β-Asarone Induces Apoptosis and Cell Cycle Arrest of Human Glioma U251 Cells via Suppression of HnRNP A2/B1-Mediated Pathway In Vitro and In Vivo

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
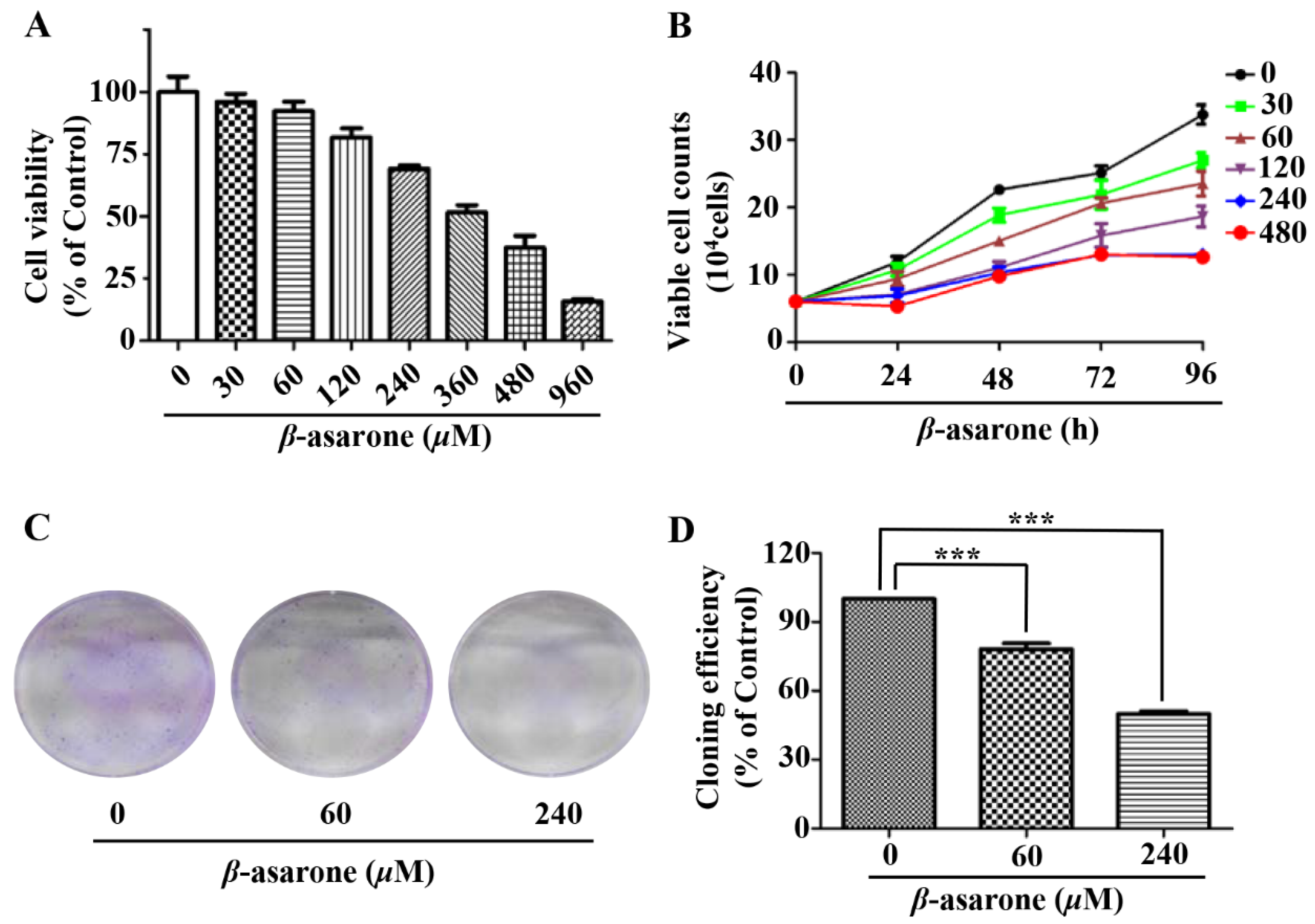
2.1. β-Asarone Inhibited the Growth of U251 Cells
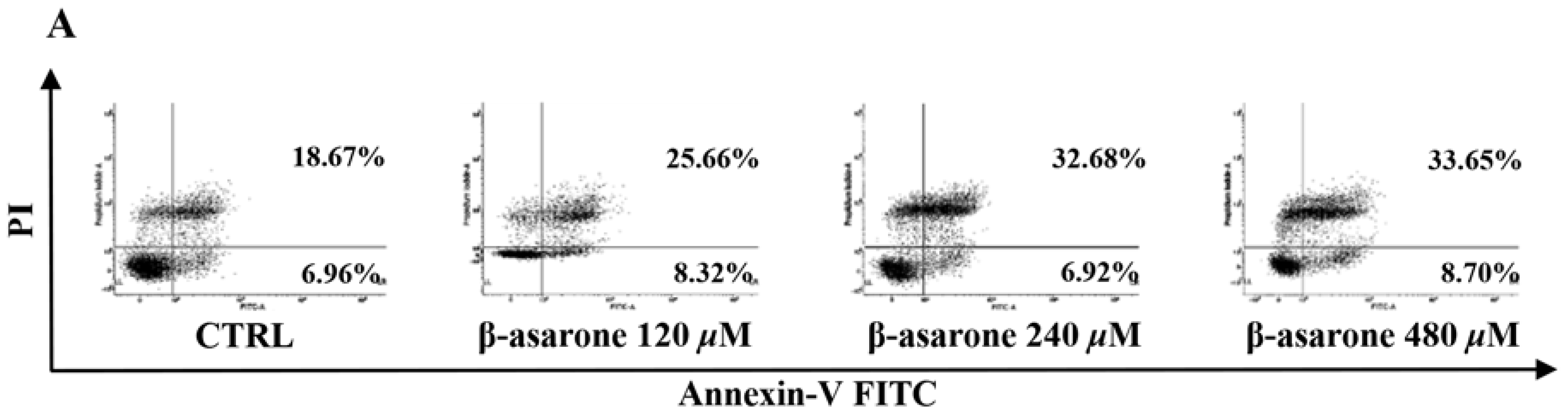
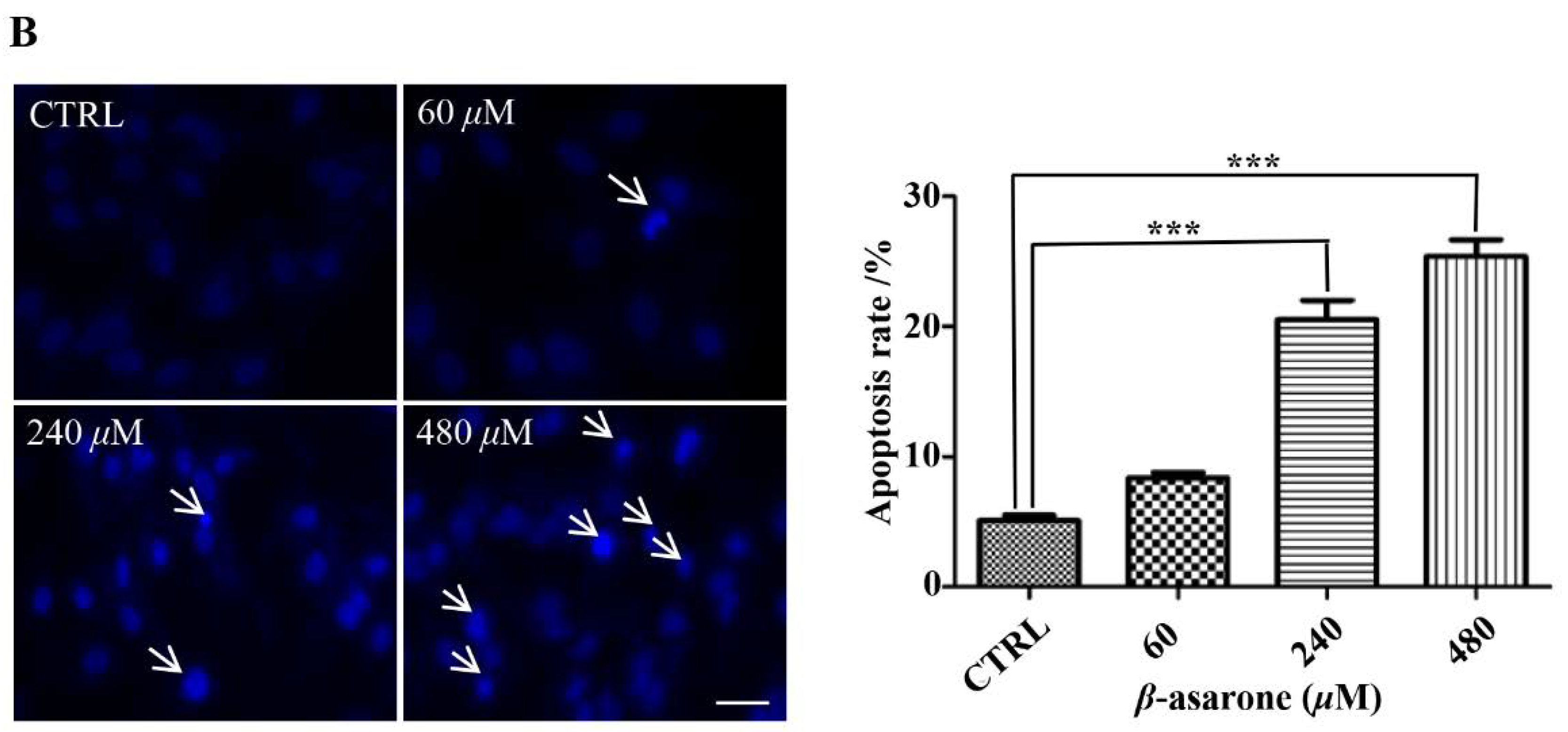
2.2. β-Asarone Induced Apoptosis of U251 Cells
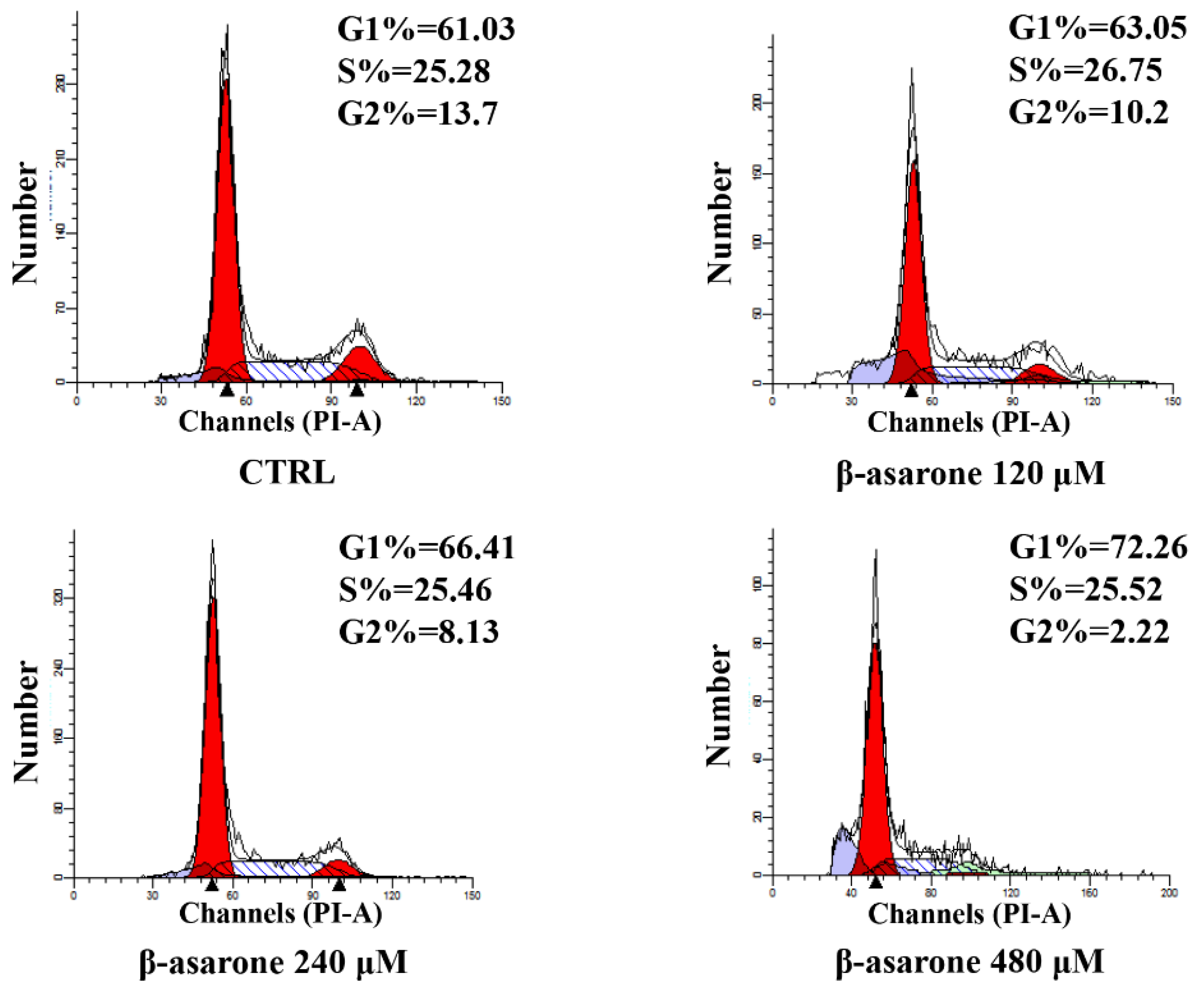
2.3. β-Asarone Induced Cell Cycle Arrest of U251 Cells
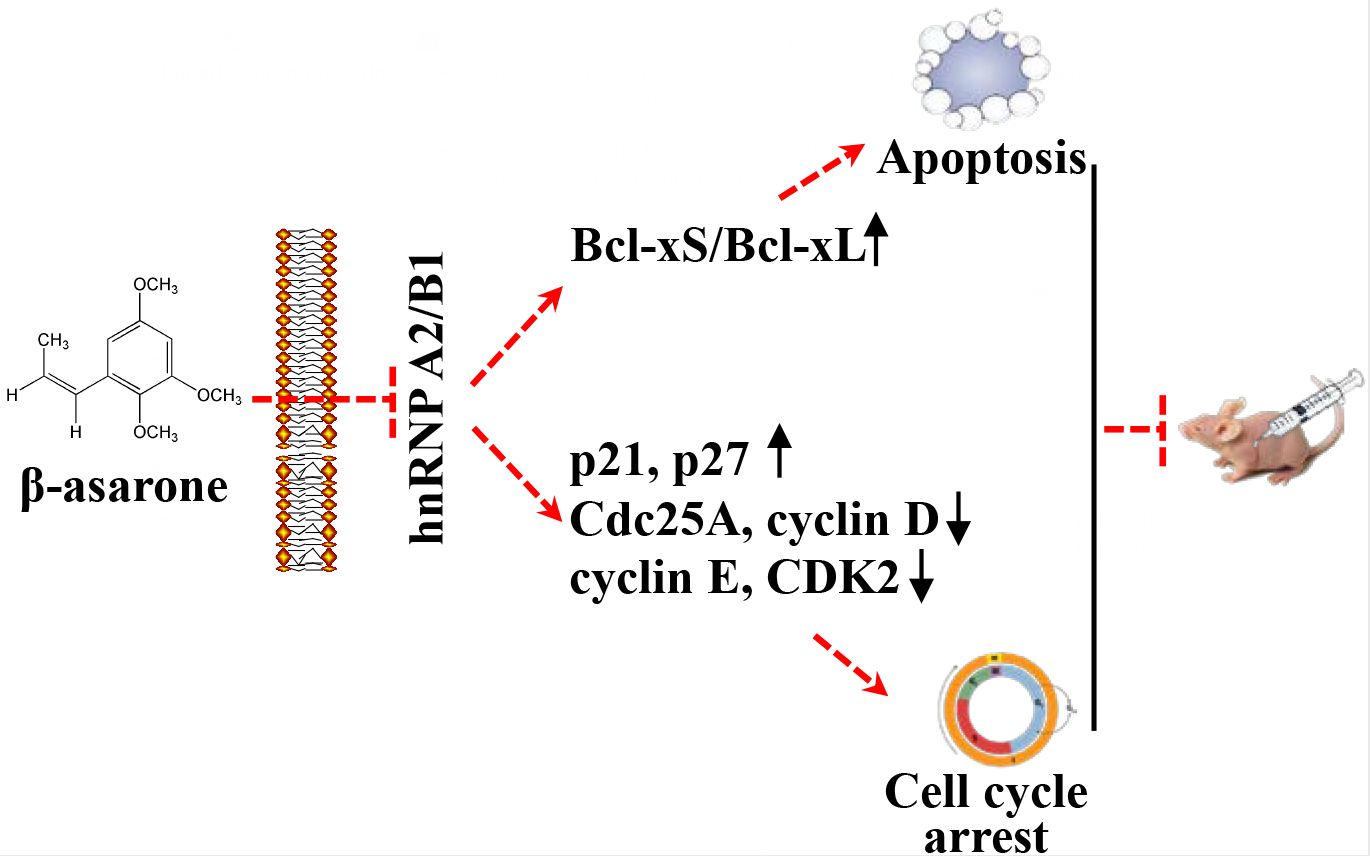
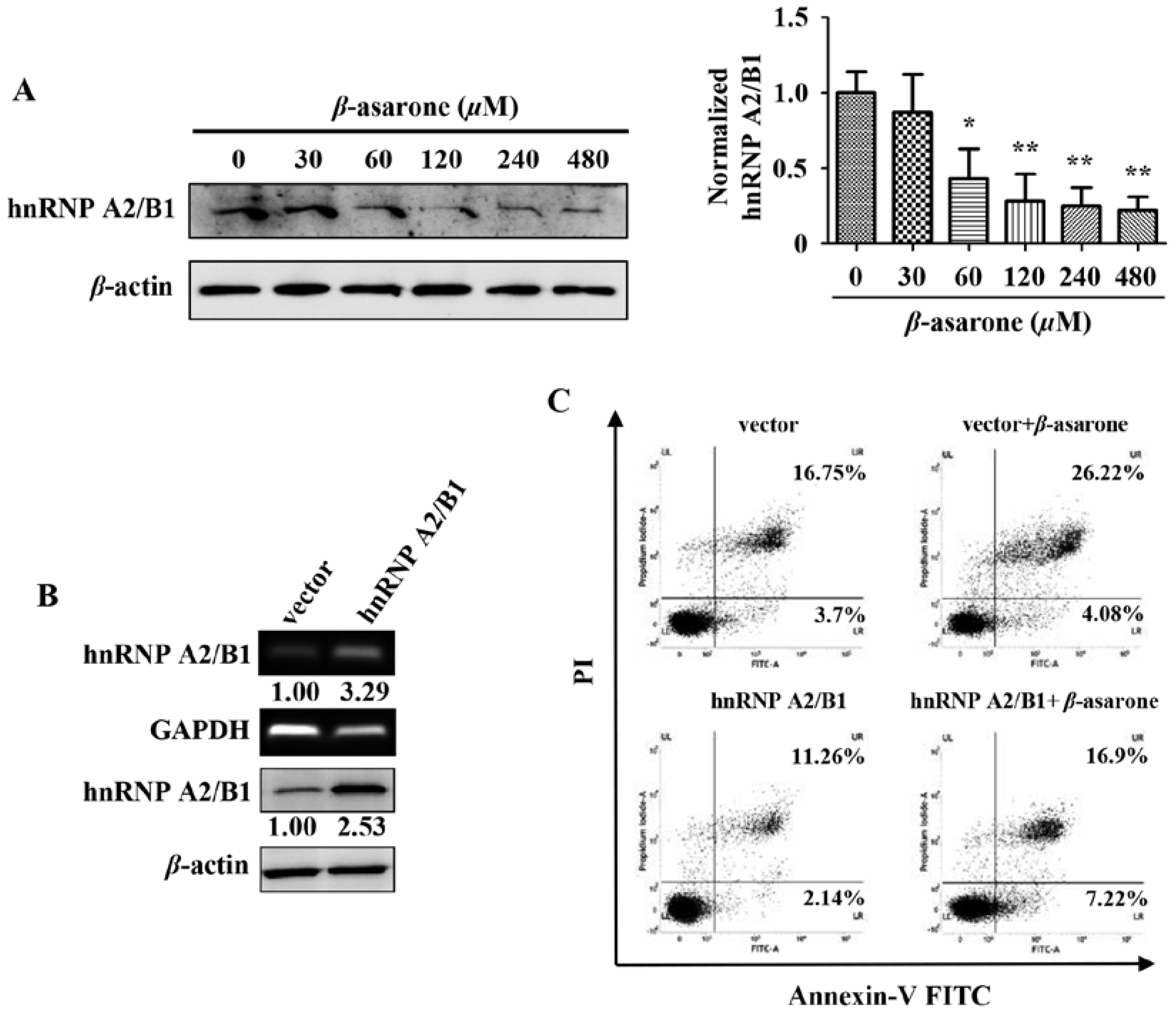
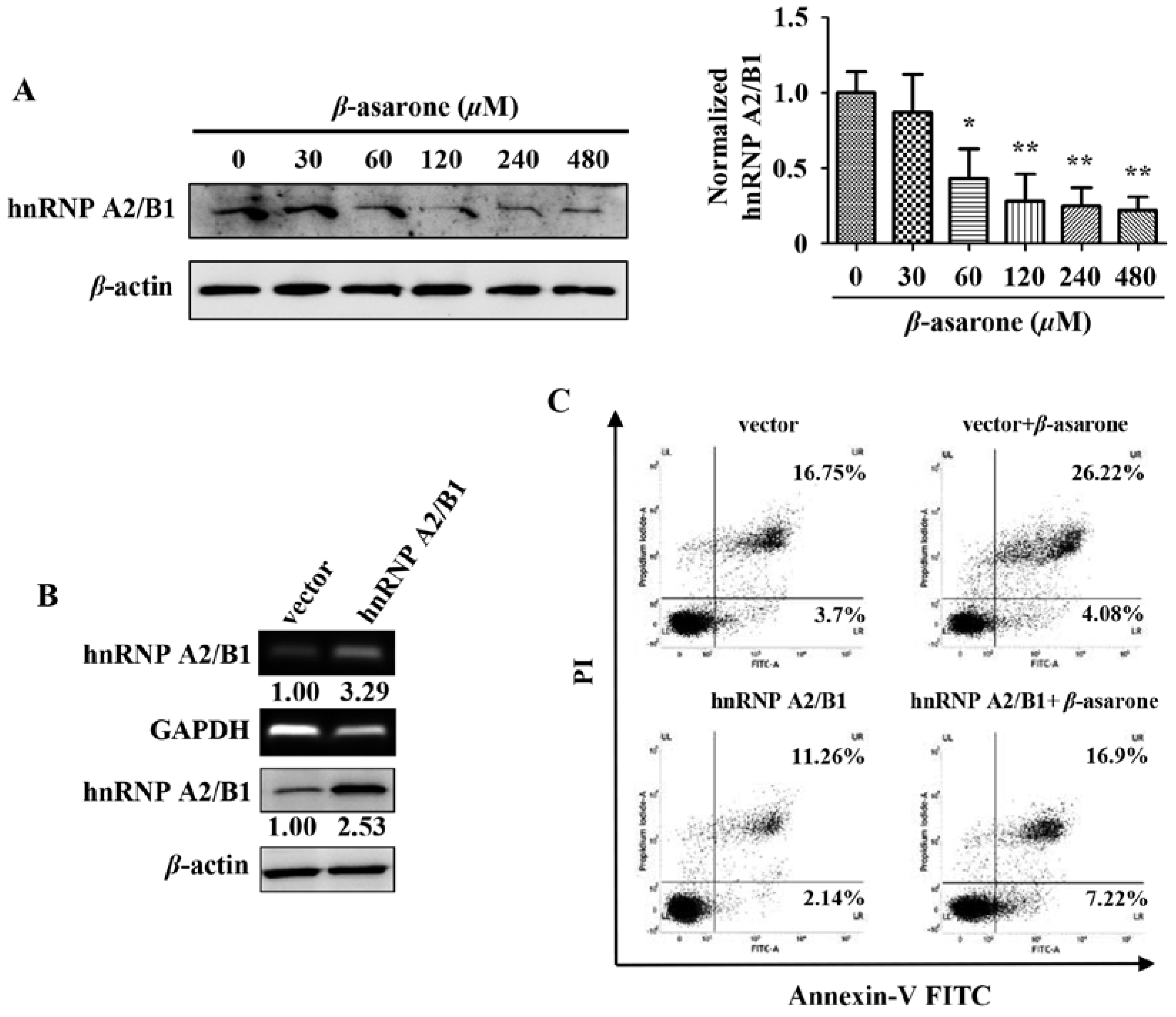
2.4. Suppression of HnRNP A2/B1 Contributes to β-Asarone-Induced Apoptosis
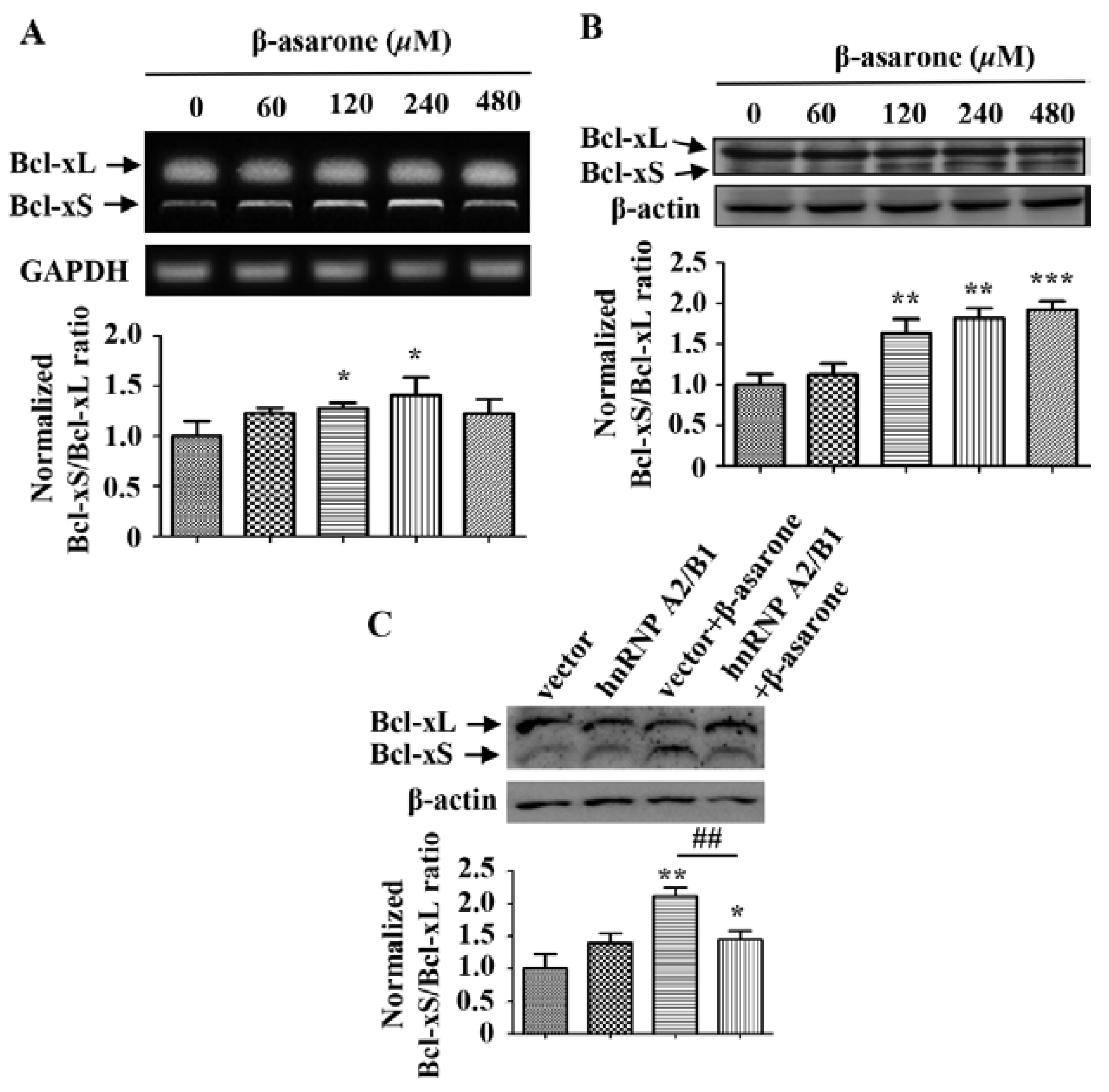
2.5. β-Asarone Promoted the Splicing of Bcl-x via the Inhibition of HnRNP A2/B1
2.6. Cell Death Receptor Pathway and Mitochondrial Pathway Are Involved in the β-Asarone-Induced Apoptosis of U251 Cells
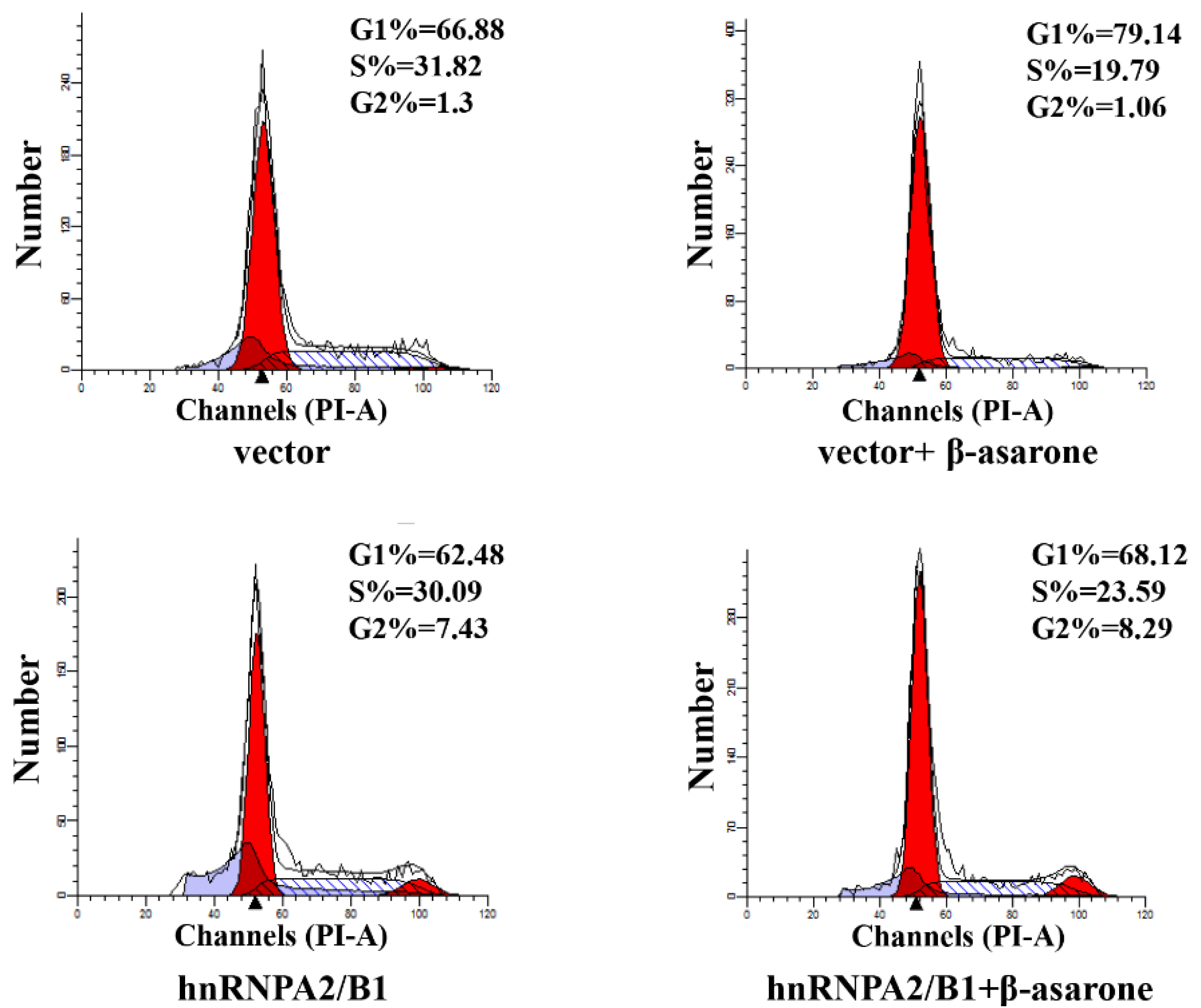
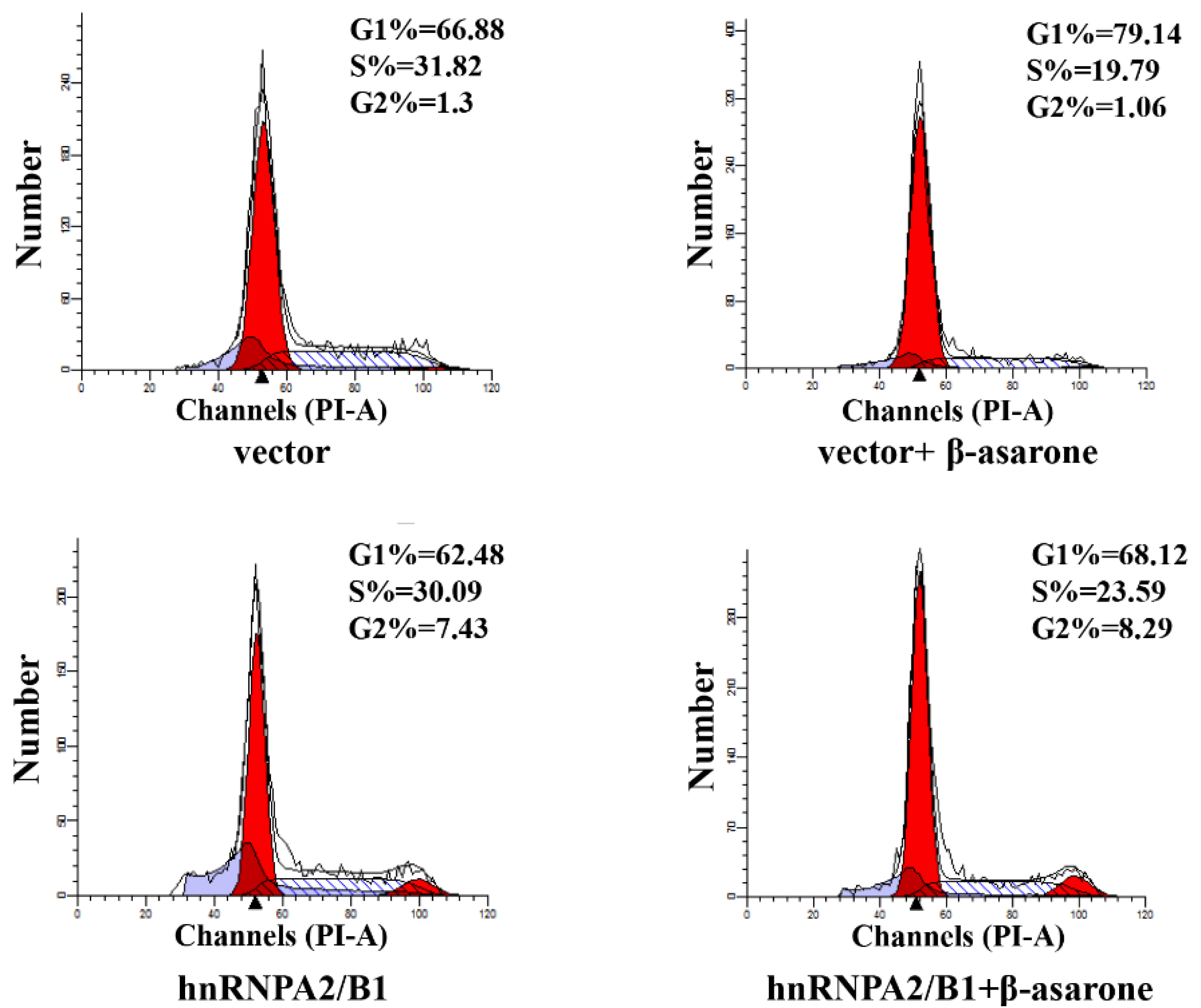
2.7. Suppression of HnRNP A2/B1 Contributes to β-Asarone-Induced Cell Cycle Arrest
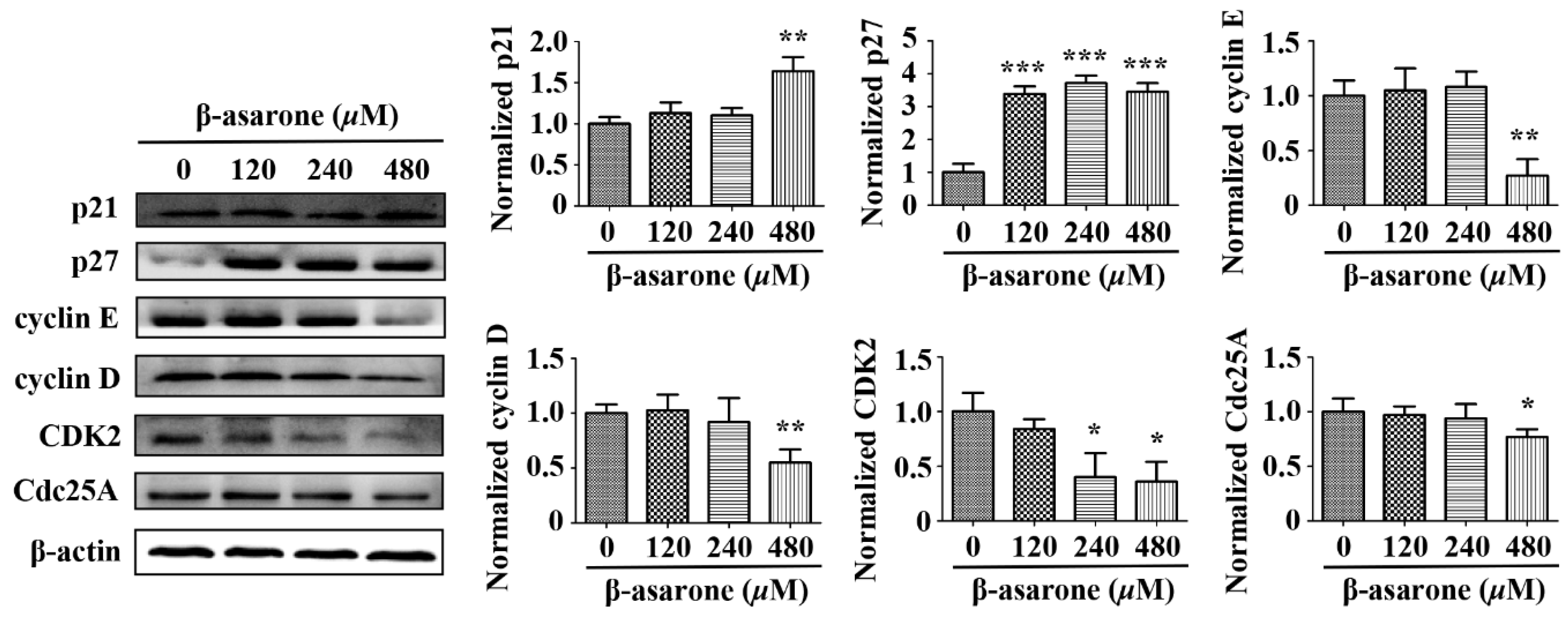
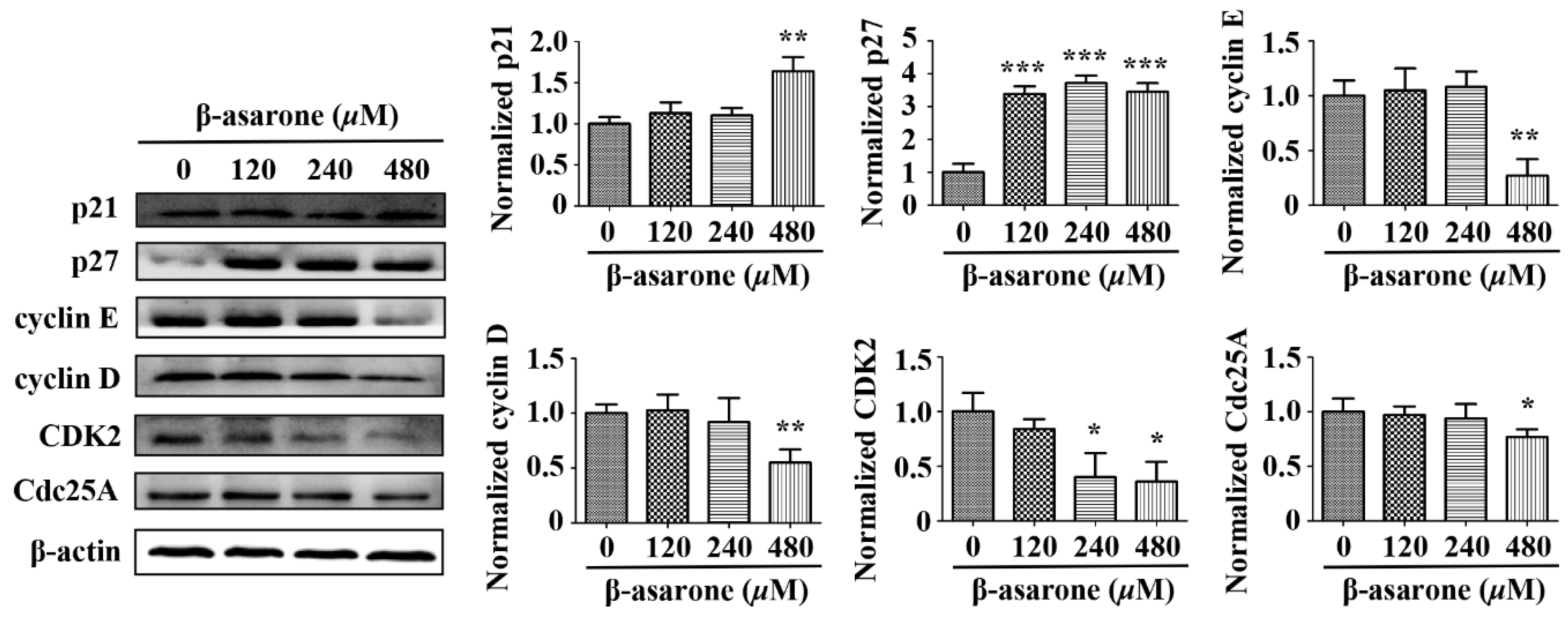
2.8. β-Asarone Modulated the Expression of Cell Cycle Related Proteins in U251 Cells
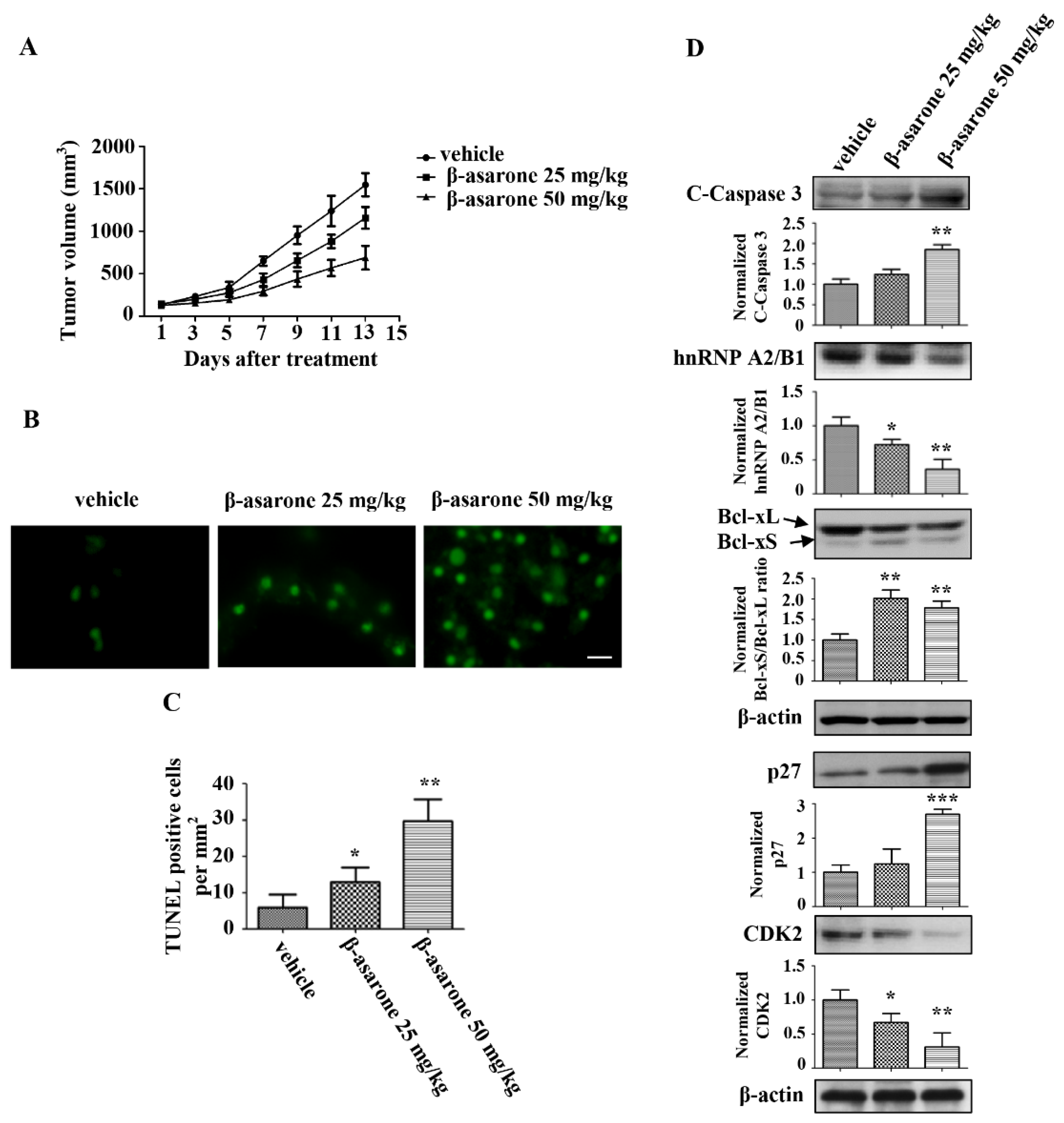
2.9. β-Asarone Suppressed the Tumor Growth of U251 Xenografts in Nude Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Trypan Blue Dye Exclusion Test
4.5. Colony Formation Assay
4.6. Cell Apoptosis Analyzed by Flow Cytometry
4.7. Hoechst 33342 Staining
4.8. Western Blotting Analysis
4.9. Cell Cycle Distribution Analyzed by Flow Cytometry
4.10. Plasmids and Transient Transfection
4.11. Semi-Quantitative RT-PCR Detection
4.12. Nude Mouse Xenograft Model
4.13. Terminal Dexynucleotidyl Transferase-Mediated dUTP Nick End Labeling (TUNEL) Assay
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| hnRNPs | Heterogeneous nuclear ribonucleoproteins |
| BBB | blood–brain barrier |
| SRB | Sulforhodamine B |
| DMSO | dimethyl sulfoxide |
| TUNEL | Terminal Dexynucleotidyl Transferase-Mediated dUTP Nick End Labeling |
| C-Caspase 8 | Cleaved-Caspase 8 |
| C-BID | Cleaved-BID |
| C-Caspase 3 | Cleaved-Caspase 3 |
| CDKs | cyclin-dependent kinases |
| CKIs | cyclins and cyclin-dependent kinase inhibitors |
References
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Van Tellingen, O.; Yetkin-Arik, B.; de Gooijer, M.C.; Wesseling, P.; Wurdinger, T.; de Vries, H.E. Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updates 2015, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Han, S.P.; Tang, Y.H.; Smith, R. Functional diversity of the hnRNPs: Past, present and perspectives. Biochem. J. 2010, 430, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Golan-Gerstl, R.; Cohen, M.; Shilo, A.; Suh, S.S.; Bakacs, A.; Coppola, L.; Karni, R. Splicing factor hnRNP A2/B1 regulates tumor suppressor gene splicing and is an oncogenic driver in glioblastoma. Cancer Res. 2011, 71, 4464–4472. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Allred, D.C.; Avis, I.; Martinez, A.; Vos, M.D.; Smith, L.; Treston, A.M.; Mulshine, J.L. Differential expression of the early lung cancer detection marker, heterogeneous nuclear ribonucleoprotein-A2/B1 (hnRNP-A2/B1) in normal breast and neoplastic breast cancer. Breast Cancer Res. Treat. 2001, 66, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Yan-Sanders, Y.; Hammons, G.J.; Lyn-Cook, B.D. Increased expression of heterogeneous nuclear ribonucleoprotein A2/B1 (hnRNP) in pancreatic tissue from smokers and pancreatic tumor cells. Cancer Lett. 2002, 183, 215–220. [Google Scholar] [CrossRef]
- Lee, C.L.; Hsiao, H.H.; Lin, C.W.; Wu, S.P.; Huang, S.Y.; Wu, C.Y.; Wang, A.H.; Khoo, K.H. Strategic shotgun proteomics approach for efficient construction of an expression map of targeted protein families in hepatoma cell lines. Proteomics 2003, 3, 2472–2486. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lum, J.H.; Cheung, B.P.; Wong, M.S.; Butt, Y.K.; Tam, M.F.; Chan, W.Y.; Chow, C.; Hui, P.K.; Kwok, F.S.; et al. Identification of the heterogeneous nuclear ribonucleoprotein A2/B1 as the antigen for the gastrointestinal cancer specific monoclonal antibody MG7. Proteomics 2005, 5, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Dowling, P.; Pollard, D.; Larkin, A.; Henry, M.; Meleady, P.; Gately, K.; O’Byrne, K.; Barr, M.P.; Lynch, V.; Ballot, J.; et al. Abnormal levels of heterogeneous nuclear ribonucleoprotein A2B1 (hnRNPA2B1) in tumour tissue and blood samples from patients diagnosed with lung cancer. Mol. Biosyst. 2015, 11, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Chen, S.; Wang, F.; Zhao, H.; Xie, Z.; Xu, Z.; Zhang, Q.; Liang, P.; Zhai, X.; Cheng, Y. Effects of hnRNP A2/B1 Knockdown on Inhibition of Glioblastoma Cell Invasion, Growth and Survival. Mol. Neurobiol. 2016, 53, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- Shilo, A.; Siegfried, Z.; Karni, R. The role of splicing factors in deregulation of alternative splicing during oncogenesis and tumor progression. Mol. Cell. Oncol. 2015, 2, e970955. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Gupta, S.C.; Peng, W.X.; Zhou, N.; Pochampally, R.; Atfi, A.; Watabe, K.; Lu, Z.; Mo, Y.Y. Regulation of alternative splicing of Bcl-x by BC200 contributes to breast cancer pathogenesis. Cell Death Dis. 2016, 7, e2262. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Cai, L.; Zhu, J.; Chen, M.; Chen, J.; Li, Z.H.; Liu, X.D.; Wang, S.G.; Bie, P.; Jiang, P.; et al. Fyn requires HnRNPA2B1 and Sam68 to synergistically regulate apoptosis in pancreatic cancer. Carcinogenesis 2011, 32, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Paronetto, M.P.; Achsel, T.; Massiello, A.; Chalfant, C.E.; Sette, C. The RNA-binding protein Sam68 modulates the alternative splicing of Bcl-x. J. Cell Biol. 2007, 176, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Ran, L.; Liu, Y.; Zhong, S.H.; Zhou, P.P.; Liao, M.X.; Fang, W. Knockdown of hnRNP A2/B1 inhibits cell proliferation, invasion and cell cycle triggering apoptosis in cervical cancer via PI3K/AKT signaling pathway. Oncol. Rep. 2018, 39, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Tang, F.M.; Pu, D.; Xu, D.; Wang, T.; Li, W. Mechanisms underlying regulation of cell cycle and apoptosis by hnRNP B1 in human lung adenocarcinoma A549 cells. Tumori 2014, 100, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, W.M.; Tang, F.M.; Pu, D.; Chen, W.B. Effects of hnRNP B1 on DNA-PK activity, cell cycle and apoptosis in human lung adenocarcinoma cell line A549. Sichuan Da Xue Xue Bao Yi Xue Ban 2008, 39, 815–818. [Google Scholar] [PubMed]
- Choi, H.S.; Lee, H.M.; Jang, Y.J.; Kim, C.H.; Ryu, C.J. Heterogeneous nuclear ribonucleoprotein A2/B1 regulates the self-renewal and pluripotency of human embryonic stem cells via the control of the G1/S transition. Stem Cells 2013, 31, 2647–2658. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Jiang, Z.; Ma, H.; Ning, L.; Chen, H.; Li, L.; Qi, H. Volatile Oil of Acori Graminei Rhizoma-Induced Apoptosis and Autophagy are dependent on p53 Status in Human Glioma Cells. Sci. Rep. 2016, 6, 21148. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.N.; Wang, J.J.; Shi, L.; Zhang, W.J.; Du, X.Y.; Wang, Z.P.; Zhang, Y. Beta-Asarone induces senescence in colorectal cancer cells by inducing lamin B1 expression. Phytomedicine 2013, 20, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Liu, S.L.; Zhou, J.Y.; Wu, J.; Ling, B.F.; Wang, R.P. Beta-asarone Induces LoVo Colon Cancer Cell Apoptosis by Up-regulation of Caspases through a Mitochondrial Pathway In Vitro and In Vivo. Asian Pac. J. Cancer Prev. 2012, 13, 5291–5298. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, X.X.; Sun, Q.M.; Chen, M.; Liu, S.L.; Zhang, X.; Zhou, J.Y.; Zou, X. Beta-Asarone inhibits gastric cancer cell proliferation. Oncol. Rep. 2015, 34, 3043–3050. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Chen, L.; Ning, L.; Ma, H.; Jiang, Z.; Fu, Y.; Li, L. Proteomic analysis of beta-asarone induced cytotoxicity in human glioblastoma U251 cells. J. Pharm. Biomed. Anal. 2015, 115, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, Q.; Luo, L.; Ning, B.; Fang, Y. Beta-asarone inhibited cell growth and promoted autophagy via P53/Bcl-2/Bclin-1 and P53/AMPK/mTOR pathways in Human Glioma U251 cells. J. Cell. Physiol. 2018, 233, 2434–2443. [Google Scholar] [CrossRef] [PubMed]
- Hongmei Tang, R.L. Determination of chemical components in Rhizoma Acori graminei after permeability of rat blood brain barrier. Tradit. Chin. Med. Res. 2002, 18, 40–41. [Google Scholar]
- Hu, Y.; Yuan, M.; Liu, P.; Mu, L.; Wang, H. [Effect of Acorus tatarinowii schott on ultrastructure and permeability of blood-brain barrier]. Zhongguo Zhong Yao Za Zhi 2009, 34, 349–351. [Google Scholar] [PubMed]
- Meng, X.; Liao, S.; Wang, X.; Wang, S.; Zhao, X.; Jia, P.; Pei, W.; Zheng, X. Reversing P-glycoprotein-mediated multidrug resistance in vitro by alpha-asarone and beta-asarone, bioactive cis-trans isomers from Acorus tatarinowii. Biotechnol. Lett. 2014, 36, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, M.; Wang, C.; Yu, Z.; Wang, H.; Qi, H.; Xu, X. β-Asarone Inhibits Invasion and EMT in Human Glioma U251 Cells by Suppressing Splicing Factor HnRNP A2/B1. Molecules 2018, 23, 671. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, Y.; Lou, Y.; Zhong, H. Knockdown of HNRNPA1 inhibits lung adenocarcinoma cell proliferation through cell cycle arrest at G0/G1 phase. Gene 2016, 576, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Development of cell-cycle checkpoint therapy for solid tumors. Jpn. J. Clin. Oncol. 2015, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed]
- Shilo, A.; Ben Hur, V.; Denichenko, P.; Stein, I.; Pikarsky, E.; Rauch, J.; Kolch, W.; Zender, L.; Karni, R. Splicing factor hnRNP A2 activates the Ras-MAPK-ERK pathway by controlling A-Raf splicing in hepatocellular carcinoma development. RNA 2014, 20, 505–515. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: The compound β-asarone is available from the authors. |


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yang, Y.; Wu, M.; Yu, Z.; Wang, C.; Dou, G.; He, H.; Wang, H.; Yang, N.; Qi, H.; et al. β-Asarone Induces Apoptosis and Cell Cycle Arrest of Human Glioma U251 Cells via Suppression of HnRNP A2/B1-Mediated Pathway In Vitro and In Vivo. Molecules 2018, 23, 1072. https://doi.org/10.3390/molecules23051072
Li L, Yang Y, Wu M, Yu Z, Wang C, Dou G, He H, Wang H, Yang N, Qi H, et al. β-Asarone Induces Apoptosis and Cell Cycle Arrest of Human Glioma U251 Cells via Suppression of HnRNP A2/B1-Mediated Pathway In Vitro and In Vivo. Molecules. 2018; 23(5):1072. https://doi.org/10.3390/molecules23051072
Chicago/Turabian StyleLi, Li, Yi Yang, Mingxia Wu, Zanyang Yu, Chengqiang Wang, Guojun Dou, Hui He, Hongmei Wang, Na Yang, Hongyi Qi, and et al. 2018. "β-Asarone Induces Apoptosis and Cell Cycle Arrest of Human Glioma U251 Cells via Suppression of HnRNP A2/B1-Mediated Pathway In Vitro and In Vivo" Molecules 23, no. 5: 1072. https://doi.org/10.3390/molecules23051072
APA StyleLi, L., Yang, Y., Wu, M., Yu, Z., Wang, C., Dou, G., He, H., Wang, H., Yang, N., Qi, H., & Xu, X. (2018). β-Asarone Induces Apoptosis and Cell Cycle Arrest of Human Glioma U251 Cells via Suppression of HnRNP A2/B1-Mediated Pathway In Vitro and In Vivo. Molecules, 23(5), 1072. https://doi.org/10.3390/molecules23051072