Abstract
Given the growing number of diseases caused by emerging or endemic viruses, original strategies are urgently required: (1) for the identification of new drugs active against new viruses and (2) to deal with viral mutants in which resistance to existing antiviral molecules has been selected. In this context, antiviral peptides constitute a promising area for disease prevention and treatment. The identification and development of these inhibitory peptides require the high-throughput screening of combinatorial libraries. Phage-display is a powerful technique for selecting unique molecules with selective affinity for a specific target from highly diverse combinatorial libraries. In the last 15 years, the use of this technique for antiviral purposes and for the isolation of candidate inhibitory peptides in drug discovery has been explored. We present here a review of the use of phage display in antiviral research and drug discovery, with a discussion of optimized strategies combining the strong screening potential of this technique with complementary rational approaches for identification of the best target. By combining such approaches, it should be possible to maximize the selection of molecules with strong antiviral potential.
1. Introduction
Phage display technology was first used by George Smith in 1985 [1]. It took several years to appreciate the potential of this new tool, but the increasing number of publications relating to its use attests to the versatility of this technique for many applications, including epitope mapping, the isolation of high-affinity proteins, protein engineering and drug discovery. Phage display is based on the expression of the molecules of interest (peptides or proteins) as a fusion with the amino- or carboxy-terminus of a protein present on the surface of the bacteriophage [1,2]. This makes it possible to expose combinatorial protein/peptide libraries on the surface of recombinant phages. The exposed proteins/peptides are then selected by an affinity selection procedure, based on their ability to bind a specific target, such as an antibody, an enzyme, a purified receptor, a nucleic acid or other non protein molecule [2]. The selection process consists of several iterative cycles, each comprising capture, washing and elution steps, for progressive enrichment and amplification of the phage population carrying molecules with a higher affinity for the target. These molecules are then tested individually, to assess their activity or ability to perform the desired function. We focus here on the use of phage display to search for new antiviral compounds in highly diverse peptide libraries. Antibody libraries, another popular application of phage display for the selection of antibodies with high affinity/specificity, is not discussed here.
Antiviral research is an active domain of research, as there are too few molecules available for fighting viral infections and there may even be no available treatment for many neglected viral diseases. There is an urgent need to extend our antiviral arsenal, both to control diseases caused by emerging or endemic viruses and to overcome resistant mutants selected by treatment with the available antiviral molecules. Novel strategies and high-throughput technologies are required to address this need for new drug discovery.
Antiviral molecules must satisfy the same conditions as any therapeutic substance (ADME properties: absorption, distribution, metabolism, excretion): low molecular weight, good solubility, rapid elimination of the target organism, few secondary effects, simple and inexpensive production and easy administration [3] (Figure 1). Moreover, all antiviral molecules targeting the intracellular steps of the viral cycle must be able to enter cells, to express their inhibitory potential [4].
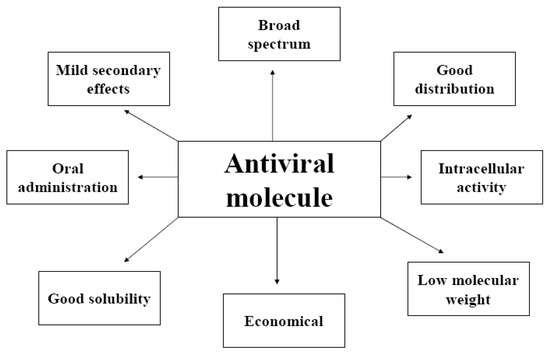
Figure 1.
Principal characteristics of a perfect antiviral molecule.
In parallel to the development of traditional molecules, such as nucleoside analogs, with well demonstrated efficacy, the pharmaceutical industry is now exploring new avenues, such as antiviral peptides. These peptides may prevent viral attachment to host cell receptors or inhibit the replication complex by interfering with protein-protein interactions, dissociating the complex and/or inhibiting its formation [4,5,6,7,8].
Two main approaches can be used to identify such peptides:
- A cognitive approach based on knowledge about the structure/function of the viral components and the interactions they establish within the complex and, possibly, with cellular partners.
- A random approach involving the high-throughput screening of highly diverse combinatorial molecule libraries generated by natural or organic chemistry (nucleic acids, peptides, proteins). This approach is particularly useful for the rapid isolation of candidate antiviral molecules [9] in cases in which little is known about the target protein, ruling out the use of more rational techniques [7].
Peptides have several advantages over traditional chemical compounds, but also a number of disadvantages. On the positive side, they are highly specific and effective, because they mimic the smallest possible part of a functional protein. They also have other very interesting therapeutic characteristics, such as high biodegradability by peptidases present in the body, limiting their accumulation in tissues and resulting in lower toxicity, because their degradation products are amino acids [3]. The potential drawbacks of peptide use are a limited ability to cross membrane barriers, a short half-life (rapid blood clearance) and potential immunogenicity [10], complex modes of action, high production costs and poor oral absorption, often necessitating intravenous administration [3,11]. Nevertheless, it is possible to overcome these limitations [3] through the use of new technologies modifying these molecules and their delivery, stability and application in preclinical settings [12]. For example, D-peptides are resistant to natural proteases (unlike L-peptides), have a longer half-life in serum, can be effectively absorbed orally [8] and are recommended for therapeutic use [13].
2. Isolation of Antiviral Peptides from Combinatorial Libraries by Phage Display
2.1. Construction of combinatorial libraries
The probability of selecting molecules with high affinity is proportional to the size of the library and, thus, to the molecular diversity of the repertoire. Combinatorial peptide libraries generally include 108–1010 clones, but some may include as many as 1013 clones [14]. Generally, maximum diversity is sought [4], but fixed positions can be artificially imposed on the peptide on the basis of data obtained during a previous screening process or to reduce the range of conformations available [15].
Peptides of 6 to 45 amino acids in length may be displayed on the surface of the bacteriophage [16], but the peptide libraries generated for antiviral research usually consist of small peptides, some 6 to 15 amino acids in length (Table 1). The generation of combinatorial peptide libraries with randomized oligonucleotides consisting of NNS / NNK codons (in which N represents A, C, G or T, S represents C or G and K represents G or T) can reduce the frequency of stop codons in peptide sequences, thereby maximizing the diversity of the library [17]. Peptide structure is another key point for the maintenance of peptide functionality. The flanking sequences are particularly important as, when synthesized individually in a soluble form, many peptides totally lose the strong and specific binding affinity they displayed on the phage surface [2,18]. Thus, although linear peptides present the specific advantage of allowing many conformations and adaptability to various structures, this flexibility may decrease their affinity for the target [19]. This problem has been partly overcome by creating libraries in which the peptides are structurally constrained, to limit the number of potential conformers and to make them less dependent on the phage environment. For example, the flanking of random peptide sequences with two cysteine residues, to generate disulfide bridges, limits the number of degrees of freedom and favors certain conformations [20]. The resulting cyclic peptide has a peptide "loop" presenting the variable sequence for interaction [21]. Constrained peptides generally have a higher affinity and specificity [2,20] and a lower conformational entropy, increasing the probability that they will retain their binding capacity when removed from the phage context. In addition, the polypeptide chain must exceed a certain minimum length, to obtain a protein structurally stable in the absence of disulfide bridges [22]. For this reason, libraries of long peptides (>40 amino acids), although not exhaustive, have proved more effective for the isolation of binders than libraries of short peptides [2].
The most widely used bacteriophages are the filamentous phages infecting E. coli, which can grow at extremely high titers (1012–1013 pfu/mL) and have a single-strand DNA genome that is easy to manipulate [17]. It is therefore possible to present highly diverse random peptide libraries on their surface [23]. Bacteriophage M13, which tolerates the insertion of very long fragments of exogenous DNA into its genome, is an ideal tool for phage display [24,25], although other bacteriophages, such as fd, f1, T4 and T7, have also been used. Bacteriohages f1 and fd are filamentous, much like M13, and were used in pioneering studies during the development of phage display [26], whereas the use of bacteriophages T4 and T7, which have an icosahedral capsid, is much less common. Nevertheless, several studies have shown that more diverse libraries (2 to 14 times as diverse) can be presented on bacteriophage T7, with sequences displaying lower charge and hydrophobicity bias than libraries presented on bacteriophage M13 [27]. Only marginal use is made of these phages in phage display, probably because their replication induces bacterial lysis [28]. By contrast, M13 phage particles assemble in the cytoplasm and are secreted without destroying the bacteria [23], simplifying particle purification.
For antiviral purposes, as in other applications, scientists are faced with two choices when designing combinatorial peptide libraries: which bacteriophage surface protein should be used for peptide display and what type of vector should be used for cloning?
In theory, the peptide of interest can be fused to any surface protein, but, in practice, the pIII and pVIII are the most effective and widely used [24]. The displayed protein or peptide is usually fused to the N-terminal region, between the signal peptide and the N-terminus of the mature protein. In antiviral research, pIII is the protein of choice for the exposure of peptide libraries, because only five copies of this protein per phage are present. Moreover, pIII can accommodate large proteins and peptides with little loss of functionality concerning the ability of the phage to infect bacteria [1,29]. By comparison, the pVIII protein, although much more abundant, with 2,700 copies per phage, can display only short peptides (6 to 8 amino acids), due to geometric constraints during assembly of the phage capsid [2]. Moreover, the presence of large numbers of copies of the chimeric protein on the phage surface may favor the selection of clones expressing peptides with a lower affinity for the target (phenomenon of avidity) [25,30].
Three methods can be used to clone libraries of peptide genes fused to the phage coat protein gene [4,15]. The first method is based on fusion of the genes encoding the peptide or protein and the phage capsid protein in an expression vector called a phagemid [31]. A phagemid is a plasmid with the origin of replication and packaging signal of a bacteriophage and sequences for replication in E. coli. It is used to generate constructs encoding the phage protein fused at its N-terminus to each peptide of the library. The phagemid system requires the infection of the bacterium with a helper phage, which provides all the other necessary proteins for genome encaspidation, replication and assembly. The key advantage of a phagemid vector is that it generates one copy of each peptide per phage (monovalent display, the other proteins being encoded by the phage genome itself), preventing avidity effects due to expression of multiple copies of the peptide and therefore increasing the stringency of selection [32,33]. This system can also be used to clone large DNA fragments [19] and limits possible changes to phage infection capacity by reducing the number of chimeric proteins at the surface [31]. Indeed, after synthesis and assembly in bacteria, infectious recombinant phages are released into the culture medium, each phage containing a specific phagemid and displaying at its surface both the wild-type protein and the chimeric protein encoded by the phagemid.
The second method involves inserting the chimeric gene directly into the phage genome [4]. This is a simpler process, because infection with a helper phage is not required. However, it has the drawback that all the copies of the phage surface proteins are fused to the peptide, resulting in multivalent display and a higher risk of selecting less specific binders due to the avidity. The advantages and disadvantages of each system must be taken into account when designing libraries. Monovalent display is generally more appropriate for the characterization of high-affinity peptide inhibitors [19,32,33].
In the third method, the phage genome carries two genes, encoding two different types of protein: one recombinant and the other wild-type. The resulting phage coat is a mosaic displaying both the wild-type and the recombinant protein; it is thus possible display recombinant proteins on the phage surface even if they cannot support phage assembly [15].
2.2. Candidate selection
The affinity selection approach is based on reversible binding between a protein library (antibodies, peptides or enzymes) and a target [2]. Selected targets can be fixed onto a solid phase under conditions similar to those used in ELISA (target used to coat plastic wells) or fused to biotin captured on magnetic beads coated with streptavidin [17,25]. Several rounds of selection/amplification are required to obtain specific binders from large repertoires expressed on the surface of phages. Selection is often carried out with the purified target and involves the following steps: binding of the phages to the target, washing to remove less specific binders, elution to obtain the phages with the best affinity and, finally, the infection of fresh E. coli host bacteria with the selected phage for amplification [17]. The repetition of these binding-elution-amplification steps in vitro greatly increases the proportion of rare selected phages displaying a strong and specific interaction with the protein or peptide target in high-diversity libraries (up to 109 different clones). Typically three to five rounds of selection under conditions of increasing stringency are required to isolate high-affinity binders [4].
As each phage in the library contains the nucleotide sequence encoding the peptide displayed on its surface (in its genome or in the phagemid), characterization of the selected peptides displayed on the phage is straightforward, and is based on the amplification and sequencing of the corresponding genes. When a population of recombinant phages with a highly specific binding signal is obtained, their nucleotide sequences can be analyzed to deduce the global skeleton of important amino acids, which are then fixed through the generation of a sublibrary. The inhibitory activity of the most potent selected phage sequences is then validated, either with a synthetic form of the peptides or by subcloning the corresponding gene and producing the peptide in larger amounts. Validation is based on appropriate functional antiviral assays for the virus studied, such as plaque assays [14], an immunofluorescence assays [34], inoculated cell viability assays [35] or luciferase assays [36,37].
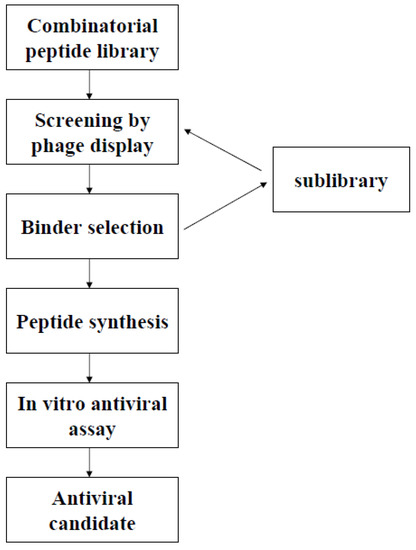
Figure 2.
Process for the isolation of an antiviral peptide from combinatorial libraries by phage display techniques.
Phage display techniques thus have two major advantages for antiviral research. They can be used for the rapid high-throughput screening of large numbers of different molecules (up to 1013) in a small volume (a few microliters). The feasibility of selecting a phage with a frequency of 1/108 in the original library has been demonstrated [23], for example. In addition, phage display defines a physical and molecular link between genotype and phenotype, because the gene encoding the protein (or peptide) exposed at the surface is directly sequenced and isolated from phage DNA [2]. Furthermore, some phagemid vectors can also be directly used as expression vectors, for production of the peptide/protein of interest [38]. Finally, the use of phages in a display strategy has several other specific advantages in itself: (i) the small size of the genome, simplifying the cloning steps [29]; (ii) the stability of phage particles, which can be stored at 4 °C; (iii) the efficiency of phage infection, making it possible to obtain large amounts of material, compatible with the screening of large libraries; (iv) phage display libraries can be periodically regenerated, unlike chemical libraries, which are consumed over time [2].
Traditional methods for generating new drugs involve the in vitro screening of hundreds of thousands of compounds against the selected targets, followed by appropriate in vivo activity assays. Viral diseases are a particular problem in developing countries. It must therefore be possible to produce and sell antiviral molecules as cheaply as possible [39,40]. The characteristics of phage display make this a cheap and powerful and method for discovering peptides able to bind the target with a high affinity and specificity, which could be improved to produce new therapeutic molecules [12].
However, phage display technology has several potential drawbacks that must be taken into account [2]. Clones may be toxic or may reduce phage infectivity and secretion or bacterial growth. Such clones are unlikely to be selected and are therefore likely to disappear rapidly from the library, regardless of the selection process. In addition, the level of expression of certain peptide sequences in E. coli is variable (depending on codon usage), introducing another potential bias into the selection process [41]. Finally, successive amplifications of the library may rapidly decrease the diversity of the library, because the clones with a growth advantage are likely to increase significantly within the population, at the expense of other clones. It has been shown that lower affinity clones may be isolated more efficiently than high-affinity clones during the selection steps in some cases [42]. This phenomenon has mostly been reported with antibody libraries, but may also operate in the case of antiviral molecules from combinatorial peptide libraries. The factors involved are not yet understood but may be related to the protein/peptide sequences displayed, affecting the efficiency of expression and/or being toxic for E. coli. Thus, selection may be dependent on the production of a less toxic phage-peptide and/or favored expression, rather than just the optimization of phage-peptide affinity for the target [43]. In addition, peptide motifs, such as those rich in hydrophobic residues, may interact non specifically with any molecule or surface present during experimental selection (plastic solid phase, for example) [44]. Unfortunately, as mentioned above, the selected peptides, once synthesized as soluble compounds, may not retain their optimal activity outside the microenvironment of the phage and/or in the complicated environment of a living cell or animal [25].
3. The Choice of the Target Protein: A Critical Decision
Antiviral molecules should target key steps of the viral life cycle. These steps can be defined, in particular, by studying the structure-function relationships of viral proteins and their cellular partners. All infection steps (Figure 3) can potentially be targeted for the development of antiviral agents: penetration, release of the viral capsid or genome into the cell, transcription/replication, translation, assembly or the release of progeny virus [45]. Molecules targeting internal steps of the viral cycle are confronted by an additional challenge: entering the cell to exert their inhibitory effects. Cellular proteins essential for the viral replication cycle are also potential targets. The designers of antiviral agents tend to prefer agents with the broadest spectrum of antiviral efficiency, for reasons of both efficiency and economy [26,46]. Treatments should therefore preferentially target proteins involved in generic steps of the viral cycle [47].
The phage display of combinatorial peptide libraries has already been used successfully several times over the last 15 years, in the discovery of inhibitory peptides active against viruses (Table 1). The first viruses to be targeted were hepatitis B virus (HBV) and human immunodeficiency virus (HIV), and both these two viruses remain the most frequently targeted, but other viruses from different families were also considered. Diverse strategies and targets have been used, providing an incidental demonstration of the versatility of the method. Two main approaches are used:
- (1). Targeting of the viral proteins responsible for virus entry into cells (extracellular steps)
- (2). Targeting of the various elements of the viral replication complex and the cellular partners of viral proteins within infected cells (intracellular steps).

Table 1.
Examples of antiviral peptides selected by phage display of combinatorial peptide libraries.
| Viral replication step | Virus | Target | Library | Year | Ref. |
|---|---|---|---|---|---|
| Extracellular steps | IBV | whole virus | 12-mer | 2006 | [48] |
| CMV | whole virus | 9-mer | 1999 | [49] | |
| Human rotavirus | whole virus | 15-mer | 2007 | [50] | |
| WSSV | whole virus | 10-mer | 2003 | [51] | |
| NDV | whole virus | Cyclic 7-mer | 2002 | [52,53] | |
| NDV | whole virus | 7-mer | 2005 | [54] | |
| GCHV | whole virus | 9-mer | 2000 | [55] | |
| ANDV | whole virus | 9-mer | 2009 | [34] | |
| AIV (H9N2) | whole virus | 7-mer | 2009 | [35,56] | |
| West-Nile | protein E | Mouse brain cDNA | 2007 | [57] | |
| HCV | protein E2 | 7-mer | 2010 | [59] | |
| HBV | protein domain PreS | 12-mer | 2007 | [58] | |
| HIV-1 | protein gp41 | 10-mer | 1999 | [37] | |
| HIV-1 | protein gp41 | 8-mer / 7-mer | 2007 | [8] | |
| HIV-1 | protein gp41 | 8-mer / 7-mer | 2010 | [13] | |
| Influenza A | protein HA | 15-mer | 2010 | [14] | |
| SNV, HTNV, PHV | integrin alpha/beta | cyclic 9-mer | 2005 | [73] | |
| Intracellular steps | HBV | protein HBcAg | 6-mer | 1995 | [60] |
| HBV | protein HBcAg | C-7-mer-C | 2003 | [20] | |
| HBV | protein HBsAg | C-7-mer-C | 2005 | [62] | |
| HIV | integrase | 7-mer | 2004 | [65] | |
| HIV | protein GAG | 12-mer | 2005 | [66] | |
| HIV | protein Tat | fd pVIII fragments | 2005 | [69] | |
| HIV | protein LcK | 12-mer | 2005 | [71] | |
| HCV | polymerase NS5B | C-7-mer-C | 2003 | [63] | |
| HCV | polymerase NS5B | 7-mer/12-mer/C-9-mer-C | 2008 | [64] | |
| HPV16 | protein E2 | 7-mer / 12-mer | 2003 | [36] | |
| VSV | IFN receptor | 7-mer | 2008 | [75] | |
| HCV | interleukin 10 | 15-mer | 2011 | [76] |
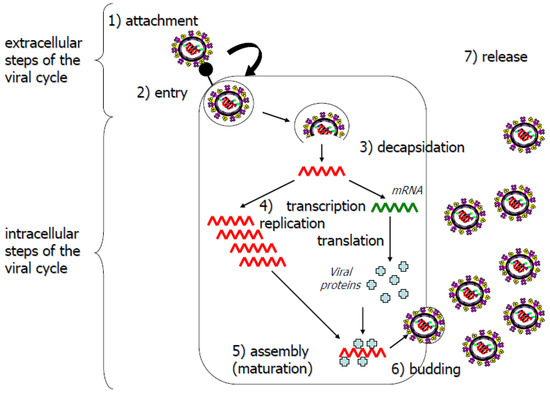
Figure 3.
Simplified diagram of the viral life cycle. Extracellular (1, 2 and 7) and intracellular (3, 4, 5 and 6) steps are indicated.
3.3. Targeting cellular proteins
Targeting a cellular protein playing a role in viral infection or in the antiviral immune response is a particularly appropriate approach for viruses with a high mutation frequency capable of developing resistance against drugs targeting their own proteins.
Again, much of the pioneering research in this domain was carried out on HIV, with the demonstration that a peptide isolated for its affinity for the SH3 domain of the cellular kinase Lck acts as a competitive inhibitor of the HIV Nef protein [71], opening up new perspectives for the development of anti-HIV molecules [72]. However, the use of cellular targets has the particular advantage of providing a potential response to several different viruses. For example, αvβ3 integrin, which is responsible for the entry of hantaviruses into cells, has been used for the screening of a combinatorial cyclic 9-mer peptide library by phage display [73]. Seventy peptides were found to be effective competitors for this receptor and eight of these peptides had a sequence similar to a region of the viral glycoprotein. Some of the selected peptides were able to inhibit infection by Sin Nombre virus (SNV) in a focus reduction assay. In tests assessing the ability of these peptides to inhibit infection by other hantaviruses, the most promising peptides inhibited Hantaan virus (HTNV) infection more effectively than Prospect Hill virus (PHV), which uses a different receptor to enter cells. This strongly suggests that peptides act by competing with the viral glycoprotein for binding to the integrin receptor. The best four peptides, when chemically synthesized independently of the phage, retained their ability to inhibit the entry of SNV into cells, this effect being the most marked when they were used in combination [18].
Another strategy involves enhancing or complementing the immune response, particularly for viruses that naturally counteract innate immunity. Attempts have been made to identify compounds enhancing the immune response and capacity to resist microbial infection [74]. For example, screening of a peptide library against the IFN receptor led to the selection of two candidate peptides mimicking IFN-α antiviral activity against vesicular stomatitis virus (VSV) [75]. Interferon-α can also be stimulated indirectly, by targeting its inhibitors. A 15-mer peptide library has been screened against interleukin 10 (IL-10), a cytokine induced by the HCV core protein and involved in the poor cellular immune response against this virus. Two peptides inhibiting IL-10 were shown to restore the ability of dendritic cells to produce INF-α and to strengthen the anti-HCV T-cell response [76]. This approach is similar to the use of siRNA to target Hsp70, which is overproduced during dengue infection and favors virus multiplication by suppressing type 1 IFN production [77]. Consequently, Hsp70 is a potentially promising cellular target for a phage display strategies aiming to identify peptides active against dengue for use in treatment.
4. Conclusions
Phage display is an efficient technology for selecting peptides with a high affinity for the target, but it is not always possible to obtain candidates with strong antiviral potential by this approach. However, selected peptides can subsequently be improved by rational approaches (drug design, modified residues, mutagenesis, etc.) [24]. In addition, phage display techniques facilitate the emergence of motifs common to selected peptides that can be used to fix positions in a secondary library, thereby retaining the best features of the selected peptides and allowing a gradual improvement in their affinity through successive selections [12].
We have described here success stories resulting in promising antiviral peptide candidates and demonstrating the potential of phage display for use in drug discovery. However, several similar studies have failed to select candidates with sufficient antiviral effects, even when the peptides were selected for their high affinity for the target used. On the one hand, it is not uncommon for molecules displaying significant binding when displayed on the phage surface to lose this capacity and their antiviral effect when synthesized without the carrier and tested in vitro/in vivo. On the other hand, a high capacity to bind the target is not necessarily associated with an antiviral effect, particularly when the binding domain is located outside the active site of the target.
A reverse strategy may therefore constitute a viable alternative approach for increasing the inhibitory potential of molecules. This would involve the initial development of functional screens [78,79], with phage display techniques then used to improve the affinity of the peptides displaying some ability to inhibit the function targeted. This rational approach would help to guide and to limit the degree of “freedom” of peptide libraries, with the phage display combinatorial approach contributing a greater diversity than would be possible with a purely rational approach [25]. The choice of amino acids constituting the candidate peptide can be left random, to provide flexibility and optimize affinity for the target [25]. Directed evolution by phage display has already been used to improve the catalytic properties of enzymes [80,81,82] and to isolate peptides mimicking interferon with stronger biological properties than the wild-type molecule [83,84]. Similarly, the characteristics of the antiviral drug palivizumab, a humanized monoclonal antibody used to treat respiratory syncytial virus (RSV) infection, have been improved by an iterative approach of mutagenesis associated with phage display [85], a method that has also proved effective for an antiviral candidate peptide [61]. Thus, although the considerable potential for the use of phage display in antiviral research is well established, it remains clear how best to use it and how (and when) to integrate it into a broader process including rational, functional and structural studies of the interactions within viral complexes and with cellular partners [24].
With a view to the development of therapeutic applications, it is important to take into account, very early in the selection strategy, all the essential parameters for the design of the molecule, which must ultimately be active in animals or humans: transmembrane penetration, metabolic stability, tissue distribution and elimination, etc. These data should ideally be integrated into the original library. For example, a few small peptide sequences, such as those of penetratin, Tat or VP22, are useful for promoting passage across the membrane and the specific delivery of the peptide to a cell or organ [12]. The generation of peptide libraries fused to a transmembrane signal peptide, such as the HIV Tat peptide, would make it possible to select peptides with a conformation similar to that they are likely to adopt during cell delivery. This would prevent the selection of peptides with characteristics that change during their transformation into therapeutic molecules. In any case, the use of short sequences is commercially advantageous, as it can greatly reduce the cost of production. Identification of the minimal antiviral sequence is therefore of great importance [14].
Good specificity for the target is a general requirement of the candidate peptides. However, when different strains of a virus or different viruses are targeted, extreme specificity may limit the spectrum of activity of the peptide in clinical use [14]. In this case, it is preferable to develop antiviral drugs with a broad spectrum of inhibitory activity. The choice of targets with a structure and/or function common to different viruses is the first step in the antiviral strategy [14]. Screening the library against a protein target conserved among viruses, or successive screenings of the same library against targets of similar function from different viruses could lead to the emergence of molecules with broad-spectrum potential. This approach can be completed by a functional approach involving the testing of molecules with antiviral effects against one virus against related viruses from the same family [57].
For a broad-spectrum approach, proteins involved in virus attachment and penetration are obvious potential targets, but elements of the viral transcription/replication complex and their cellular partners are frequently more strongly conserved and should not be ignored. As mentioned above, one target of choice is the replication ribonucleoprotein complex (RNP) of NSRV, which has a similar structure in unsegmented (Rhabdoviridae, Paramyxoviridae and Filoviridae families) and segmented (Orthomyxoviridae and Bunyaviridae families) viruses, and which typically operates independently of cellular polymerases. In addition, there is no obvious counterpart of this replication complex in humans, suggesting a low likelihood of side effects if used for treatment. Typically, the use of broad-spectrum molecules is the only way to compensate for the low economic attractiveness of neglected diseases to the pharmaceutical industry. Phage display, which has been preferentially applied to diseases with high economic potential, such as HIV or hepatitis, is a unique tool combining original strategies with an exceptional screening potential. It should be applied to neglected diseases against which no antiviral molecule is currently available.
Acknowledgments
Guillaume Castel held a fellowship from Institut Pasteur. Mohamed Chteoui held a fellowship of Fondation Mérieux, 17, rue Bourgelat, Lyon. Bernadette Heyd held a postdoctoral fellowship supported by the ANR project ANRAGE: “Structural Dynamics of the Rabies Virus Replication Complex: Search for New Antiviral Targets” (domain “Microbiology - Immunology - Emerging Diseases”).
References and Notes
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar]
- Souriau, C.; Hua, T.; Lefranc, M.; Weill, M. Présentation à la surface de phages filamenteux: Les multiples applications du phage display. Médecine/Sciences 1998, 14, 300–309. [Google Scholar]
- Decaffmeyer, M.; Thomas, A.; Brasseur, R. Les médicaments peptidiques: Mythe ou réalité ? Biotechnol. Agron. Soc. 2008, 12, 2057. [Google Scholar]
- Brissette, R.; Goldstein, N.I. The use of phage display peptide libraries for basic and translational research. Methods Mol. Biol. 2007, 383, 203–213. [Google Scholar]
- Loregian, A.; Palu, G. Disruption of the interactions between the subunits of herpesvirus DNA polymerases as a novel antiviral strategy. Clin. Microbiol. Infect. 2005, 11, 437–446. [Google Scholar] [CrossRef]
- Loregian, A.; Palu, G. Disruption of protein-protein interactions: Towards new targets for chemotherapy. J. Cell Physiol. 2005, 204, 750–762. [Google Scholar] [CrossRef]
- Vicent, M.J.; Perez-Paya, E.; Orzaez, M. Discovery of inhibitors of protein-protein interactions from combinatorial libraries. Curr. Top. Med. Chem. 2007, 7, 83–95. [Google Scholar]
- Welch, B.D.; VanDemark, A.P.; Heroux, A.; Hill, C.P.; Kay, M.S. Potent D-peptide inhibitors of HIV-1 entry. Proc. Natl. Acad. Sci. USA 2007, 104, 16828–16833. [Google Scholar]
- Vidal, M.; Endoh, H. Prospects for drug screening using the reverse two-hybrid system. Trends Biotechnol. 1999, 17, 374–381. [Google Scholar] [CrossRef]
- Xie, D.; Yao, C.; Wang, L.; Min, W.; Xu, J.; Xiao, J.; Huang, M.; Chen, B.; Liu, B.; Li, X.; Jiang, H. An albumin-conjugated peptide exhibits potent anti-HIV activity and long in vivo half-life. Antimicrob. Agents Chemother. 2010, 54, 191–196. [Google Scholar] [CrossRef]
- Edwards, C.M.; Cohen, M.A.; Bloom, S.R. Peptides as drugs. QJM 1999, 92, 1–4. [Google Scholar] [CrossRef]
- Ladner, R.C.; Sato, A.K.; Gorzelany, J.; de Souza, M. Phage display-derived peptides as therapeutic alternatives to antibodies. Drug Discov. Today 2004, 9, 525–529. [Google Scholar] [CrossRef]
- Welch, B.D.; Francis, J.N.; Redman, J.S.; Paul, S.; Weinstock, M.T.; Reeves, J.D.; Lie, Y.S.; Whitby, F.G.; Eckert, D.M.; Hill, C.P.; Root, M.J.; Kay, M.S. Design of a potent D-peptide HIV-1 entry inhibitor with a strong barrier to resistance. J. Virol. 2010, 84, 11235–11244. [Google Scholar]
- Matsubara, T.; Onishi, A.; Saito, T.; Shimada, A.; Inoue, H.; Taki, T.; Nagata, K.; Okahata, Y.; Sato, T. Sialic acid-mimic peptides as hemagglutinin inhibitors for anti-influenza therapy. J. Med. Chem. 2010, 53, 4441–4449. [Google Scholar]
- Smith, G.P.; Petrenko, V.A. Phage display. Chem. Rev. 1997, 97, 391–410. [Google Scholar]
- Newton, J.R.; Deutscher, S.L. In vivo bacteriophage display for the discovery of novel peptide-based tumor-targeting agents. Methods Mol. Biol. 2009, 504, 275–290. [Google Scholar] [CrossRef]
- Devlin, J.J.; Panganiban, L.C.; Devlin, P.E. Random peptide libraries: A source of specific protein binding molecules. Science 1990, 249, 404–406. [Google Scholar]
- Hall, P.R.; Hjelle, B.; Brown, D.C.; Ye, C.; Bondu-Hawkins, V.; Kilpatrick, K.A.; Larson, R.S. Multivalent presentation of antihantavirus peptides on nanoparticles enhances infection blockade. Antimicrob. Agents Chemother. 2008, 52, 2079–2088. [Google Scholar] [CrossRef]
- Lowman, H.B. Bacteriophage display and discovery of peptide leads for drug development. Annu Rev. Biophys. Biomol. Struct. 1997, 26, 401–424. [Google Scholar] [CrossRef]
- Ho, K.L.; Yusoff, K.; Seow, H.F.; Tan, W.S. Selection of high affinity ligands to hepatitis B core antigen from a phage-displayed cyclic peptide library. J. Med. Virol. 2003, 69, 27–32. [Google Scholar] [CrossRef]
- Ladner, R.C. Constrained peptides as binding entities. Trends Biotechnol. 1995, 13, 426–430. [Google Scholar] [CrossRef]
- Privalov, P.L.; Gill, S.J. Stability of protein structure and hydrophobic interaction. Adv. Protein Chem. 1988, 39, 191–234. [Google Scholar] [CrossRef]
- Sergeeva, A.; Kolonin, M.G.; Molldrem, J.J.; Pasqualini, R.; Arap, W. Display technologies: Application for the discovery of drug and gene delivery agents. Adv. Drug Deliv. Rev. 2006, 58, 1622–1654. [Google Scholar] [CrossRef]
- Pande, J.; Szewczyk, M.M.; Grover, A.K. Phage display: Concept, innovations, applications and future. Biotechnol. Adv. 2010, 28, 849–858. [Google Scholar] [CrossRef]
- Falciani, C.; Lozzi, L.; Pini, A.; Bracci, L. Bioactive peptides from libraries. Chem. Biol. 2005, 12, 417–426. [Google Scholar] [CrossRef]
- Scott, J.K.; Smith, G.P. Searching for peptide ligands with an epitope library. Science 1990, 249, 386–390. [Google Scholar]
- Krumpe, L.R.; Atkinson, A.J.; Smythers, G.W.; Kandel, A.; Schumacher, K.M.; McMahon, J.B.; Makowski, L.; Mori, T. T7 lytic phage-displayed peptide libraries exhibit less sequence bias than M13 filamentous phage-displayed peptide libraries. Proteomics 2006, 6, 4210–4222. [Google Scholar]
- Castagnoli, L.; Zucconi, A.; Quondam, M.; Rossi, M.; Vaccaro, P.; Panni, S.; Paoluzi, S.; Santonico, E.; Dente, L.; Cesareni, G. Alternative bacteriophage display systems. Comb. Chem. High Throughput Screen. 2001, 4, 121–133. [Google Scholar]
- Burritt, J.B.; Bond, C.W.; Doss, K.W.; Jesaitis, A.J. Filamentous phage display of oligopeptide libraries. Anal. Biochem. 1996, 238, 1–13. [Google Scholar] [CrossRef]
- Levin, A.M.; Weiss, G.A. Optimizing the affinity and specificity of proteins with molecular display. Mol. Biosyst. 2006, 2, 49–57. [Google Scholar] [CrossRef]
- Sidhu, S.S. Engineering M13 for phage display. Biomol. Eng. 2001, 18, 57–63. [Google Scholar] [CrossRef]
- O'Connell, D.; Becerril, B.; Roy-Burman, A.; Daws, M.; Marks, J.D. Phage versus phagemid libraries for generation of human monoclonal antibodies. J. Mol. Biol. 2002, 321, 49–56. [Google Scholar] [CrossRef]
- Jestin, J.L. Functional cloning by phage display. Biochimie 2008, 90, 1273–1278. [Google Scholar] [CrossRef]
- Hall, P.R.; Hjelle, B.; Njus, H.; Ye, C.; Bondu-Hawkins, V.; Brown, D.C.; Kilpatrick, K.A.; Larson, R.S. Phage display selection of cyclic peptides that inhibit Andes virus infection. J. Virol. 2009, 83, 8965–8969. [Google Scholar]
- Rajik, M.; Jahanshiri, F.; Omar, A.R.; Ideris, A.; Hassan, S.S.; Yusoff, K. Identification and characterisation of a novel anti-viral peptide against avian influenza virus H9N2. Virol. J. 2009, 6, 74. [Google Scholar] [CrossRef]
- Fujii, T.; Austin, D.; Guo, D.; Srimatkandada, S.; Wang, T.; Kubushiro, K.; Masumoto, N.; Tsukazaki, K.; Nozawa, S.; Deisseroth, A.B. Peptides inhibitory for the transcriptional regulatory function of human papillomavirus E2. Clin. Cancer Res. 2003, 9, 5423–5428. [Google Scholar]
- Eckert, D.M.; Malashkevich, V.N.; Hong, L.H.; Carr, P.A.; Kim, P.S. Inhibiting HIV-1 entry: Discovery of D-peptide inhibitors that target the gp41 coiled-coil pocket. Cell 1999, 99, 103–115. [Google Scholar] [CrossRef]
- Heyd, B.; Pecorari, F.; Collinet, B.; Adjadj, E.; Desmadril, M.; Minard, P. In vitro evolution of the binding specificity of neocarzinostatin, an enediyne-binding chromoprotein. Biochemistry 2003, 42, 5674–5683. [Google Scholar] [CrossRef]
- Knobel, D.L.; Cleaveland, S.; Coleman, P.G.; Fevre, E.M.; Meltzer, M.I.; Miranda, M.E.; Shaw, A.; Zinsstag, J.; Meslin, F.X. Re-evaluating the burden of rabies in Africa and Asia. Bull World Health Organ. 2005, 83, 360–368. [Google Scholar]
- Massé, N.; Selisko, B.; Malet, H.; Peyrane, F.; Debarnot, C.; Decroly, E.; Benarroch, D.; Egloff, M.; Guillemot, J.; Alvarez, K.; Canard, B. Le virus de la dengue: Cibles virales et antiviraux. Virologie 2007, 11, 121–133. [Google Scholar]
- Rodi, D.J.; Soares, A.S.; Makowski, L. Quantitative assessment of peptide sequence diversity in M13 combinatorial peptide phage display libraries. J. Mol. Biol. 2002, 322, 1039–1052. [Google Scholar]
- Malone, J.; Sullivan, M.A. Analysis of antibody selection by phage display utilizing anti-phenobarbital antibodies. J. Mol. Recognit. 1996, 9, 738–745. [Google Scholar] [CrossRef]
- Schier, R.; Marks, J.D. Efficient in vitro affinity maturation of phage antibodies using BIAcore guided selections. Hum. Antibodies Hybridomas 1996, 7, 97–105. [Google Scholar]
- Adey, N.B.; Mataragnon, A.H.; Rider, J.E.; Carter, J.M.; Kay, B.K. Characterization of phage that bind plastic from phage-displayed random peptide libraries. Gene 1995, 156, 27–31. [Google Scholar] [CrossRef]
- Leyssen, P.; De Clercq, E.; Neyts, J. Molecular strategies to inhibit the replication of RNA viruses. Antiviral Res. 2008, 78, 9–25. [Google Scholar] [CrossRef]
- Aman, M.J.; Kinch, M.S.; Warfield, K.; Warren, T.; Yunus, A.; Enterlein, S.; Stavale, E.; Wang, P.; Chang, S.; Tang, Q.; Porter, K.; Goldblatt, M.; Bavari, S. Development of a broad-spectrum antiviral with activity against Ebola virus. Antiviral Res. 2009, 83, 245–251. [Google Scholar]
- Bray, M. Highly pathogenic RNA viral infections: Challenges for antiviral research. Antiviral Res. 2008, 78, 1–8. [Google Scholar] [CrossRef]
- Peng, B.; Chen, H.; Tan, Y.; Jin, M.; Guo, A. Identification of one peptide which inhibited infectivity of avian infectious bronchitis virus in vitro. Sci. Chin. C Life Sci. 2006, 49, 158–163. [Google Scholar] [CrossRef]
- Gough, K.C.; Cockburn, W.; Whitelam, G.C. Selection of phage-display peptides that bind to cucumber mosaic virus coat protein. J. Virol. Methods 1999, 79, 169–180. [Google Scholar] [CrossRef]
- Yao, N.; Yao, L.G.; Zhang, X.M.; Guo, T.L.; Kan, Y.C. Screening for peptides of anti-rotavirus by phage-displayed technique. Sheng Wu Gong Cheng Xue Bao 2007, 23, 403–408. [Google Scholar]
- Yi, G.; Qian, J.; Wang, Z.; Qi, Y. A phage-displayed peptide can inhibit infection by white spot syndrome virus of shrimp. J. Gen. Virol. 2003, 84, 2545–2553. [Google Scholar] [CrossRef]
- Ramanujam, P.; Tan, W.S.; Nathan, S.; Yusoff, K. Novel peptides that inhibit the propagation of Newcastle disease virus. Arch. Virol. 2002, 147, 981–993. [Google Scholar] [CrossRef]
- Chia, S.L.; Tan, W.S.; Shaari, K.; Abdul Rahman, N.; Yusoff, K.; Satyanarayanajois, S.D. Structural analysis of peptides that interact with Newcastle disease virus. Peptides 2006, 27, 1217–1225. [Google Scholar] [CrossRef]
- Ozawa, M.; Ohashi, K.; Onuma, M. Identification and characterization of peptides binding to newcastle disease virus by phage display. J. Vet. Med. Sci. 2005, 67, 1237–1241. [Google Scholar]
- Wang, B.; Ke, L.H.; Jiang, H.; Li, C.Z.; Tien, P. Selection of a specific peptide from a nona-peptide library for in vitro inhibition of grass carp hemorrhage virus replication. Virus Res. 2000, 67, 119–125. [Google Scholar] [CrossRef]
- Rajik, M.; Omar, A.R.; Ideris, A.; Hassan, S.S.; Yusoff, K. A novel peptide inhibits the influenza virus replication by preventing the viral attachment to the host cells. Int. J. Biol. Sci. 2009, 5, 543–548. [Google Scholar]
- Bai, F.; Town, T.; Pradhan, D.; Cox, J.; Ashish; Ledizet, M.; Anderson, J.F.; Flavell, R.A.; Krueger, J.K.; Koski, R.A.; Fikrig, E. Antiviral peptides targeting the west nile virus envelope protein. J. Virol. 2007, 81, 2047–2055. [Google Scholar]
- Deng, Q.; Zhai, J.W.; Michel, M.L.; Zhang, J.; Qin, J.; Kong, Y.Y.; Zhang, X.X.; Budkowska, A.; Tiollais, P.; Wang, Y.; Xie, Y.H. Identification and characterization of peptides that interact with hepatitis B virus via the putative receptor binding site. J. Virol. 2007, 81, 4244–4254. [Google Scholar]
- Hong, H.W.; Lee, S.W.; Myung, H. Selection of peptides binding to HCV e2 and inhibiting viral infectivity. J. Microbiol. Biotechnol. 2010, 20, 1769–1771. [Google Scholar]
- Dyson, M.R.; Murray, K. Selection of peptide inhibitors of interactions involved in complex protein assemblies: Association of the core and surface antigens of hepatitis B virus. Proc. Natl. Acad. Sci. USA 1995, 92, 2194–2198. [Google Scholar] [CrossRef]
- Bottcher, B.; Tsuji, N.; Takahashi, H.; Dyson, M.R.; Zhao, S.; Crowther, R.A.; Murray, K. Peptides that block hepatitis B virus assembly: Analysis by cryomicroscopy, mutagenesis and transfection. EMBO J. 1998, 17, 6839–6845. [Google Scholar] [CrossRef]
- Tan, W.S.; Tan, G.H.; Yusoff, K.; Seow, H.F. A phage-displayed cyclic peptide that interacts tightly with the immunodominant region of hepatitis B surface antigen. J. Clin. Virol. 2005, 34, 35–41. [Google Scholar] [CrossRef]
- Amin, A.; Zaccardi, J.; Mullen, S.; Olland, S.; Orlowski, M.; Feld, B.; Labonte, P.; Mak, P. Identification of constrained peptides that bind to and preferentially inhibit the activity of the hepatitis C viral RNA-dependent RNA polymerase. Virology 2003, 313, 158–169. [Google Scholar] [CrossRef]
- Kim, M.S.; Park, C.; Lee, J.H.; Myung, H. Selection and target-site mapping of peptides inhibiting HCV NS5B polymerase using phage display. J. Microbiol. Biotechnol. 2008, 18, 328–333. [Google Scholar]
- Desjobert, C.; de Soultrait, V.R.; Faure, A.; Parissi, V.; Litvak, S.; Tarrago-Litvak, L.; Fournier, M. Identification by phage display selection of a short peptide able to inhibit only the strand transfer reaction catalyzed by human immunodeficiency virus type 1 integrase. Biochemistry 2004, 43, 13097–13105. [Google Scholar]
- Sticht, J.; Humbert, M.; Findlow, S.; Bodem, J.; Muller, B.; Dietrich, U.; Werner, J.; Krausslich, H.G. A peptide inhibitor of HIV-1 assembly in vitro. Nat. Struct. Mol. Biol. 2005, 12, 671–677. [Google Scholar]
- Dietz, J.; Koch, J.; Kaur, A.; Raja, C.; Stein, S.; Grez, M.; Pustowka, A.; Mensch, S.; Ferner, J.; Moller, L.; Bannert, N.; Tampe, R.; Divita, G.; Mely, Y.; Schwalbe, H.; Dietrich, U. Inhibition of HIV-1 by a peptide ligand of the genomic RNA packaging signal Psi. Chem. Med. Chem. 2008, 3, 749–755. [Google Scholar]
- Enshell-Seijffers, D.; Smelyanski, L.; Gershoni, J.M. The rational design of a 'type 88' genetically stable peptide display vector in the filamentous bacteriophage fd. Nucl. Acids Res. 2001, 29, E50. [Google Scholar] [CrossRef]
- Krichevsky, A.; Rusnati, M.; Bugatti, A.; Waigmann, E.; Shohat, S.; Loyter, A. The fd phage and a peptide derived from its p8 coat protein interact with the HIV-1 Tat-NLS and inhibit its biological functions. Antiviral Res. 2005, 66, 67–78. [Google Scholar] [CrossRef]
- Moran, G.J.; Talan, D.A.; Abrahamian, F.M. Biological terrorism. Infect Dis. Clin. North Am. 2008, 22, 145–187. [Google Scholar] [CrossRef]
- Tran, T.; Hoffmann, S.; Wiesehan, K.; Jonas, E.; Luge, C.; Aladag, A.; Willbold, D. Insights into human Lck SH3 domain binding specificity: Different binding modes of artificial and native ligands. Biochemistry 2005, 44, 15042–15052. [Google Scholar] [CrossRef]
- Stangler, T.; Tran, T.; Hoffmann, S.; Schmidt, H.; Jonas, E.; Willbold, D. Competitive displacement of full-length HIV-1 Nef from the Hck SH3 domain by a high-affinity artificial peptide. Biol. Chem. 2007, 388, 611–615. [Google Scholar]
- Larson, R.S.; Brown, D.C.; Ye, C.; Hjelle, B. Peptide antagonists that inhibit Sin Nombre virus and hantaan virus entry through the beta3-integrin receptor. J. Virol. 2005, 79, 7319–7326. [Google Scholar] [CrossRef]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef]
- Zhang, Q.; Bai, G.; Chen, J.Q.; Tian, W.; Cao, Y.; Pan, P.W.; Wang, C. Identification of antiviral mimetic peptides with interferon alpha-2b-like activity from a random peptide library using a novel functional biopanning method. Acta Pharmacol. Sin. 2008, 29, 634–640. [Google Scholar] [CrossRef]
- Diaz-Valdes, N.; Manterola, L.; Belsue, V.; Riezu-Boj, J.I.; Larrea, E.; Echeverria, I.; Llopiz, D.; Lopez-Sagaseta, J.; Lerat, H.; Pawlotsky, J.M.; Prieto, J.; Lasarte, J.J.; Borras-Cuesta, F.; Sarobe, P. Improved dendritic cell-based immunization against hepatitis C virus using peptide inhibitors of interleukin 10. Hepatology 2011, 53, 23–31. [Google Scholar]
- Padwad, Y.S.; Mishra, K.P.; Jain, M.; Chanda, S.; Ganju, L. Dengue virus infection activates cellular chaperone Hsp70 in THP-1 cells: Downregulation of Hsp70 by siRNA revealed decreased viral replication. Viral. Immunol. 2010, 23, 557–565. [Google Scholar] [CrossRef]
- Wunner, W.H.; Pallatroni, C.; Curtis, P.J. Selection of genetic inhibitors of rabies virus. Arch. Virol. 2004, 149, 1653–1662. [Google Scholar]
- Pelet, T.; Miazza, V.; Mottet, G.; Roux, L. High throughput screening assay for negative single stranded RNA virus polymerase inhibitors. J. Virol. Methods 2005, 128, 29–36. [Google Scholar]
- Strobel, H.; Ladant, D.; Jestin, J.L. In vitro selection for enzymatic activity: A model study using adenylate cyclase. J. Mol. Biol. 2003, 332, 1–7. [Google Scholar]
- Soumillion, P.; Jespers, L.; Bouchet, M.; Marchand-Brynaert, J.; Winter, G.; Fastrez, J. Selection of beta-lactamase on filamentous bacteriophage by catalytic activity. J. Mol. Biol. 1994, 237, 415–422. [Google Scholar] [CrossRef]
- Pedersen, H.; Holder, S.; Sutherlin, D.P.; Schwitter, U.; King, D.S.; Schultz, P.G. A method for directed evolution and functional cloning of enzymes. Proc. Natl. Acad. Sci. USA 1998, 95, 10523–10528. [Google Scholar]
- Yamamoto, K.; Taniai, M.; Torigoe, K.; Yamamoto, S.; Arai, N.; Suemoto, Y.; Yoshida, K.; Okura, T.; Mori, T.; Fujioka, N.; Tanimoto, T.; Miyata, M.; Ariyasu, H.; Ushio, C.; Fujii, M.; Ariyasu, T.; Ikeda, M.; Ohta, T.; Kurimoto, M.; Fukuda, S. Creation of interferon-alpha8 mutants with amino acid substitutions against interferon-alpha receptor-2 binding sites using phage display system and evaluation of their biologic properties. J. Interferon Cytokine Res. 2009, 29, 161–170. [Google Scholar]
- Kalie, E.; Jaitin, D.A.; Abramovich, R.; Schreiber, G. An interferon alpha2 mutant optimized by phage display for IFNAR1 binding confers specifically enhanced antitumor activities. J. Biol. Chem. 2007, 282, 11602–11611. [Google Scholar] [CrossRef]
- Wu, H.; Pfarr, D.S.; Tang, Y.; An, L.L.; Patel, N.K.; Watkins, J.D.; Huse, W.D.; Kiener, P.A.; Young, J.F. Ultra-potent antibodies against respiratory syncytial virus: Effects of binding kinetics and binding valence on viral neutralization. J. Mol. Biol. 2005, 350, 126–144. [Google Scholar]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).