Electro-Acupuncture at Neiguan Pretreatment Alters Genome-Wide Gene Expressions and Protects Rat Myocardium against Ischemia-Reperfusion

Abstract
:
1. Introduction
2. Results and Discussion
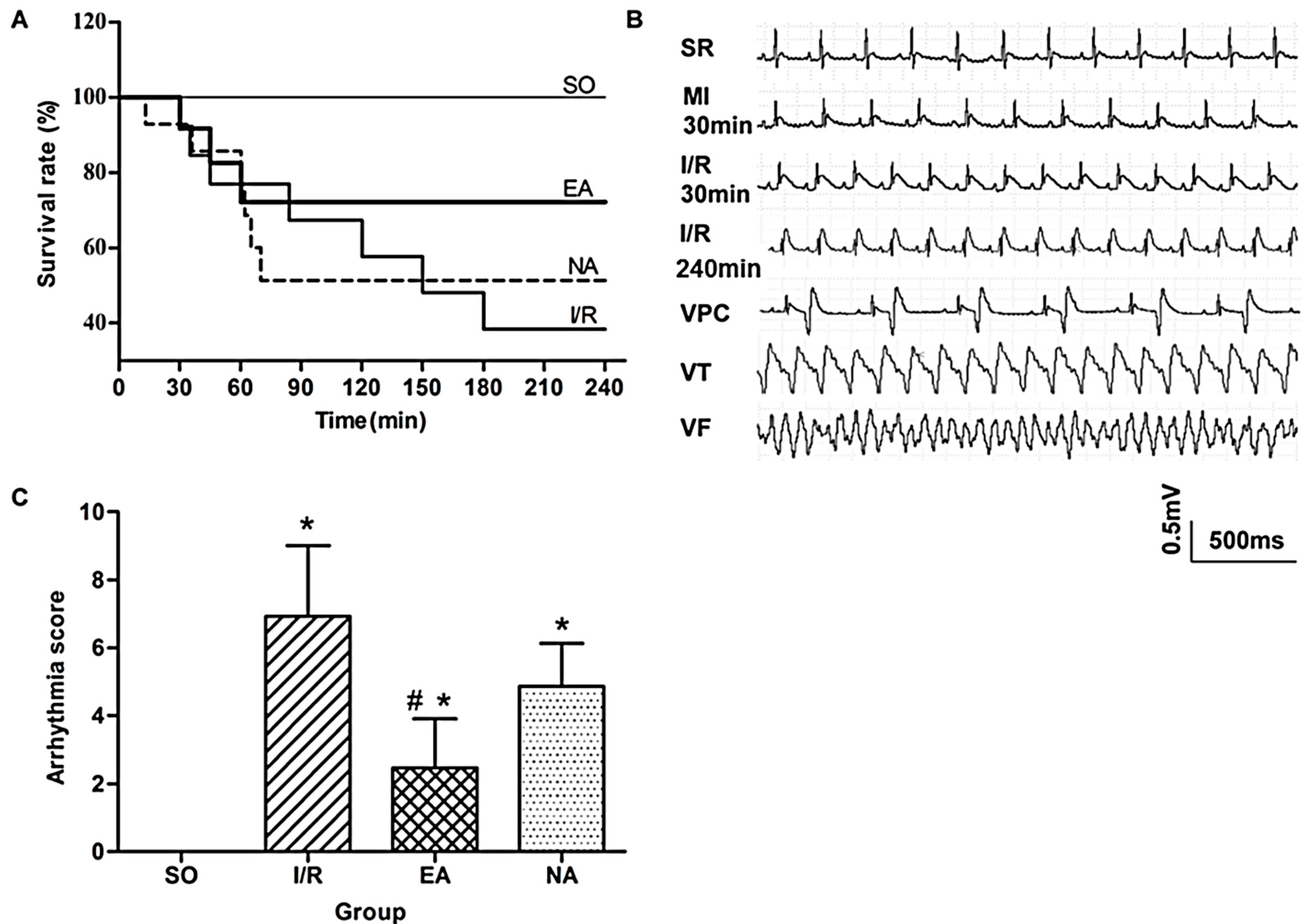
2.1. Electro-Acupuncture at the PC6 Neiguan Acupoint Pretreatment Effectively Protected Myocardium from I/R Injury

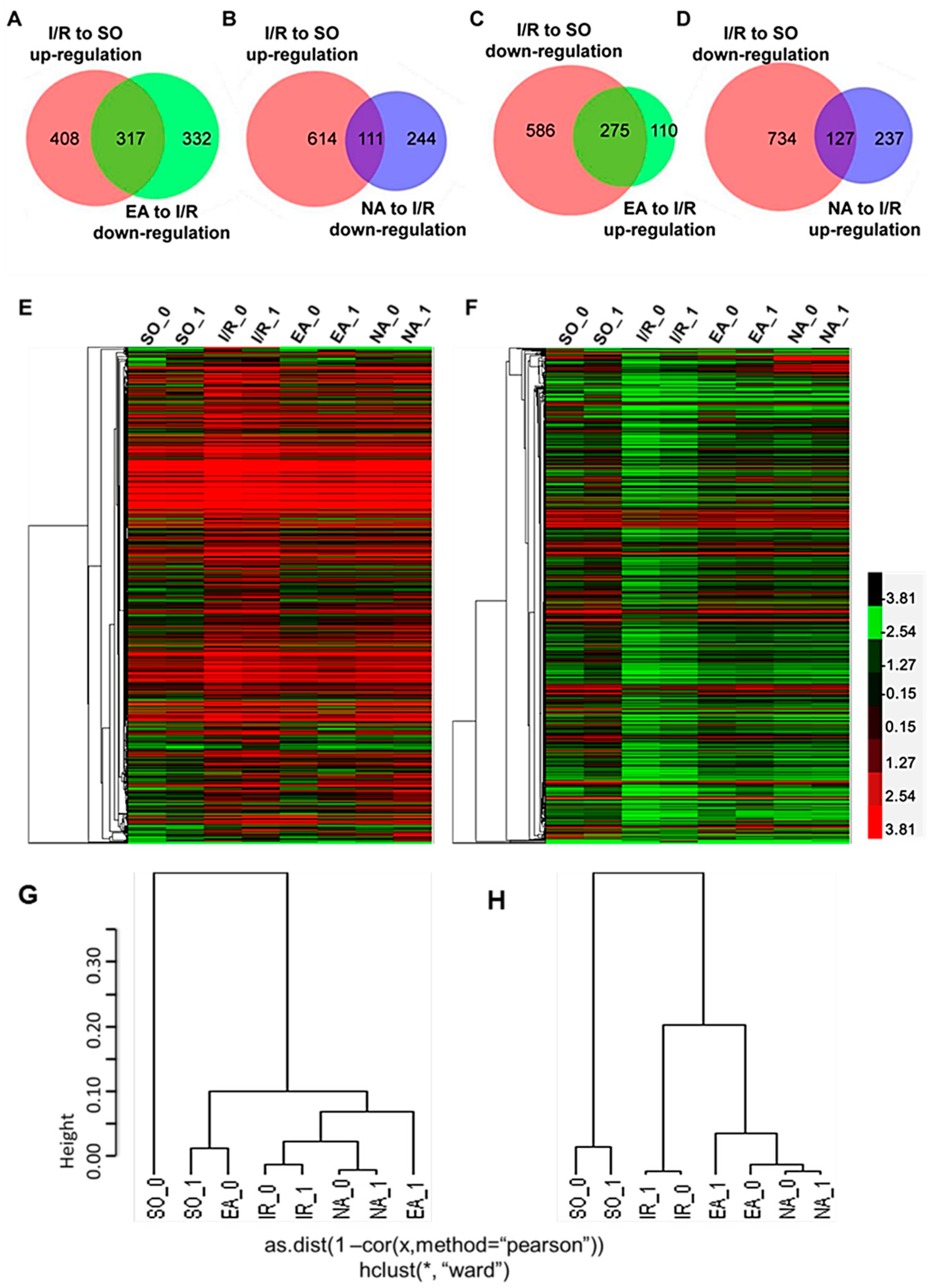
2.2. Electro-Acupuncture at the PC6 Neiguan Acupoint Pretreatment Altered Gene Expressions Genome-Widely

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEGs | I/R:SO | EA:I/R | NA:I/R |
|---|---|---|---|
| Up regulated | 725 | 385 | 364 |
| Down regulated | 861 | 649 | 355 |
| Total | 1586 | 1034 | 719 |
2.2.1. The Most Differentially Expressed Genes
| A: The Top 30 Up-Regulated Genes in I/R | ||||
|---|---|---|---|---|
| Gene Name | Description | I/R vs. SO | EA vs. I/R | NA vs. I/R |
| Hspa1b | heat shock 70 kD protein 1B | 3.58 | −1.12 | 0.97 |
| Mir3074 | microRNA 3074 | 3.1 | −3.13 | 0.6 |
| Kbtbd5 | kelch repeat and BTB (POZ) domain containing 5 | 3.04 | −2.87 | −0.39 |
| Fam46b | family with sequence similarity 46, member B | 2.7 | −1.44 | −1.15 |
| Hist1h1d | histone cluster 1, H1d | 2.7 | - | −1.85 |
| Tnfrsf12a | tumor necrosis factor receptor superfamily, member 12A | 2.67 | −2.39 | −0.43 |
| Fos | FBJ osteosarcoma oncogene | 2.57 | −1.18 | −1.15 |
| Atf3 | activating transcription factor 3 | 2.5 | −1.31 | −0.72 |
| Has1 | hyaluronan synthase 1 | 2.47 | −1.83 | −0.31 |
| Camp | cathelicidin antimicrobial peptide | 2.46 | −0.06 | 1.3 |
| Mir3556b | microRNA 3556b | 2.37 | - | - |
| Vdac3 | voltage-dependent anion channel 3 | 2.28 | 0.3 | −0.14 |
| Trh | thyrotropin releasing hormone | 2.24 | −2.14 | −2.75 |
| Hmox1 | hemeoxygenase (decycling) 1 | 2.23 | −0.78 | 0.7 |
| Cyr61 | cysteine-rich, angiogenic inducer, 61 | 2.22 | −0.98 | −0.09 |
| Pgf | placental growth factor | 2.21 | −1.81 | 0.32 |
| Slc7a5 | solute carrier family 7, member 5 | 2.21 | −1.33 | −0.08 |
| Klk12 | kallikrein related-peptidase 12 | 2.09 | −1.5 | −1.81 |
| Sphk1 | sphingosine kinase 1 | 2.09 | −1.74 | −0.72 |
| Cblc | Cbl proto-oncogene C, E3 ubiquitin protein ligase | 2.03 | −1.84 | −2.24 |
| Nr4a1 | nuclear receptor subfamily 4, group A, member 1 | 2 | −0.72 | −2.35 |
| Numbl | numb homolog (Drosophila)-like | 2 | −1.58 | −1.14 |
| Mt1a | metallothionein 1A | 1.97 | −1.77 | −0.44 |
| Flnc | filamin C, gamma | 1.95 | −1.69 | 0.14 |
| Lcn2 | lipocalin 2 | 1.95 | −0.84 | −3.48 |
| Gal | Galanin | 1.94 | −1.32 | −1.67 |
| Alox15 | arachidonate 15-lipoxygenase | 1.94 | −1.74 | −0.66 |
| Akr1b8 | aldo-keto reductase family 1, member B8 | 1.94 | −1.19 | −0.41 |
| Mt2A | metallothionein 2A | 1.9 | −0.8 | 0.62 |
| Cnot3 | CCR4-NOT transcription complex, subunit 3 | 1.89 | −1.47 | −1.13 |
| LOC367975 | unknown | −5.86 | 2.19 | 2.72 |
| Mir145 | microRNA 145 | −5.32 | - | - |
| Klre1 | killer cell lectin-like receptor family E member 1 | −4.58 | 4.27 | 4.8 |
| Ky | kyphoscoliosis peptidase | −3.62 | 0.78 | 0.42 |
| Clecsf6 | C-type lectin domain family 4, member A | −3.45 | 2.68 | 1.56 |
| RGD1306750 | unknown | −3.21 | 2.09 | 1.78 |
| Cybb | cytochrome b-245, beta polypeptide | −2.83 | 2.38 | 2.45 |
| Ptplad2 | protein tyrosine phosphatase-like A domain containing 2 | −2.78 | 1.91 | 0.86 |
| Cyp2e1 | cytochrome P450, family 2, subfamily E, polypeptide 1 | −2.74 | 2.82 | −0.38 |
| Sh2d1a | SH2 domain containing 1A | −2.71 | 2.4 | 2.73 |
| Tas2r120 | taste receptor, type 2, member 120 | −2.6 | 1.95 | 0.92 |
| Epsti1 | epithelial stromal interaction 1 | −2.57 | 1.84 | 1.74 |
| C6 | complement component 6 | −2.56 | 1.35 | 1.54 |
| F13a1 | coagulation factor XIII, A1 polypeptide | −2.54 | 1.9 | 1.17 |
| Bex4 | brain expressed, X-linked 4 | −2.5 | 1.31 | 1.05 |
| Tfec | transcription factor EC | −2.5 | 2.58 | 2.16 |
| Msr1 | macrophage scavenger receptor 1 | −2.48 | 2.15 | 3.17 |
| Ptprc | protein tyrosine phosphatase, receptor type, C | −2.47 | 0.3 | 1.93 |
| Clec4a2 | C-type lectin domain family 4, member A2 | −2.45 | 2.31 | 1.75 |
| Cd69 | CD69molecule | −2.42 | 1.89 | 1.04 |
| Agr2 | anterior gradient 2 | −2.41 | 0.25 | 1.26 |
| Igsf6 | immunoglobulin superfamily, member 6 | −2.41 | 1.73 | 2.51 |
| Lilra5 | leukocyte immunoglobulin-like receptor, subfamily A, member 5 | −2.38 | 2.06 | −0.23 |
| Klri1 | killer cell lectin-like receptor family I member 1 | −2.34 | 2.18 | 1.73 |
| Ubd | ubiquitin D | −2.32 | 0.46 | 0.98 |
| Ahr | aryl hydrocarbon receptor | −2.32 | 1.78 | 1.33 |
| Ccr2 | chemokine (C-C motif) receptor 2 | −2.31 | 2.52 | 2.7 |
| Pkhd1l1 | polycystic kidney and hepatic disease 1-like 1 | −2.31 | 2.03 | 1.45 |
| Mpeg1 | macrophage expressed 1 | −2.29 | 2.07 | 2.25 |
| Dock11 | dedicator of cytokinesis 11 | −2.28 | 1.62 | 1.19 |

2.2.2. Pathway Analysis
2.2.3. Confirmation of RNA-seq Data by qRT-PCR

3. Experimental Section
3.1. Ethics Statement
3.2. General Reagents
3.3. Experimental Animals and Groups

3.4. In Vivo Myocardial Ischemia-Reperfusion
3.5. Restraint and Electro-Acupuncture Intervention
3.6. Arrhythmia Score
3.7. Infarct Size
3.8. Measurement of Serum Levels of CK, LDH, CK-Mb, Plasma Level of cTnT
3.9. RNA-seq
3.10. Computational Analysis for RNA-seq Data
3.11. qRT-PCR
3.12. Statistics Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jones, D.A.; Andiapen, M.; Van-Eijl, T.J.; Webb, A.J.; Antoniou, S.; Schilling, R.J.; Ahluwalia, A.; Mathur, A. The safety and efficacy of intracoronary nitrite infusion during acute myocardial infarction (NITRITE-AMI): Study protocol of a randomised controlled trial. BMJ Open 2013, 3, e002813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, L.; Huang, G.; Zhou, L.; Wu, W.; Zhang, T.; Huang, H. Protective effect of electroacupuncture at the Neiguan point in a rabbit model of myocardial ischemia-reperfusion injury. Can. J. Cardiol. 2009, 25, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Meng, J. The Effects of Acupuncture in Treatment of Coronary Heart Diseases. J. Tradit. Chin. Med. 2004, 24, 16–19. [Google Scholar] [PubMed]
- Yang, L.; Yang, J.; Wang, Q.; Chen, M.; Lu, Z.; Chen, S.; Xiong, L. Cardioprotective Effects of Electroacupuncture Pretreatment on Patients Undergoing Heart Valve Replacement Surgery: A Randomized Controlled Trial. Ann. Thorac. Surg. 2010, 89, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.T.; Ho, J.Y.; Lin, C.H.; Chiu, J.H. Proteomic analysis finds different myocardial protective mechanisms for median nerve stimulation by electroacupuncture and by local somatothermal stimulation. Int. J. Mol. Med. 2004, 14, 553–563. [Google Scholar] [PubMed]
- Curtis, M.J.; Walker, M.J. Quantification of arrhythmias using scoring systems: An examination of seven scores in an in vivo model of regional myocardial ischaemia. Cardiovasc. Res. 1988, 22, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.P.; He, S.Y.; Xu, B.; Hu, C.J.; Lu, S.F.; Shen, W.X.; Huang, Y.; Hong, H.; Li, Q.; Wang, N.; et al. Acupuncture promotes angiogenesis after myocardial ischemia through H3K9 acetylation regulation at VEGF gene. PLoS One 2014, 9, e94604. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Fu, W.; Jin, Z.; Yu, X. Acupuncture pretreatment protects heart from injury in rats with myocardial ischemia and reperfusion via inhibition of the beta(1)-adrenoceptor signaling pathway. Life Sci. 2007, 80, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Roderic, D.M. Treeview Software, v1.6.6. Available online: http://taxonomy.zoology.gla.ac.uk/rod/treeview.html (accessed on 3 September 2001).
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Suarez, J.; Hu, Y.; McDonough, P.M.; Boer, C.; Dix, D.J.; Dillmann, W.H. Deletion of the inducible 70-kDa heat shock protein genes in mice impairs cardiac ontractile function and calcium handling associated with hypertrophy. Circulation 2006, 113, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Akçetin, Z.; Pregla, R.; Darmer, D.; Heynemann, H.; Haerting, J.; Brömme, H.J. Differential expression of heat shock proteins 70–1 and 70–2 mRNA after ischemia-reperfusion injury of rat kidney. Urol. Res. 1999, 27, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Chorianopoulos, E.; Heger, T.; Lutz, M.; Frank, D.; Bea, F.; Katus, H.A.; Frey, N. FGF-inducible 14-kDa protein (Fn14) is regulated via the RhoA/ROCK kinase pathway in cardiomyocytes and mediates nuclear factor-kappaB activation by TWEAK. Basic Res. Cardiol. 2009, 105, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Causey, M.W.; Salgar, S.; Singh, N.; Martin, M.; Stallings, J.D. Valproic acid reversed pathologic endothelial cell gene expression profile associated with ischemia-reperfusion injury in a swine hemorrhagic shock model. J. Vasc. Surg. 2012, 55, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Fan, P.; Xu, A.; Lam, K.S.; Berger, T.; Mak, T.W.; Tse, H.F.; Yue, J.W.; Song, E.; Vanhoutte, P.M.; et al. Improved functional recovery to I/R injury in hearts from lipocalin-2 deficiency mice: Restoration of mitochondrial function and phospholipids remodeling. Am. J. Transl. Res. 2012, 4, 60–71. [Google Scholar] [PubMed]
- Cheng, Z.; Völkers, M.; Din, S.; Avitabile, D.; Khan, M.; Gude, N.; Mohsin, S.; Bo, T.; Truffa, S.; Alvarez, R.; et al. Mitochondrial translocation of Nur77 mediates cardiomyocyte apoptosis. Eur. Heart J. 2011, 32, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Snoeckx, L.H.; Contard, F.; Samuel, J.L.; Marotte, F.; Rappaport, L. Expression and cellular distribution of heat-shock and nuclear oncogene proteins in rat hearts. Am. J. Physiol. 1991, 261, H1443–H1451. [Google Scholar] [PubMed]
- Zhan, X.; Ander, B.P.; Liao, I.H.; Hansen, J.E.; Kim, C.; Clements, D.; Weisbart, R.H.; Nishimura, R.N.; Sharp, F.R. Recombinant Fv-Hsp70 protein mediates neuroprotection after focal cerebral ischemia in rats. Stroke 2010, 41, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, J.; Nakajima, T.; Osborne, O.C.; Mies, G.; Nowak, T.S. Coexpression of c-fos and hsp70 mRNAs in gerbil brain after ischemia: Induction threshold, distribution and time course evaluated by in situ hybridization. Brain Res. Mol. Brain Res. 1994, 26, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Schiaffonati, L.; Tacchini, L.; Pappalardo, C. Heat shock response in the liver: Expression and regulation of the hsp70 gene family and early response genes after in vivo hyperthermia. Hepatology 1994, 20, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, A.J.; Malt, R.A.; Sukhatme, V.P.; Bonventre, J.V. Expression of two “immediate early” genes, Egr-1 and c-fos, in response to renal ischemia and during compensatory renal hypertrophy in mice. J. Clin. Investig. 1990, 85, 766–771. [Google Scholar] [CrossRef] [PubMed]
- McCommis, K.S.; Baines, C.P. The role of VDAC in cell death: Friend or foe? Biochim. Biophys. Acta 2012, 1818, 1440–1450. [Google Scholar]
- Shoshan-Barmatz, V.; Golan, M. Mitochondrial VDAC1: Function in cell life and death and a target for cancer therapy. Curr. Med. Chem. 2012, 19, 714–735. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Mizrachi, D. VDAC1: From structure to cancer therapy. Front. Oncol. 2012, 29, 164. [Google Scholar] [CrossRef]
- Shimizu, S.; Matsuoka, Y.; Shinohara, Y.; Yoneda, Y.; Tsujimoto, Y. Essential role of voltage-dependent anion channel in various forms of apoptosis in mammalian cells. J. Cell Biol. 2001, 152, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Kaiser, R.A.; Sheiko, T.; Craigen, W.J.; Molkentin, J.D. Voltage-dependent anion channels are dispensable for mitochondrial-dependent cell death. Nat. Cell Biol. 2007, 9, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Mascanfroni, I.; Montesinos Mdel, M.; Susperreguy, S.; Cervi, L.; Ilarregui, J.M.; Ramseyer, V.D.; Masini-Repiso, A.M.; Targovnik, H.M.; Rabinovich, G.A.; Pellizas, C.G. Control of dendritic cell maturation and function by triiodothyronine. FASEB J. 2008, 22, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- García, S.I.; Porto, P.I.; Dieuzeide, G.; Landa, M.S.; Kirszner, T.; Plotquin, Y.; Gonzalez, C.; Pirola, C.J. Thyrotropin-releasing hormone receptor (TRHR) gene is associated with essential hypertension. Hypertension 2001, 8, 683–687. [Google Scholar] [CrossRef]
- Marsh, B.J.; Stevens, S.L.; Hunter, B.; Stenzel-Poore, M.P. Inflammation and the emerging role of the Toll-like receptor system in acute brain ischemia. Stroke 2009, 40, S34–S37. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.L.; Keum, S.; Marchuk, D.A. A novel genetic locus modulates infarct volume independently of the extent of collateral circulation. Phys. Genomics 2013, 45, 751–763. [Google Scholar] [CrossRef]
- Tsujita, K.; Kaikita, K.; Hayasaki, T.; Honda, T.; Kobayashi, H.; Sakashita, N.; Suzuki, H.; Kodama, T.; Ogawa, H.; Takeya, M. Targeted Deletion of Class A Macrophage Scavenger Receptor Increases the Risk of Cardiac Rupture After Experimental Myocardial Infarction. Circulation 2007, 115, 1904–1911. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Hua, F.; Liu, L.; Ha, T.; Kalbfleisch, J.; Schweitzer, J.; Kelley, J.; Kao, R.; Williams, D.; Li, C. Scavenger receptor class-A has a central role in cerebral ischemia-reperfusion injury. J. Cereb. Blood Flow Metab. 2010, 30, 1972–1981. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- DAVID, v6.7. Available online: http://david.abcc.ncifcrf.gov/ (accessed on 5 March 2010).
- Obregón, R.; Ahadian, S.; Ramón-Azcón, J.; Chen, L.; Fujita, T.; Shiku, H.; Chen, M.; Matsue, T. Non-invasive measurement of glucose uptake of skeletal muscle tissue models using a glucose nanobiosensor. Biosens. Bioelectron. 2013, 15, 194–201. [Google Scholar] [CrossRef]
- NCBI GEO Database, Accession GSE61840. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE61840 (accessed on 29 September 2014).
- Ashton, K.J.; Tupicoff, A.; Williams-Pritchard, G.; Kiessling, C.J.; See Hoe, L.E.; Headrick, J.P.; Peart, J.N. Unique transcriptional profile of sustained ligand-activated preconditioning in pre- and post-ischemic myocardium. PLoS One 2013, 8, e72278. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.M.; Colecraft, H.M. L-type calcium channel targeting and local signalling in cardiac myocytes. Cardiovasc. Res. 2013, 98, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Szobi, A.; Rajtik, T.; Carnicka, S.; Ravingerova, T.; Adameova, A. Mitigation of postischemic cardiac contractile dysfunction by CaMKII inhibition: Effects on programmed necrotic and apoptotic cell death. Mol. Cell. Biochem. 2014, 388, 269–276. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) Institute for Laboratory Animal Research. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 1996. [Google Scholar]
- Zingarelli, B.; Salzman, A.L.; Szabó, C. Genetic disruption of poly (ADP-ribose) synthetase inhibits the expression of P-selectin and intercellular adhesion molecule-1 in myocardial ischemia/reperfusion injury. Circ. Res. 1998, 83, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, Y.; Ye, Y.; Rosanio, S.; Tavackoli, S.; Hu, Z.Y.; Schwarz, E.R.; Uretsky, B.F. Prostaglandins mediate the cardioprotective effects of atorvastatin against ischemia-reperfusion injury. Cardiovasc. Res. 2005, 65, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.T.; Huang, C.H.; Chiu, J.H. Electroacupuncture on PC6 (Neiguan) attenuates ischemia/reperfusion injury in rat hearts. Am. J. Chin. Med. 2004, 32, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.M.; Shen, L.L.; Tjen-A-Looi, S.; Pitsillides, K.F.; Li, P.; Longhurst, J.C. Naloxone reverses inhibitory effect of electroacupuncture on sympathetic cardiovascular reflex responses. Am. J. Physiol. 1999, 276, H2127–H2134. [Google Scholar] [PubMed]
- Yu, J.B.; Shi, J.; Gong, L.R.; Dong, S.A.; Xu, Y.; Zhang, Y.; Cao, X.S.; Wu, L.L. Role of Nrf2/ARE Pathway in Protective Effect of Electroacupuncture against Endotoxic Shock-Induced Acute Lung Injury in Rabbits. PLoS One 2014, 9, e104924. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Sun, Z.; Tong, G.; Yi, W.; Ma, L.; Zhao, B.; Cheng, L.; Zhang, J.; Cao, F.; Yi, D. α-Lipoic acid reduces infarct size and preserves cardiac function in rat myocardial ischemia/reperfusion injury through activation of PI3K/Akt/Nrf2 pathway. PLoS One 2013, 8, e58371. [Google Scholar] [CrossRef] [PubMed]
- Image J, Version 1.46d. Available online: http://rsb.info.nih.gov/ij/download.html (accessed on 1 January 2012).
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Cuffdiff, Version 2.1. Available online: http://cufflinks.cbcb.umd.edu/tutorial.html (accessed on 11 April 2013).
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the extract from left ventricular are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Lu, S.-F.; Hu, C.-J.; Fu, S.-P.; Shen, W.-X.; Liu, W.-X.; Li, Q.; Wang, N.; He, S.-Y.; Liang, F.-R.; et al. Electro-Acupuncture at Neiguan Pretreatment Alters Genome-Wide Gene Expressions and Protects Rat Myocardium against Ischemia-Reperfusion. Molecules 2014, 19, 16158-16178. https://doi.org/10.3390/molecules191016158
Huang Y, Lu S-F, Hu C-J, Fu S-P, Shen W-X, Liu W-X, Li Q, Wang N, He S-Y, Liang F-R, et al. Electro-Acupuncture at Neiguan Pretreatment Alters Genome-Wide Gene Expressions and Protects Rat Myocardium against Ischemia-Reperfusion. Molecules. 2014; 19(10):16158-16178. https://doi.org/10.3390/molecules191016158
Chicago/Turabian StyleHuang, Yan, Sheng-Feng Lu, Chen-Jun Hu, Shu-Ping Fu, Wei-Xing Shen, Wan-Xin Liu, Qian Li, Ning Wang, Su-Yun He, Fan-Rong Liang, and et al. 2014. "Electro-Acupuncture at Neiguan Pretreatment Alters Genome-Wide Gene Expressions and Protects Rat Myocardium against Ischemia-Reperfusion" Molecules 19, no. 10: 16158-16178. https://doi.org/10.3390/molecules191016158
APA StyleHuang, Y., Lu, S.-F., Hu, C.-J., Fu, S.-P., Shen, W.-X., Liu, W.-X., Li, Q., Wang, N., He, S.-Y., Liang, F.-R., & Zhu, B.-M. (2014). Electro-Acupuncture at Neiguan Pretreatment Alters Genome-Wide Gene Expressions and Protects Rat Myocardium against Ischemia-Reperfusion. Molecules, 19(10), 16158-16178. https://doi.org/10.3390/molecules191016158