
Response of Cellular Innate Immunity to Cnidarian Pore-Forming Toxins

Abstract
1. Introduction
2. Diverse Toxins Targeting Same Membrane Receptors
3. Pore-Forming Toxins
4. Pore Forming Mechanism in Host Cell Membrane
5. Cnidarian PFTs
5.1. Actinoporins (20–22 kD)
5.2. Hydralysin-Related Toxins (27–31 kD)
5.3. Jellyfish Toxins (42–46 kD)
5.4. MACPF (60 kD)
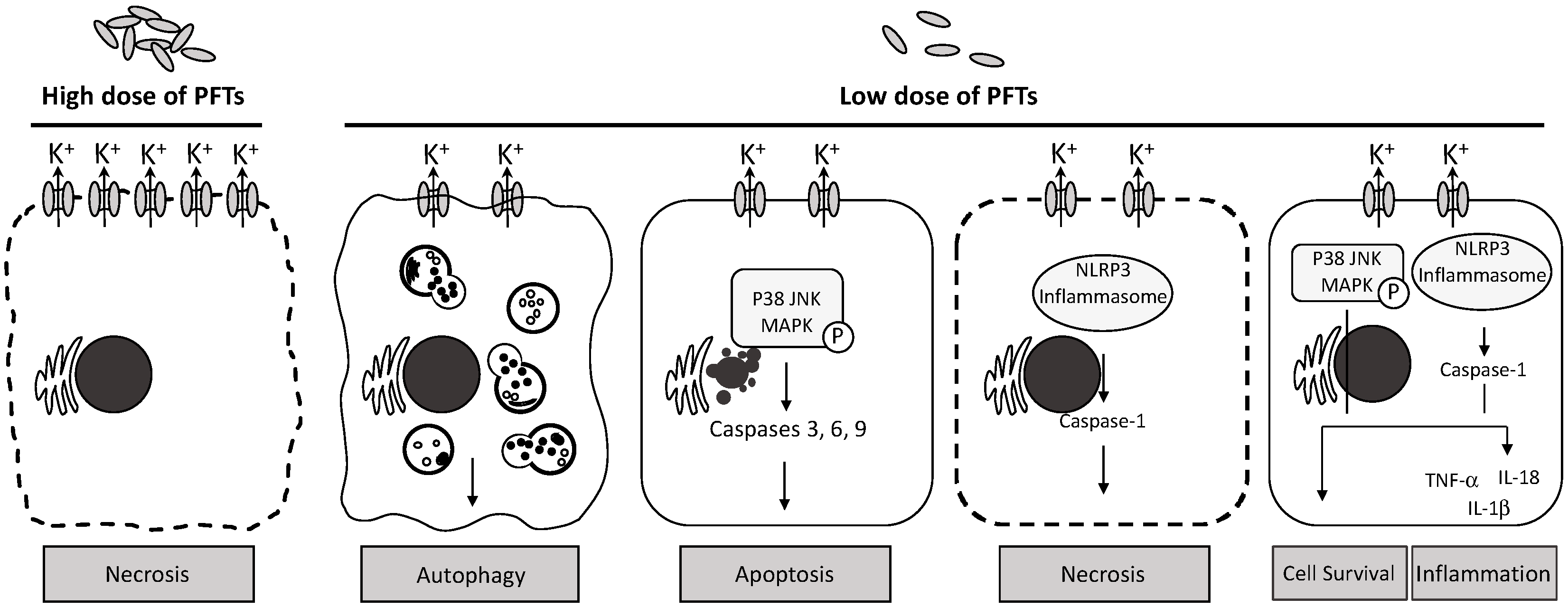
6. Activation of Innate Immunity by Pore-Forming Toxins
7. Molecular Mechanism of Cellular Immune Responses to Actinoporin
7.1. Activation of MAPK Pathways by Actinoporins
7.2. Does Actinoporin Activate NLRP3 Inflammasome?
7.3. Signaling Pathways via Pattern Recognition Receptors
8. What Have We Learnt So Far and How Do We Move Forward?
9. Cnidarian Pore-Forming Toxins for Targeted Toxin Therapy
Funding
Acknowledgments
Conflicts of Interest
References
- Burnett, J.; Calton, G. Jellyfish envenomation syndromes updated. Ann. Emerg. Med. 1987, 16, 1000–1005. [Google Scholar] [CrossRef]
- Russo, A.; Calton, G.; Burnett, J. The relationship of the possible allergic response to jellyfish envenomation and serum antibody titers. Toxicon 1983, 21, 475–480. [Google Scholar] [CrossRef]
- Kemp, S.F.; Lockey, R.F. Anaphylaxis: A review of causes and mechanisms. J. Allergy Clin. Immunol. 2002, 110, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: A review on cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- Tinnballs, J.; Yanagihara, A.A.; Turner, H.C.; Winkel, K. Immunological and toxinological responses to jellyfish stings. Inflamm. Allergy Drug Targets 2011, 10, 438–446. [Google Scholar] [CrossRef]
- Remigante, A.; Costa, R.; Morabito, R.; La Spada, G.; Marino, A.; Dossena, S. Impact of scyphozoan venoms on Human health and current first aid options for stings. Toxins 2018, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Why do we study animal toxins? Zool. Res. 2015, 36, 183–222. [Google Scholar] [PubMed]
- Cornelisse, C.; Harvey, R.; Fisher, B. Lippincott’s Illustrated Reviews: Microbiology, 3rd ed.; Lippincott, Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 69–177. [Google Scholar]
- Wigger, E.; Kuhn-Nentwig, L.; Nentwig, W. The venom optimisation hypothesis: A spider injects large venom quantities only into difficult prey types. Toxicon 2002, 40, 749–752. [Google Scholar] [CrossRef]
- Duterte, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venomes. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Kuo, I.; Ehrlich, B. Signaling in Muscle Contraction. Cold Spring Harb. Perspect. Biol. 2015, 7, a006023. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Groebe, D.R.; Abramson, S.N. Lophotoxin is a slow binding irreversible inhibitor of nicothinic acetylcholine receptors. J. Biol. Chem. 1995, 270, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Gouaux, E. Channel-forming toxins: Tales of transformation. Curr. Opin. Struct. Biol. 1997, 7, 566–573. [Google Scholar] [CrossRef]
- Smith, I.D.M.; Milto, K.M.; Doherty, C.J.; Amyes, S.G.B.; Simpson, A.H.R.W.; Hall, A.C. A potential key role for alpha-haemolysin of Staphylococcus aureus in mediating chondrocyte death in septic arthritis. Bone Jt. Res. 2018, 7, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.; Borges, A.; Simões, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F.; Torres, V.J. The bicomponent pore forming leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubès, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed]
- Abrami, L.; Velluz, M.C.; Hong, Y.; Ohishi, K.; Mehlert, A.; Ferguson, M.; Kinoshita, T.; Gisou van der Goot, F. The glycan core of GPI-anchored proteins modulates aerolysin binding but is not sufficient: The polypeptide moiety is required for the toxin-receptor interaction. FEBS Lett. 2002, 512, 249–254. [Google Scholar] [CrossRef]
- Wilmsen, H.U.; Pattus, F.; Buckley, J.T. Aerolysin, a hemolysin from Aeromonas hydrophila, forms voltage-gated channels in planar lipid bilayers. J. Membr. Biol. 1990, 115, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Gordon, V.M.; Nelson, K.L.; Buckley, J.T.; Stevens, V.L.; Tweten, R.K.; Elwood, P.C.; Leppla, S.H. Clostridium septicum alpha toxin uses glycosylphosphatidyli-nositol-anchored protein receptors. J. Biol. Chem. 1999, 274, 27274–27280. [Google Scholar] [CrossRef] [PubMed]
- Akiba, T.; Abe, Y.; Kitada, S.; Kusaka, Y.; Ito, A.; Ichimatsu, T.; Katayama, H.; Akao, T.; Higuchi, K.; Mizuki, E.; et al. Crystal Structure of the Parasporin-2 Bacillus thuringiensis Toxin That Recognizes Cancer Cells. J. Mol. Biol. 2009, 386, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Griffitts, J.S.; Haslam, S.M.; Yang, T.; Garczynski, S.F.; Mulloy, B.; Morris, H.; Cremer, P.S.; Dell, A.; Adang, M.J.; Aroian, R.V. Glycolipids as receptors for Bacillus thuringiensis crystal toxin. Science 2005, 307, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, S.K.B.; O′Riordan, M.X.D. More than a pore: The cellular response to cholesterol-dependent cytolysins. Toxins 2013, 5, 618–636. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Andrew, P.W.; Boulnois, G.J.; Mitchell, T.J. Molecular analysis of the pathogenicity of Streptococcus pneumoniae: The role of pneumococcal proteins. Annu. Rev. Microbiol. 1993, 47, 89–115. [Google Scholar] [CrossRef] [PubMed]
- Saha, N.; Banerjee, K.K. Carbohydrate-mediated regulation of interaction of Vibrio cholerae hemolysin with erythrocyte and phospholipid vesicle. J. Biol. Chem. 1997, 272, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Leppla, S.H.; Moayeri, M. Bacterial exotoxins and the inflammasome. Front. Immunol. 2015, 6, 570. [Google Scholar] [CrossRef] [PubMed]
- Naglich, J.G.; Metherall, J.E.; Russell, D.W.; Eidels, L. Expression cloning of a diphtheria toxin receptor: Identity with a heparin-binding EGF-like growth factor precursor. Cell 1992, 69, 1051–1061. [Google Scholar] [CrossRef]
- Tateno, H.; Goldstein, I.J. Molecular cloning, expression, and characterization of novel hemolytic lectins from the mushroom Laetiporus sulphureus, which show homology to bacterial toxins. J. Biol. Chem. 2003, 278, 40455–40463. [Google Scholar] [CrossRef] [PubMed]
- Lanio, M.E.; Morera, V.; Alvarez, C.; Tejuca, M.; Gómez, T.; Pazos, F.; Besada, V.; Martínez, D.; Huerta, V.; Padrón, G.; et al. Purification and characterization of two hemolysins from Stichodactyla helianthus. Toxicon 2001, 39, 187–194. [Google Scholar] [CrossRef]
- Macek, P.; Lebez, D. Isolation and characterization of three lethal and hemolytic toxins from the sea anemone Actinia equina L. Toxicon 1988, 26, 441–451. [Google Scholar] [CrossRef]
- Anderluh, G.; Pungercar, J.; Strukelj, B.; Macek, P.; Gubensek, F. Cloning, sequencing, and expression of equinatoxin II. Biochem. Biophys. Res. Commun. 1996, 220, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Bellomio, A.; Morante, K.; Barlič, A.; Gutiérrez-Aguirre, I.; Viguera, A.R.; González-Mañas, J.M. Purification, cloning and characterization of fragaceatoxin C, a novel actinoporin from the sea anemone Actinia fragacea. Toxicon 2009, 54, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Glasser, E.; Rachamim, T.; Aharonovich, D.; Sher, D. Hydra actinoporin-like toxin-1, an unusual hemolysin from the nematocyst venom of Hydra magnipapillata which belongs to an extended gene family. Toxicon 2014, 91, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Sher, D.; Fishman, Y.; Zhang, M.; Lebendiker, M.; Gaathon, A.; Mancheño, J.-M.; Zlotkin, E. Hydralysins: A new category of beta-pore-forming toxins in Cnidaria. Characterization and preliminary structure-function analysis. J. Biol. Chem. 2005, 280, 22847–22855. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Fredman, D.; Szczesny, P.; Grynberg, M.; Technau, U. Recurrent horizontal transfer of bacterial toxin genes to eukaryotes. Mol. Biol. Evol. 2012, 29, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa, K.; Nakao, M.; Ito, E.; Miyake, M.; Noda, M.; Nakajima, T. Novel proteinaceous toxins from the box jellyfish (Sea Wasp) Carybdea rastoni. Biochem. Biophys. Res. Commun. 2000, 275, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa, K.; Nakao, M.; Sakomoto, B.; Crow, G.L.; Nakajima, T. Isolation and characterization of a novel protein from the Hawaiian box jellysfish (Sea Wasp) Carybdea alata. Biochem. Biophys. Res. Commun. 2000, 275, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa, K.; Nakao, M.; Oshiro, N.; Iwanaga, S.; Nakajima, T. A novel protein toxin from the deadly box jellyfish (Sea Wasp, Habu-Kurage) Chiropsalmus quadrigatus. Biosci. Biotechnol. Biochem. 2002, 66, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Avila Soria, G. Molecular Characterization of Carukia barnesi and Malo kingi, Cnidaria; Cubozoa; Carybdeidae. Ph.D. Thesis, James Cook University, Townsville, Australia, 2009. [Google Scholar]
- Nagai, H.; Oshiro, N.; Takuwa-Kuroda, K.; Iwanaga, S.; Nozaki, M.; Nakajima, T. A new polypeptide toxin from the nematocyst venom of an Okinawa sea anemone Phyllodiscus semoni (Japanese name “unbachi-isoginchaku”). Biosci. Biotechnol. Biochem. 2002, 66, 2621–2625. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Oshiro, N.; Iwanaga, S.; Namikoshi, M.; Nagai, H. Characterization of PsTX-60B, a new membrane-attack complex/ perforin (MACPF) family toxin, from the venomous sea anemone Phyllodiscus semoni. Toxicon 2007, 49, 1208–1210. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, N.; Kobayashi, C.; Iwanaga, S.; Nozaki, M.; Namikoshi, M.; Spring, J.; Nagai, H. A new membrane attack complex/perforin (MACPF) domain lethal toxin from the nematocyst venom of the Okinawan sea anemone Actineria villosa. Toxicon 2004, 43, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J.; Hemmrich, G.; Ball, E.E.; Hayward, D.C.; Khalturin, K.; Funayama, N.; Agata, K.; Bosch, T.C. The innate immune repertoire in Cnidaria-ancestral complexity and stochastic gene loss. Genome Biol. 2007, 8, R59. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.; Burnell, J. Identification, cloning and sequencing of two major venom proteins from the box jellyfish, Chironex fleckeri. Toxicon 2007, 50, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L.; Konstantakopoulos, N.; McInerney, B.V.; Mulvenna, J.; Seymour, J.E.; Isbister, G.K.; Hodgson, W.C. Chironex fleckeri (Box Jellyfish) venom proteins: Expansion of a cnidarian toxin family that elicits variable cytolytic and cardiovascular effects. J. Biol. Chem. 2014, 289, 4798–4812. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, S.; Dyshlovoy, S.; Monastyrnaya, M.; Shubina, L.; Leychenko, E.; Kozlovskaya, E.; Jin, J.; Kwak, J.-Y.; Bode, A.M.; Dong, Z.; et al. The anticancer effects of actinoporin RTX-A from the sea anemone Heteractis crispa (=Radianthus macrodactylus). Toxicon 2010, 55, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.W.; Feil, S.C. Pore-forming protein toxins: From structure to function. Prog. Biophys. Mol. Biol. 2005, 88, 91–142. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.; Bischofberger, M.; Pernot, L.; van der Goot, F.; Frêche, B. Bacterial pore-forming toxins: The (w)hole story? Cell. Mol. Life Sci. 2007, 65, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Geny, B.; Popoff, M.R. Bacterial protein toxins and lipids: Pore formation or toxin entry into cells. Biol. Cell 2006, 98, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Skočaj, M.; Bakrač, B.; Križaj, I.; Maček, P.; Anderluh, G.; Sepčić, K. The sensing of membrane microdomains based on pore-forming toxins. Curr. Med. Chem. 2013, 20, 491–501. [Google Scholar] [PubMed]
- Anderluh, G.; Lakey, J. Disparate proteins use similar architectures to damage membranes. Trends Biol. Sci. 2008, 33, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Gutiérrez-Aguirre, I.; Barlič, A.; Malovrh, P.; Kristan, K.; Podlesek, Z.; Maček, P.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; et al. Two-step membrane binding by equinatoxin II, a pore-forming toxin from the sea anemone, involves an exposed aromatic cluster and a flexible helix. J. Biol. Chem. 2002, 277, 41916–41924. [Google Scholar] [CrossRef] [PubMed]
- Bakrač, B.; Gutierrez-Aguirre, I.; Podlesek, Z.; Sonnen, A.; Gilbert, R.; Macek, P.; Lakey, J.; Anderluh, G. Molecular determinants of sphingomyelin specificity of a eukaryotic pore-forming toxin. J. Biol. Chem. 2008, 283, 18665–18677. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, M.P.; Chan, C.M.; Shorr, A.F. Bacteremia in Staphylococcus aureus pneumonia: Outcomes and epidemiology. J. Crit. Care 2011, 26, 395–401. [Google Scholar] [CrossRef] [PubMed]
- DuMont, A.L.; Torres, V.J. Cell targeting by the Staphylococcus aureus pore-forming toxins: It′s not just about lipids. Trends Microbiol. 2014, 22, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wilke, G.A.; Bubeck Wardenburg, J. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus α-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus α-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Inoshima, I.; Inoshima, N.; Wilke, G.A.; Powers, M.E.; Frank, K.M.; Wang, Y.; Wardenburg, J.B. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 2011, 17, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.; Henry, T.; van Rooijen, W.; Perret, M.; Badiou, C.; Aerts, P.; Kemmink, J.; de Haas, C.; van Kessel, K.; Vandenesch, F.; et al. The Staphylococcal Toxin Panton-Valentine Leukocidin Targets Human C5a Receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.; Fernandes da Costa, S.; Bokori-Brown, M.; Naylor, C.; Cole, A.; Moss, D.; Titball, R.; Basak, A. Molecular architecture and functional analysis of NetB, a pore-forming toxin from Clostridium perfringens. J. Biol. Chem. 2013, 288, 3512–3522. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F., III; Kozhaya, L.; Rawlings, S.A.; Reyes-Robles, T.; DuMont, A.L.; Myszka, D.G.; Landau, N.R.; Unutmaz, D.; Torres, V.J. CCR5 is a receptor for Staphylococcus aureus leukotoxin ED. Nature 2013, 493, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Robles, T.; Alonzo, F.; Kozhaya, L.; Lacy, D.; Unutmaz, D.; Torres, V. Staphylococcus aureus leukotoxin ED targets the chemokine receptors CXCR1 and CXCR2 to kill leukocytes and promote infection. Cell Host Microbe 2013, 14, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Alouf, J.E. Pore-Forming Bacterial Protein Toxins: An Overview. In Pore-Forming Toxins; van der Goot, F.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 157, pp. 1–14. [Google Scholar]
- Anderluh, G.; Maček, P. Dissecting the actinoporin pore-forming mechanism. Structure 2003, 11, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2015, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Razpotnik, A.; Podlesek, Z.; Maček, P.; Separovic, F.; Norton, R. Interaction of the Eukaryotic Pore-forming Cytolysin Equinatoxin II with Model Membranes: 19F NMR Studies. J. Mol. Biol. 2005, 347, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Iacovache, I.; van der Goot, F.; Pernot, L. Pore formation: An ancient yet complex form of attack. BBA–Biomembr. 2018, 1778, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Iacovache, I.; Bischofberger, M.; van der Goot, F.G. Structure and assembly of pore-forming proteins. Curr. Opin. Struct. Biol. 2010, 20, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Bischofberger, M.; Gonzalez, M.; van der Goot, F. Membrane injury by pore-forming proteins. Curr. Opin. Cell Biol. 2009, 21, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Lakey, J.H.; Massotte, D.; Heitz, F.; Dasseux, J.; Faucon, J.; Parker, M.W.; Pattus, F. Membrane insertion of the pore-forming domain of colicin A. A spectroscopic study. Eur. J. Biochem. 1991, 196, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Ridley, H.; Johnson, C.L.; Lakey, J.H. Interfacial Interactions of Pore-Forming Colicins. Adv. Exp. Med. Biol. 2010, 677, 81–90. [Google Scholar] [PubMed]
- Bischofberger, M.; Iacovache, I.; van der Goot, F.G. Pathogenic pore-forming proteins: Function and host response. Cell Host Microbe 2012, 12, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Anderluh, G. Pore-forming toxins in Cnidaria. Semin. Cell Dev. Biol. 2017, 72, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Sepčić, K.; Turk, T.; Maček, P. Cytolytic Proteins from Cnidarians-an Overview. Acta Chim. Slov. 2011, 58, 724–729. [Google Scholar] [PubMed]
- Ferlan, I.; Lebez, D. Equinatoxin, a lethal protein from Actinia equina-I. Purification and characterization. Toxicon 1974, 12, 57–61. [Google Scholar] [CrossRef]
- Kem, W.; Dunn, B. Separation and characterization of four different amino acid sequence variants of a sea anemone (Stichodactyla helianthus) protein cytolysin. Toxicon 1988, 26, 997–1008. [Google Scholar] [CrossRef]
- Turk, T.; Kem, W. The phylum Cnidaria and investigations of its toxins and venoms until 1990. Toxicon 2009, 54, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Norton, R. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Kristan, K.C.; Viero, G.; Serra, M.D.; Maček, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Maček, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- García-Ortega, L.; Alegre-Cebollada, J.; García-Linares, S.; Bruix, M.; Martínez-del-Pozo, Á.; Gavilanes, J. The behavior of sea anemone actinoporins at the water–membrane interface. BBA–Biomembr. 2011, 1808, 2275–2288. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, G.; Menestrina, G.; Pederzolli, C.; Krizaj, I.; Gubenšek, F.; Turk, T.; Maček, P. Primary and secondary structure of a pore-forming toxin from the sea anemone, Actinia equina L., and its association with lipid vesicles. BBA–Biomembr. 1994, 1192, 197–204. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Anderluh, G.; Maček, P.; Turk, D. Crystal structure of the soluble form of equinatoxin II, a pore-forming toxin from the sea anemone Actinia equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Liew, Y.; Soh, W.T.; Jiemy, W.F.; Hwang, J.S. Mutagenesis and functional analysis of the pore-forming toxin HALT-1 from Hydra magnipapillata. Toxins 2015, 7, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Mancheño, J.; Tateno, H.; Sher, D.; Goldstein, I. Laetiporus sulphureus Lectin and Aerolysin Protein Family. Adv. Exp. Med. Biol. 2010, 677, 67–80. [Google Scholar] [PubMed]
- Zheng, M.; Fishman, Y.; Sher, D.; Zlotkin, E. Hydralysin, a novel animal group-selective paralytic and cytolytic protein from a noncnidocystic origin in hydra. Biochemistry 2003, 42, 8939–8944. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.; Burnell, J. Biochemical and molecular characterization of cubazoan protein toxins. Toxicon 2009, 54, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Oshiro, N.; Takuwa-Kuroda, K.; Iwanaga, S.; Nozaki, M.; Nakajima, T. Novel proteinaceous toxins from the nematocyst venom of the okinawan sea anemone Phyllodiscus semoni Kwietniewski. Biochem. Biophys. Res. Commun. 2002, 294, 760–763. [Google Scholar] [CrossRef]
- Cabezas, S.; Ho, S.; Ros, U.; Lanio, M.; Alvarez, C.; van der Goot, F. Damage of eukaryotic cells by the pore-forming toxin sticholysin II: Consequences of the potassium efflux. Biochim. Biophys. Acta Biomembr. 2017, 1859, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Porta, H.; Cancino-Rodezno, A.; Soberon, M.; Bravo, A. Role of MAPK p38 in the cellular responses to pore-forming toxins. Peptides 2011, 32, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.R.; Bischofberger, M.; Frêche, B.; Ho, S.; Parton, R.G.; van der Goot, F.G. Pore-forming toxins induce multiple cellular responses promoting survival. Cell. Microbiol. 2011, 13, 1026–1043. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L. The Molecular and Biochemical Characterization of Venom Proteins from the Box Jellyfish, Chironex fleckeri. Ph.D. Thesis, James Cook University, Queensland, Australia, 2008. [Google Scholar]
- Togias, A.; Burnett, J.; Kageysobotka, A.; Lichtenstein, L. Anaphylaxis after contact with a jellyfish. J. Allergy Clin. Immunol. 1985, 75, 672–675. [Google Scholar] [CrossRef]
- Mizuno, M.; Ito, Y.; Morgan, B. Exploiting the Nephrotoxic Effects of Venom from the Sea Anemone, Phyllodiscus semoni, to Create a Hemolytic Uremic Syndrome Model in the Rat. Mar. Drugs 2012, 10, 1582–1604. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Nozaki, M.; Morinea, N.; Suzuki, N.; Nishikaw, K.; Morgan, B.P.; Matsuo, S. A protein toxin from the sea anemone Phyllodiscus semoni targets the kidney and causes a severe renal injury with predominant glomerular endothelial damage. Am. J. Pathol. 2007, 171, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.; Fuentes-Silva, D.; King, G.F. Development of a rational nomenclature for naming peptide and protein toxins from sea anemones. Toxicon 2012, 60, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Saka, H.A.; Gutiérrez, M.G.; Bocco, J.L.; Colombo, M.I. The autophagic pathway: A cell survival strategy against the bacterial pore-forming toxin ci cytolysin. Autophagy 2007, 3, 363–365. [Google Scholar] [CrossRef] [PubMed]
- De Armentia, M.M.L.; Colombo, M.I. Intracellular pathogen invasion of the host cells: Role of α-hemolysin-induced autophagy. In Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection, and Aging; Hayat, M.A., Ed.; Academic Press: Amsterdam, NY, USA, 2015; pp. 135–144. [Google Scholar]
- Fraunholz, M.; Sinha, B. Intracellular Staphylococcus aureus: Live-in and let die. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Bergado, G.; Blanco, R.; Griñán, T.; Rodríguez, H.; Ros, U.; Pazos, F.; Lanio, M.E.; Hernández, A.M. Sticholysin II-mediated cytotoxicity involves the activation of regulated intracellular responses that anticipates cell death. Biochimie 2018, 148, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.; Ley, S. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Huffman, D.; Abrami, L.; Sasik, R.; Corbeil, J.; van der Goot, F.; Aroian, R. Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins. Proc. Natl. Acad. Sci. USA 2004, 101, 10995–11000. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Noguchi, T.; Naguro, I.; Ichijo, H. Apoptosis signal-regulating kinase 1 in stress and immune response. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 199–225. [Google Scholar] [CrossRef] [PubMed]
- Irazoqui, J.; Urbach, J.; Ausubel, F. Evolution of host innate defence: Insights from Caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol. 2010, 10, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Planillo, R.; Kuffa, P.; Martínez-Colón, G.; Smith, B.; Rajendiran, T.; Núñez, G. K+ efflux is the common trigger of NLRP3 inflammasome activation by bacterial toxins and particulate matter. Immunity 2013, 38, 1142–1153. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V. Mechanisms and functions of inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Stutz, A.; Kolbe, C.C.; Stahl, R.; Horvath, G.L.; Franklin, B.S.; van Ray, O.; Brinkschulte, R.; Geyer, M.; Meissner, F.; Latz, E. NLRP3 inflammasome assembly is regulated by phosphorylation of the pyrin domain. J. Exp. Med. 2017, 214, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Tekeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Deguine, J.; Barton, G. MyD88: A central player in innate immune signaling. F1000Prime R. 2014, 6, 97. [Google Scholar] [CrossRef] [PubMed]
- Khilwani, B.; Mukhopadhaya, A.; Chattopadhyay, K. Transmembrane oligomeric form of Vibrio cholera cytolysin triggers TLR2/TLR6-dependent proinflamaatory responses in monocytes and macrophages. Biochem. J. 2015, 466, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Khilwani, B.; Chattopadhyay, K. Signaling beyond punching holes: Modulation of cellular responses by Vibrio cholera sytolysin. Toxins 2015, 7, 3344–3358. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.C.; Mukherjee, G.; Banerjee, P.; Banerjee, K.K.; Biswas, T. Hemolysin induces Toll-like receptor (TLR)-independent apoptosis and multiple TLR-associated parallel activation of macrophages. J. Biol. Chem. 2011, 286, 34542–34551. [Google Scholar] [CrossRef] [PubMed]
- Cegolon, L.; Heymann, W.; Lange, J.; Mastrangelo, G. Jellyfish Stings and Their Management: A Review. Mar. Drugs 2013, 11, 523–550. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Ito, Y.; Morgan, B. Exploiting the nephrotoxic Effects of venom from the sea anemone, Phyllodiscus semoni, to create a hemolytic uremic syndrome model in the rat. Mar. Drugs 2012, 10, 1582–1604. [Google Scholar] [CrossRef] [PubMed]
- Narat, M.; Maček, P.; Kotnik, V.; Sedmak, B. The humoral and cellular immune response to a lipid attenuated pore-forming toxin from the sea anemone Actinia equina L. Toxicon 1994, 32, 65–71. [Google Scholar] [CrossRef]
- Maček, P.; Lebez, D. Kinetics of hemolysis induced by equinatoxin, a cytolytic toxin from the sea anemone Actinia equina. Effect of some ions and pH. Toxicon 1981, 19, 233–240. [Google Scholar] [CrossRef]
- De los Ríos, V.; Oñaderra, M.; Martínez-Ruiz, A.; Lacadena, J.; Mancheño, J.; Martínez del Pozo, Á.; Gavilanes, J. Overproduction in Escherichia coli and Purification of the Hemolytic Protein Sticholysin II from the Sea Anemone Stichodactyla helianthus. Protein Expr. Purif. 2000, 18, 71–76. [Google Scholar] [CrossRef] [PubMed]
- LaRocca, T.; Stivison, E.; Hod, E.; Spitalnik, S.; Cowan, P.; Randis, T.; Ratner, A. Human-specific bacterial pore-forming toxins induce programmed necrosis in erythrocytes. mBio 2014, 5, e01251-14. [Google Scholar] [CrossRef] [PubMed]
- Haslinger, B.; Strangfeld, K.; Peters, G.; Schulze-Osthoff, K.; Sinha, B. Staphylococcus aureus alpha-toxin induces apoptosis in peripheral blood mononuclear cells: Role of endogenous tumour necrosis factor-alpha and the mitochondrial death pathway. Cell. Microbiol. 2003, 5, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Ng, V.; Maeda, S.; Rest, R.; Karin, M. Anthrolysin O and Other Gram-positive Cytolysins Are Toll-like Receptor 4 Agonists. J. Exp. Med. 2004, 200, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Saka, H.; Bidinost, C.; Sola, C.; Carranza, P.; Collino, C.; Ortiz, S.; Echenique, J.; Bocco, J. Vibrio cholerae cytolysin is essential for high enterotoxicity and apoptosis induction produced by a cholera toxin gene-negative V. cholerae non-O1, non-O139 strain. Microb. Pathog. 2008, 44, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Marucci, A.A. Mechanism of action of Staphylococcal alpha-hemolysin. I. Some factors influencing the measurement of alpha-hemolysin. J. Bacteriol. 1963, 86, 1182–1188. [Google Scholar] [PubMed]
- Da Silva, M.; Zahm, J.; Gras, D.; Bajolet, O.; Abely, M.; Hinnrasky, J.; Milliot, M.; de Assis, M.; Hologne, C.; Bonnet, N.; et al. Dynamic interaction between airway epithelial cells and Staphylococcus aureus. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Prince, L.; Graham, K.; Connolly, J.; Anwar, S.; Ridley, R.; Sabroe, I.; Foster, S.; Whyte, M. Staphylococcus aureus Induces Eosinophil Cell. Death Mediated by α-hemolysin. PLoS ONE 2012, 7, e31506. [Google Scholar] [CrossRef] [PubMed]
- Craven, R.R.; Gao, X.; Allen, I.C.; Gris, D.; Wardenburg, J.B.; McElvania-TeKippe, E.; Ting, J.P.; Duncan, J.A. Staphylococcus aureus α-hemolysin activates the NLRP3-inflammasome in human and mouse monocytic cells. PLoS ONE 2009, 4, e7446. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Muhly, M.; Korom, S.; Hugo, F. Release of interleukin-1b associated with potent cytocidal action of Staphylococcal alpha-toxin on human monocytes. Infect. Immun. 1989, 57, 3512–3519. [Google Scholar] [PubMed]
- Yang, W.S.; Park, S.; Yoon, A.; Yoo, J.Y.; Kim, M.K.; Yun, C.; Kim, C. Suicide cancer gene therapy using pore-forming toxin, streptolysin O. Mol. Cancer Ther. 2006, 5, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Hristodorov, D.; Mladenov, R.; Huhn, M.; Barth, S.; Thepen, T. Macrophage-Targeted Therapy: CD64-Based Immunotoxins for Treatment of Chronic Inflammatory Diseases. Toxins 2012, 4, 676–694. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, E.S.; Thorpe, P.E.; Uhr, J.W. Immunotoxins: Magic bullets or misguided missiles? Immunol. Today 1993, 14, 252–259. [Google Scholar] [CrossRef]
- Li, Z.; Yu, T.; Chao, P.; Ma, J. Immunotoxins and cancer therapy. Cell Mol. Immunol. 2005, 2, 106–112. [Google Scholar] [PubMed]
- Govindaraju, A. Immunotoxins: A review of their use in cancer treatment. J. Stem Cells Regen. Med. 2006, 1, 31–36. [Google Scholar]
- Shapira, A.; Benhar, I. Toxin-based therapeutic approaches. Toxin 2010, 2, 2519–2583. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Benhar, I. Antibody-based immunotoxins for the treatment of cancer. Antibodies 2012, 1, 39–69. [Google Scholar] [CrossRef]
- Antignani, A.; FitzGerald, D. Immunotoxins: The role of the toxin. Toxins 2013, 5, 1486–1502. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Liu, Y.; Wang, P. Recombinant immunotoxin therapy of solid tumors: Challenges and strategies. J. Basic Clin. Med. 2013, 2, 1–6. [Google Scholar] [PubMed]
- Tejuca, M.; Barzaga, V.P.; Pazos, F.; Alvarez, C.; Lanio, M.E. Construction of sea anemone cytolysin-based immunotoxins for selective killing of cancer cells. Revista Cubana de Fisica 2009, 26, 15–22. [Google Scholar]
- Tejuca, M.; Anderluh, G.; Dalla Serra, M. Sea anemone cytolysins as toxic components of immunotoxins. Toxicon 2009, 54, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Avila, A.D.; Acosta, C.M.; Lage, A. A new immunotoxin built by linking a hemolytic toxin to a monoclonal antibody specific for immature T lymphocytes. Int. J. Cancer 1988, 42, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Avila, A.D.; de Mateo Acosta, C.; Lage, A. A carcinoembryonic antigen-directed immunotoxin built by linking a monoclonal antibody to a hemolytic toxin. Int. J. Cancer 1989, 43, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Pederzolli, C.; Belmonte, G.; Serra, D.S.; Macek, P.; Menestrina, G. Biochemical and cytotoxic properties of conjugates of transferrin with Equinatoxin II, a cytolysin from sea anemone. Bioconj. Chem. 1995, 6, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Figueredo, D.R.; Roque, L.; Pazos, F.; Martinez, D.; Escobar, N.; Perez, R.; Alvarez, C.; Lanio, M.E. Construction of an immunotoxin with the pore forming protein StI and ior C5, a monoclonal antibody against a colon cancer cell line. Int. Immunopharm. 2004, 4, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Mathew, M.; Verma, R.S. Humanized immunotoxins: A new generation of immunotoxins for targeted cancer therapy. Cancer Sci. 2009, 100, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Mazor, R.; Onda, M.; Park, D.; Addissie, S.; Xiang, L.; Zhang, J.; Hassan, R.; Pastan, I. Dual B- and T-cell de-immunization of recombinant immunotoxin targeting mesothelin with high cytotoxic activity. Oncotarget 2016, 7, 29916–29926. [Google Scholar] [CrossRef] [PubMed]
- Schmohl, J.U.; Todhunter, D.; Taras, E.; Bachanova, V.; Vallera, D.A. Development of a deimmunized bispecific immunotoxin dDT2219 against B-cell malignancies. Toxins 2018, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Mazor, R.; King, E.M.; Oastan, I. Strategies to reduce the immunogenicity of recombinant immunotoxins. Am. J. Pathol. 2018, 188, 1736–1743. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Pore Forming Toxin | Species Name | MW (kDa) a | Type of PFT | Reference |
|---|---|---|---|---|
| α-Hemolysin (Hla) | Staphylococcus aureus | 33.2 | β | [15,16] |
| γ-Hemolysin (LukF and γHLII) | Staphylococcus aureus | 34.3 and 32.5 | β | [16] |
| Leukocidin (LukF and LukS) | Staphylococcus aureus | 34.3 and 32.5 | β | [16,17] |
| Colicin | Escherichia coli | 60 | α | [18] |
| Aerolysin | Aeromonashydrophila | 52 | β | [19,20] |
| α-Toxin | Clostridiumsepticum | 46.5 | β | [21] |
| Parasporin-2 | Bacillus thuringiensis | 37 | β | [22] |
| Cry5B | Bacillus thuringiensis | 140 | α | [23] |
| Tetanolysin O | Clostridium tetani | 55 | β | [24] |
| Pneumolysin | Streptococcus pneumonia | 52 | β | [25] |
| Cytolysin (VCC) | Vibrio cholera | 80 | β | [26] |
| Exotoxin A | Pseudomonas aeruginosa | 38 | α | [27] |
| Diphtheria toxin | Corynebacterium diphtheriae | 58.3 | α | [28] |
| Cytolysin LSL | Laetiporus sulphureus | 35 | β | [29] |
| Sticholysin | Stichodactyla heliantus | 20 | α | [30] |
| Equinatoxin | Actinia equina | 20 | α | [31,32] |
| Fragaceatoxin C | Actinia fragacea | 20 | α | [33] |
| HALTs | Hydra magnipapillata | 20 | α | [34] |
| Hydralysin (Hln) | Chlorohydra viridissima | 27 | β | [35] |
| Nvlysin-1b | Nematostella vectensis | NA | β | [36] |
| CrTX-A | Carybdea rastoni | 43 | α | [37] |
| CrTX-B | Carybdea rastoni | 46 | α | [37] |
| CaTX-A | Carybdea alata | 43 | α | [38] |
| CaTX-B | Carybdea alata | 45 | α | [38] |
| CqTX-A | Chiropsalmus quadrigatus | 44 | α | [39] |
| MkTX-A | Malo kingi | 48.55 | α | [40] |
| MkTX-B | Malo kingi | 43–46 | α | [40] |
| PsTX-60A | Phyllodiscus semoni | 60 | β | [41] |
| PsTX-60B | Phyllodiscus semoni | 60 | β | [42] |
| AvTX-60A | Actineria villosa | 60 | β | [43] |
| HyMac | Hydra magnipapillata | NA | β | [44] |
| Apextrin | Hydra magnipapillata | NA | β | [44] |
| CfTX-1 | Chironex fleckeri | 51.4 | α | [45] |
| CfTX-2 | Chironex fleckeri | 51.7 | α | [45] |
| CfTX-A | Chironex fleckeri | 40 | α | [46] |
| CfTX-B | Chironex fleckeri | 42 | α | [46] |
| RTX-A | Heteractis crispa | 20 | α | [47] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yap, W.Y.; Hwang, J.S. Response of Cellular Innate Immunity to Cnidarian Pore-Forming Toxins. Molecules 2018, 23, 2537. https://doi.org/10.3390/molecules23102537
Yap WY, Hwang JS. Response of Cellular Innate Immunity to Cnidarian Pore-Forming Toxins. Molecules. 2018; 23(10):2537. https://doi.org/10.3390/molecules23102537
Chicago/Turabian StyleYap, Wei Yuen, and Jung Shan Hwang. 2018. "Response of Cellular Innate Immunity to Cnidarian Pore-Forming Toxins" Molecules 23, no. 10: 2537. https://doi.org/10.3390/molecules23102537
APA StyleYap, W. Y., & Hwang, J. S. (2018). Response of Cellular Innate Immunity to Cnidarian Pore-Forming Toxins. Molecules, 23(10), 2537. https://doi.org/10.3390/molecules23102537