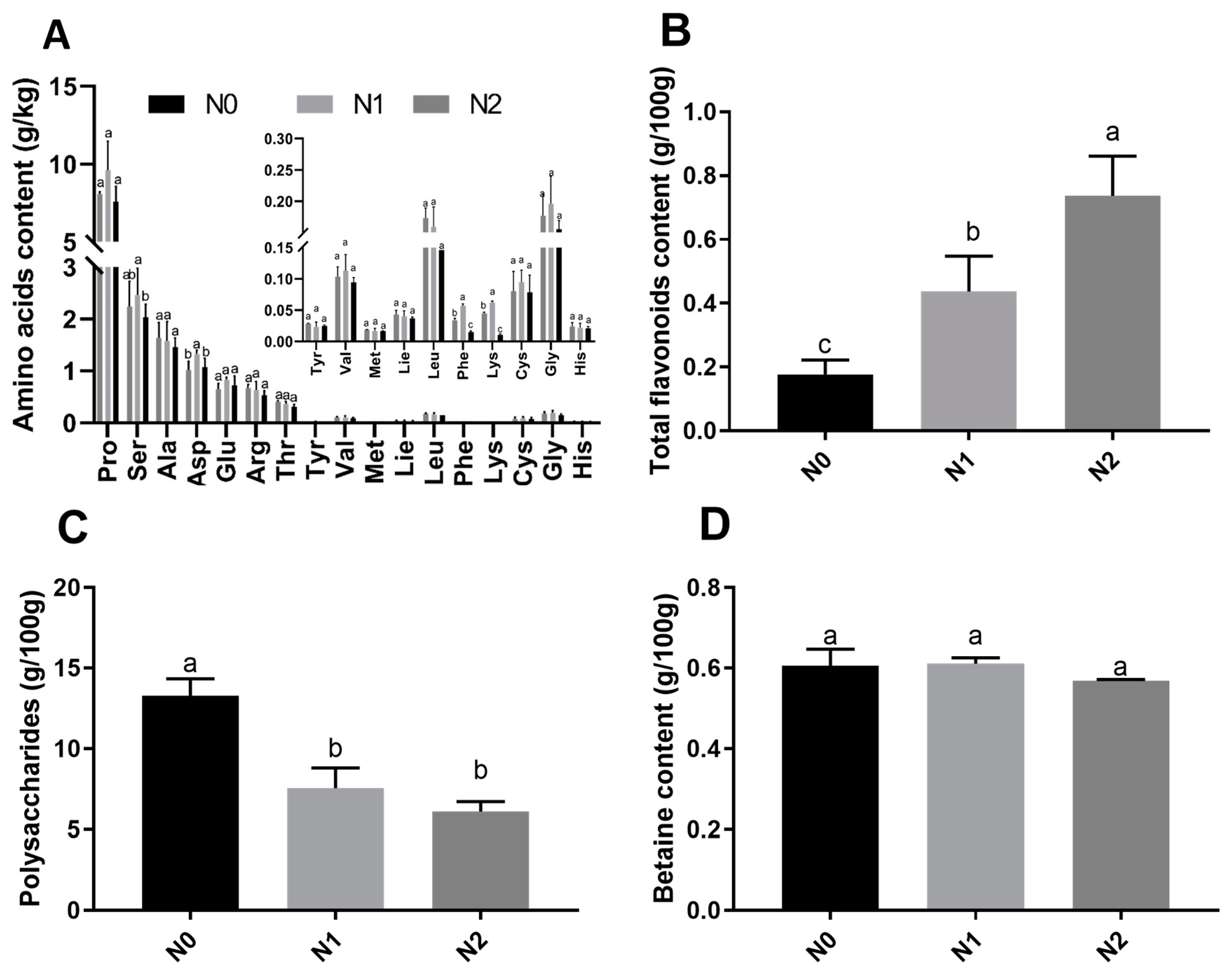
2.2. Nutritional Contents of Goji Fruits under Different Nitrogen Fertilizations
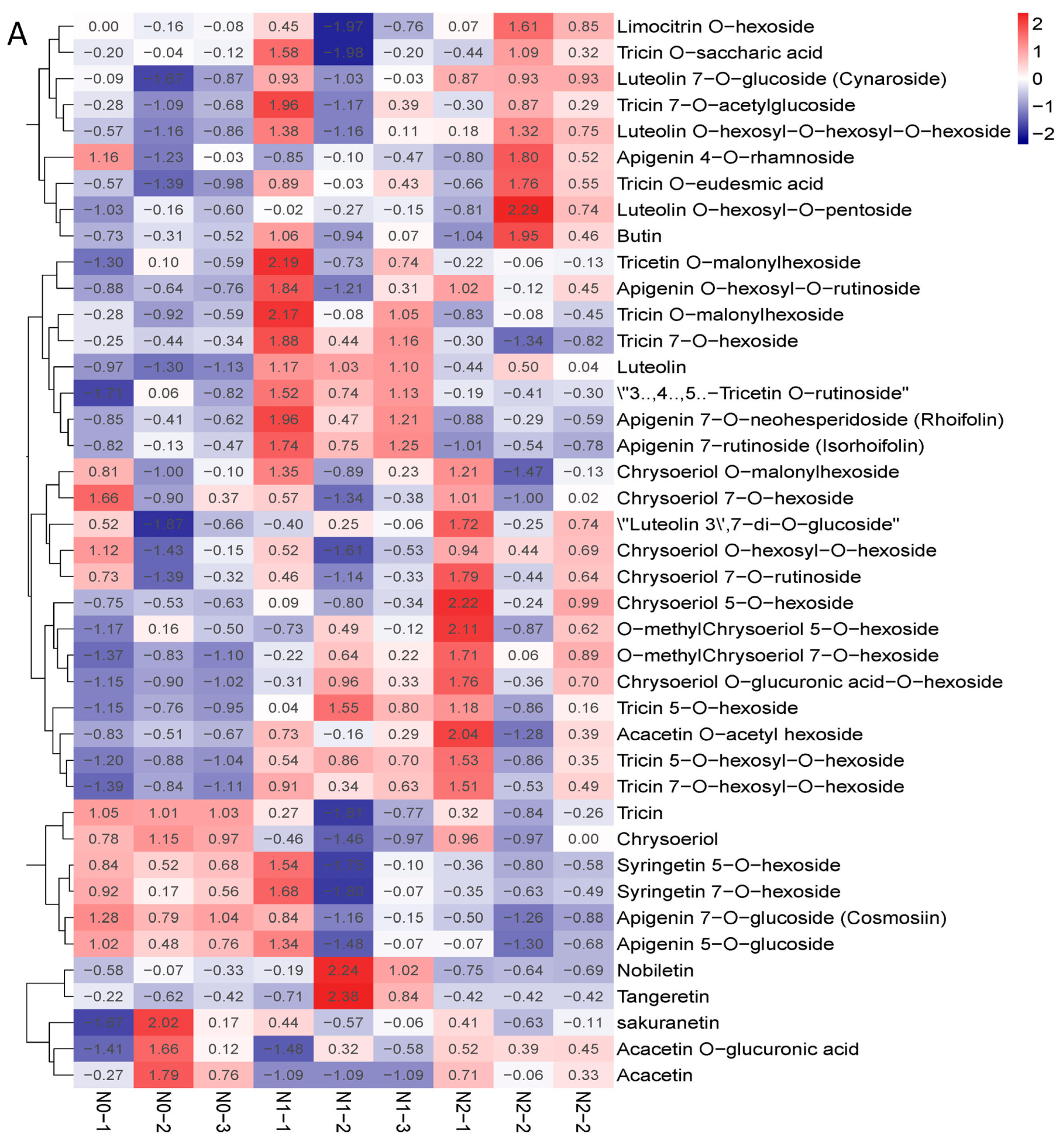
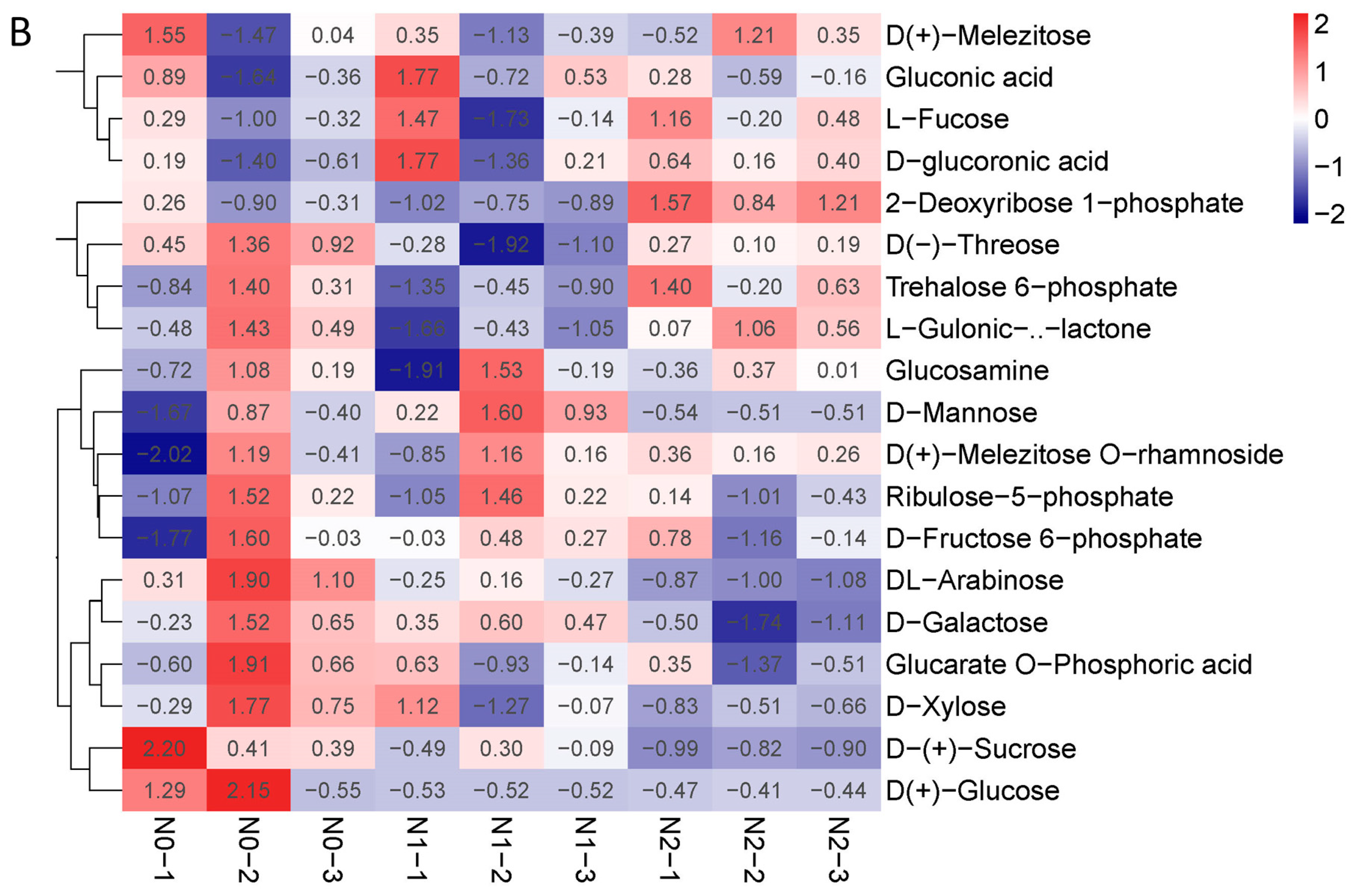
2.3. Metabolic Profiling of Goji Fruits under Different Nitrogen Fertilizations
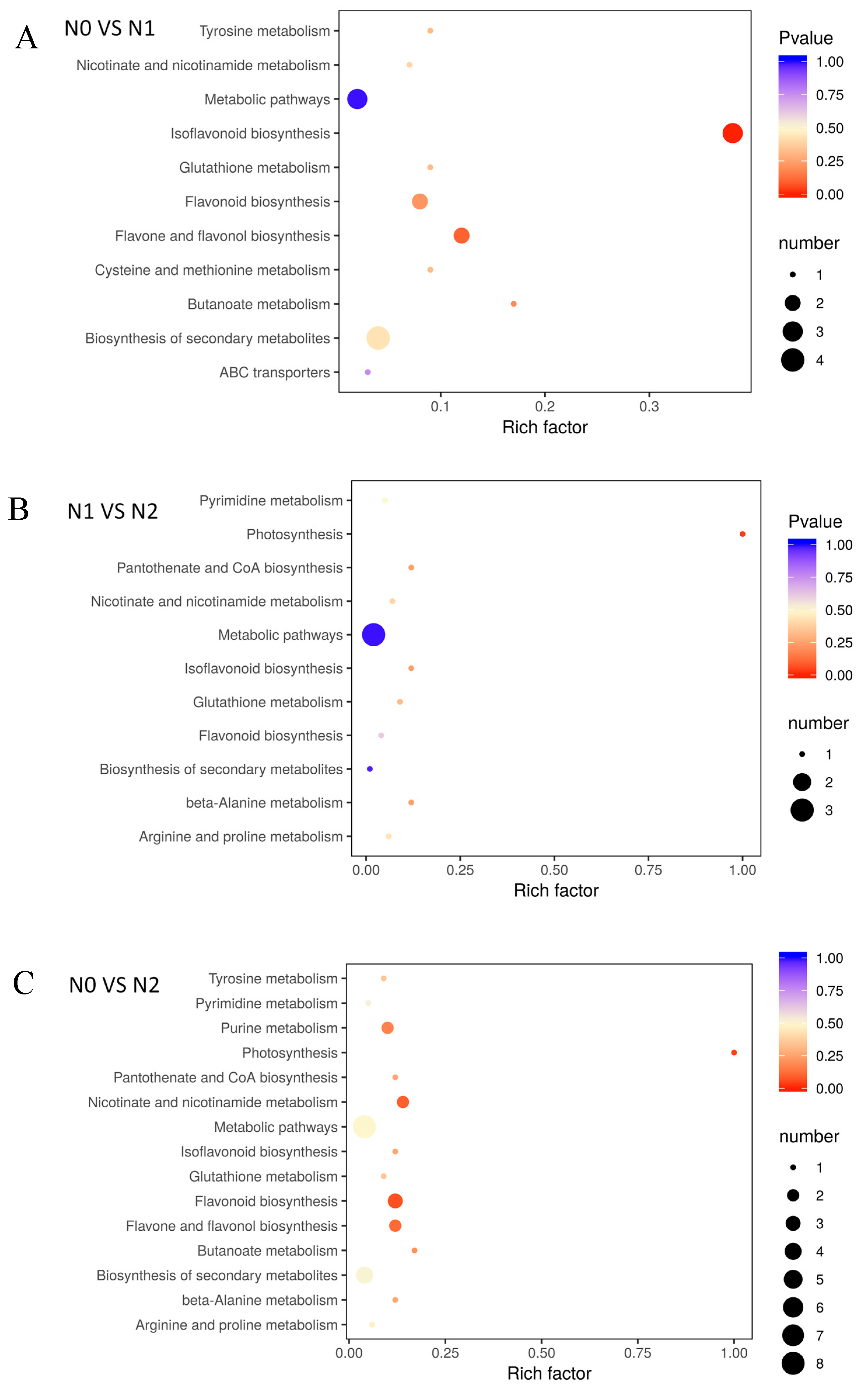
2.4. Metabolite of Goji Fruits under Different Nitrogen Fertilizations, Functional Annotation, and Enrichment Analysis of KEGG
3. Discussion
3.1. Effects of Different Nitrogen fertilizations on Goji Fruit Commodity Grade and Yield
3.2. Effects of Different Nitrogen Fertilizations on Fruit Nutrient Content of Goji
3.3. Metabolic Profiling of Goji Fruits under Different Nitrogen fertilizations
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Design and Soil Conditions
4.3. Determination of Primary Nutrients of the Goji Fruit
4.3.1. Determination of Total Flavonoid, Polysaccharide, and Betaine Contents
4.3.2. Determination of the Amino Acid Content
4.4. Sample Preparation and Extraction
4.5. HPLC Conditions
4.6. ESI-Q TRAP-MS/MS
4.7. Qualitative and Quantitative Analyses of Metabolites
4.8. Sample Quality Control Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium barbarum: A traditional Chinese herb and a promising anti-aging agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Amagase, H.; Farnsworth, N.R. A review of botanical characteristics, phytochemistry, clinical relevance in efficacy and safety of Lycium barbarum fruit (Goji). Food Res. Int. 2011, 44, 1702–1717. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Zgola-Grzeskowiak, A.; Stanisz, E.; Waskiewicz, A. Potential health benefits and quality of dried fruits: Goji fruits, cranberries and raisins. Food Chem. 2017, 221, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.; Zhao, Y.; Yang, G.; Huang, L. Systematic review of chemical constituents in the genus Lycium (Solanaceae). Molecules 2017, 22, 911. [Google Scholar] [CrossRef] [PubMed]
- Yossa Nzeuwa, I.B.; Guo, B.; Zhang, T.; Wang, L.; Ji, Q.; Xia, H.; Sun, G. Comparative metabolic profiling of Lycium fruits (Lycium barbarum and Lycium chinense) from different areas in China and from Nepal. J. Food Qual. 2019, 1, 1–6. [Google Scholar] [CrossRef]
- Lu, Y.; Guo, S.; Zhang, F.; Yan, H.; Qian, D.W.; Wang, H.Q.; Jin, L.; Duan, J.A. Comparison of Functional Components and Antioxidant Activity of Lycium barbarum L. Fruits from Different Regions in China. Molecules 2019, 24, 2228. [Google Scholar] [CrossRef]
- Liu, Y.L.; Zeng, S.H.; Sun, W.; Wu, M.; Hu, W.M.; Shen, X.F.; Wang, Y. Comparative analysis of carotenoid accumulation in two Goji (Lycium barbarum L. and Ruthenicum murr. L.) fruits. BMC Plant Biol. 2014, 14, 269. [Google Scholar] [CrossRef]
- Chung, R.S.; Chen, C.C.; Ng, L.T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crop Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Kaplan, M.; Karaman, K.; Kardes, Y.M.; Kale, H. Phytic acid content and starch properties of maize (Zea mays L.): Effects of irrigation process and nitrogen fertilizer. Food Chem. 2019, 283, 375–380. [Google Scholar] [CrossRef]
- Calatrava, V.; Hom, E.F.Y.; Llamas, Á.; Fernández, E.; Galván, A. Nitrogen scavenging from amino acids and peptides in the model alga Chlamydomonas reinhardtii. The role of extracellular l-amino oxidase. Algal Res. 2019, 38, 101395. [Google Scholar] [CrossRef]
- López-Carmona, D.A.; Alarcón, A.; Martínez-Romero, E.; Peña-Cabriales, J.J.; Larsen, J. Maize plant growth response to whole rhizosphere microbial communities in different mineral N and P fertilization scenarios. Rhizosphere 2019, 9, 38–46. [Google Scholar] [CrossRef]
- Noushina, I.; Shahid, U.; Nafees, A.K. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar]
- Xiong, Z.X.; Chen, G.S.; Chen, H.M. Study on nitrogen absorption of Lycium barbarum L. Special Wild Econ. Plant Res. 1991, 6, 16–17. [Google Scholar]
- Cai, G.J.; Zhang, G.Z.; Zhang, B.L.; Wang, S.Y.; Zhong, L.L.; Wu, L.; Hu, B.F.; Wei, Q. Effect of n, p, k on Chinese wolfberry fruit sugar and carotene content. Southwest China J. Agric. Sci. 2013, 26, 209–212. [Google Scholar]
- Jianhong, K.; Hongliang, W.; Juan, Y.; Jiantao, Y. Study on the relationship between the main secondarymetabolites and polysaccharide in fruits of Lycium barbarum at different application amounts of nitrogen. Agric. Sci. Tehnol. 2008, 36, 16008–16010. [Google Scholar]
- Liu, J.W.; Zheng, G.Q.; Liu, G.H.; Yang, J.D.; Xu, X. Different application amounts of phosphorus on commodity quality and main effective components of Lycium barbarum L. North. Hortic. 2016, 14, 167–171. [Google Scholar]
- Shi, Z.G.; Wei, F.; Wan, R.; Wang, Y.J.; An, W.; Qin, K. Effect of different fertilizeation levels on N, P, K contents and resistance of “0909” wolfberry leaves. North. Hortic. 2016, 21, 165–170. [Google Scholar]
- Chen, C.; Xu, M.; Wang, C.; Qiao, G.; Wang, W.; Tan, Z.; Wu, T.; Zhang, Z. Characterization of the Lycium barbarum fruit transcriptome and development of EST-SSR markers. PLoS ONE 2017, 12, e0187738. [Google Scholar] [CrossRef]
- Dong, J.Z.; Lu, D.Y.; Wang, Y. Analysis of flavonoids from leaves of cultivated Lycium barbarum L. Plant Foods Hum. Nutr. 2009, 64, 199–204. [Google Scholar] [CrossRef]
- Islam, T.; Yu, X.; Badwal, T.S.; Xu, B. Comparative studies on phenolic profiles, antioxidant capacities and carotenoid contents of red Goji berry (Lycium barbarum) and black Goji berry (Lycium ruthenicum). Chem. Central J. 2017, 11, 59. [Google Scholar] [CrossRef]
- Lin, N.C.; Lin, J.C.; Chen, S.H.; Ho, C.T.; Yeh, A.I. Effect of Goji (Lycium barbarum) on expression of genes related to cell survival. J. Agric. Food Chem. 2011, 59, 10088–10096. [Google Scholar] [CrossRef] [PubMed]
- Patsilinakos, A.; Ragno, R.; Carradori, S.; Petralito, S.; Cesa, S. Carotenoid content of Goji berries: CIELAB, HPLC-DAD analyses and quantitative correlation. Food Chem. 2018, 268, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Mocan, A.; Moldovan, C.; Zengin, G.; Bender, O.; Locatelli, M.; Simirgiotis, M.; Atalay, A.; Vodnar, D.C.; Rohn, S.; Crisan, G. UHPLC-QTOF-MS analysis of bioactive constituents from two Romanian Goji (Lycium barbarum L.) berries cultivars and their antioxidant, enzyme inhibitory, and real-time cytotoxicological evaluation. Food Chem Toxicol. 2018, 115, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Q.; Xiao, J.; Fan, H.X.; Yu, Y.; He, R.R.; Feng, X.L.; Kurihara, H.; So, K.F.; Yao, X.S.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Patra, J.K.; Ali, A.; Maqbool, M.; Arshad, M.I. Effect of compost and NPK fertilizer on improving biochemical and antioxidant properties of Moringa oleifera. South Afr. J. Bot. 2019. [Google Scholar] [CrossRef]
- Dong, S.D.; Qin, K.; Wan, S.Q.; Kang, H.Y.; Zhou, W.Y. Effect of different micro-irrigation methods on soil moisture, nutrients distribution and the yield of Lycium barbarbum L. Water Sav. Irrig. 2017, 38–43. (In Chinese) [Google Scholar]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araujo, W.L.; Braun, H.P. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Zhen, S.; Zhou, J.; Deng, X.; Zhu, G.; Cao, H.; Wang, Z.; Yan, Y. Metabolite profiling of the response to high-nitrogen fertilizer during grain development of bread wheat (Triticum aestivum L.). J. Cereal Sci. 2016, 69, 85–94. [Google Scholar] [CrossRef]
- Hernandez-Sebastia, C.; Marsolais, F.; Saravitz, C.; Israel, D.; Dewey, R.E.; Huber, S.C. Free amino acid profiles suggest a possible role for asparagine in the control of storage-product accumulation in developing seeds of low- and high-protein soybean lines. J. Exp. Bot. 2005, 56, 1951–1963. [Google Scholar] [CrossRef]
- Watanabe, M.; Ohta, Y.; Licang, S.; Motoyama, N.; Kikuchi, J. Profiling contents of water-soluble metabolites and mineral nutrients to evaluate the effects of pesticides and organic and chemical fertilizers on tomato fruit quality. Food Chem. 2015, 169, 387–395. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, M.; Liu, J.; Wang, W.; Liu, S. Fertilizer and soil nitrogen utilization of pear trees as affected by the timing of split fertilizer application in rain-fed orchard. Sci. Hortic. 2019, 252, 363–369. [Google Scholar] [CrossRef]
- Novoa, R.; Loomis, R.S. Nitrogen and plant production. Plant Soil 1981, 58, 177–204. [Google Scholar] [CrossRef]
- Kokotkiewicz, A.; Migas, P.; Stefanowicz, J.; Luczkiewicz, M.; Krauze-Baranowska, M. Densitometric TLC analysis for the control of tropane and steroidal alkaloids in Lycium barbarum. Food Chem. 2017, 221, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Wiedenmann, M.; Tittel, G.; Bauer, R. HPLC-MS trace analysis of atropine in Lycium barbarum berries. Phytochem. Anal. 2006, 17, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Guo, S.; Yan, H.; Lu, Y.Y.; Zhang, F.; Qian, D.W.; Wang, H.Q.; Duan, J.A. Analysis of phenolic acids and flavonoids in leaves of Lycium barbarum from different habitats by ultra-high performance liquid chromatography coupled with triple quadrupole tandem mass spectrometry. Biomed. Chromatogr. 2019, 33, e4552. [Google Scholar] [CrossRef] [PubMed]
- Bassard, J.E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, P.; Yang, Q.; Gong, X.; Ma, H.; Dang, K.; Chen, G.; Gao, X.; Feng, B. Analysis of Flavonoid Metabolites in Buckwheat Leaves Using UPLC-ESI-MS/MS. Molecules 2019, 24, 1310. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Wang, A.; Li, R.; Ren, L.; Gao, X.; Zhang, Y.; Ma, Z.; Ma, D.; Luo, Y. A comparative metabolomics study of flavonoids in sweet potato with different flesh colors (Ipomoea batatas (L.) Lam). Food Chem. 2018, 260, 124–134. [Google Scholar] [CrossRef]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Wang, S.; Tu, H.; Wan, J.; Chen, W.; Liu, X.; Luo, J.; Xu, J.; Zhang, H. Spatio-temporal distribution and natural variation of metabolites in citrus fruits. Food Chem. 2016, 199, 8–17. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Nitrogen g/plant | Phosphorus g/plant | Potassium g/plant |
|---|---|---|---|
| N0 | 42.5 | 65 | 50 |
| N1 | 85 | 65 | 50 |
| N2 | 127.5 | 65 | 50 |
| Time (min) | Mobile Phase (A) % | Mobile Phase (B) % |
|---|---|---|
| 0 | 100 | 0 |
| 14 | 85 | 15 |
| 29 | 66 | 34 |
| 30 | 0 | 100 |
| 37 | 0 | 100 |
| 38 | 100 | 0 |
| 45 | 100 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Wei, F.; Wan, R.; Li, Y.; Wang, Y.; An, W.; Qin, K.; Dai, G.; Cao, Y.; Feng, J. Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium barbarum L. Fruit. Molecules 2019, 24, 3879. https://doi.org/10.3390/molecules24213879
Shi Z, Wei F, Wan R, Li Y, Wang Y, An W, Qin K, Dai G, Cao Y, Feng J. Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium barbarum L. Fruit. Molecules. 2019; 24(21):3879. https://doi.org/10.3390/molecules24213879
Chicago/Turabian StyleShi, Zhigang, Feng Wei, Ru Wan, Yunxiang Li, Yajun Wang, Wei An, Ken Qin, Guoli Dai, Youlong Cao, and Jiayue Feng. 2019. "Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium barbarum L. Fruit" Molecules 24, no. 21: 3879. https://doi.org/10.3390/molecules24213879
APA StyleShi, Z., Wei, F., Wan, R., Li, Y., Wang, Y., An, W., Qin, K., Dai, G., Cao, Y., & Feng, J. (2019). Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium barbarum L. Fruit. Molecules, 24(21), 3879. https://doi.org/10.3390/molecules24213879